
Genome-Wide Selection Signal Analysis to Investigate Wide Genomic Heredity Divergence between Eurasian Wild Boar and Domestic Pig

 , ,
, , {kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
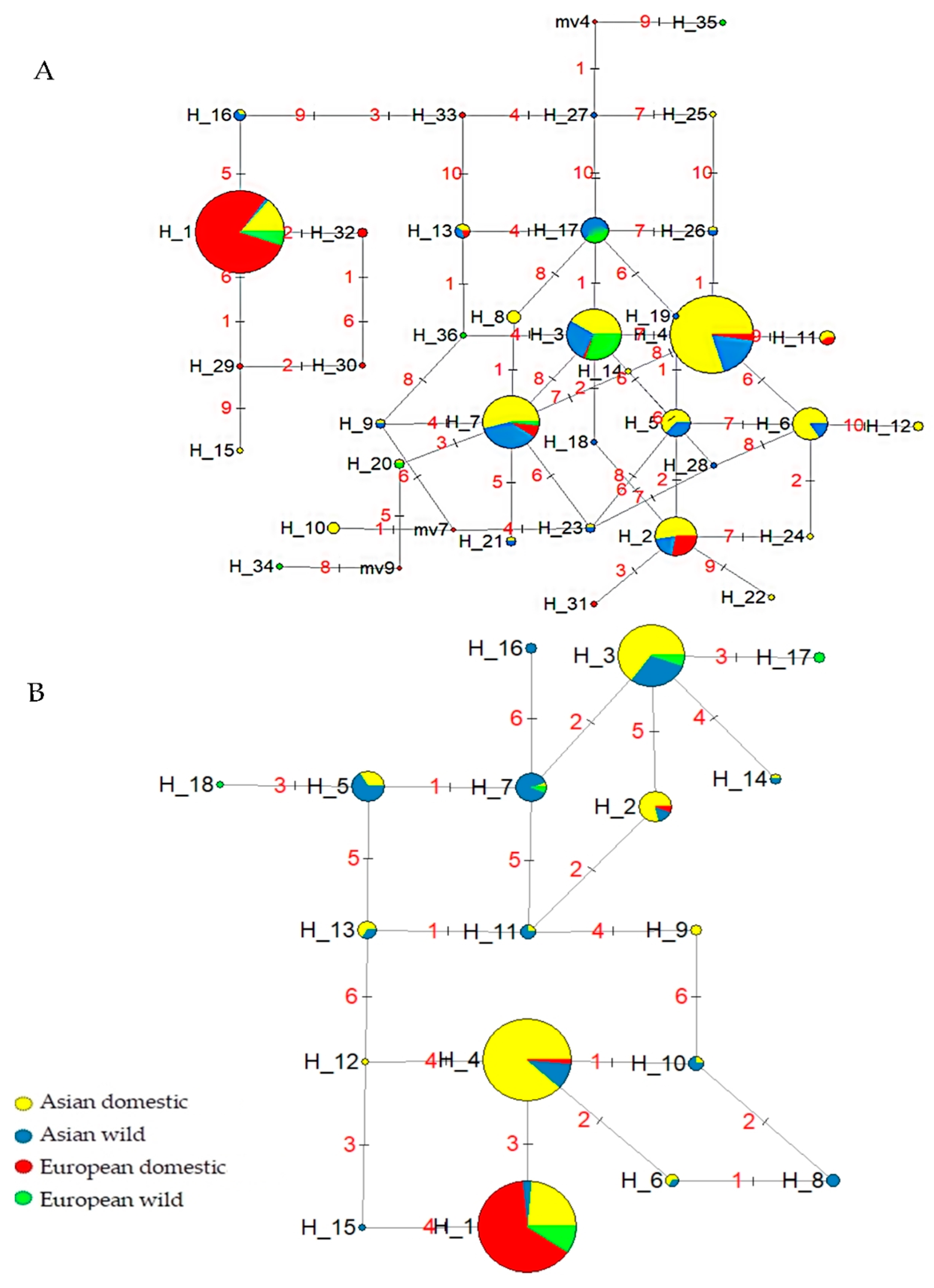
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, J.; Rowan, F. Pig domestication in ancient China. Antiquity 2002, 76, 724–732. [Google Scholar]
- Frantz, L.; Haile, J.; Lin, A.T.; Scheu, A.; Georg, C.; Benecke, N.; Alexander, M.; Linderholm, A.; Mullin, V.E.; Daly, K.G.; et al. Ancient pigs reveal a near-complete genomic turnover following their introduction to Europe. Proc. Natl. Acad. Sci. USA 2019, 116, 17231–17238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiener, P.; Wilkinson, S. Deciphering the genetic basis of animal domestication. Proc. R. Soc. B-Biol. Sci. 2011, 278, 3161–3170. [Google Scholar] [CrossRef] [Green Version]
- Brooker, R.M.; Feeney, W.E. Animal domesticators. Curr. Biol. 2019, 29, R1168–R1169. [Google Scholar] [CrossRef]
- Lord, K.A.; Larson, G.; Coppinger, R.P.; Karlsson, E.K. The history of farm foxes undermines the animal domestication syndrome. Trends Ecol. Evol. 2020, 35, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Maibam, U.; Hooda, O.K.; Sharma, P.S.; Mohanty, A.K.; Singh, S.V.; Upadhyay, R.C. Expression of hsp70 genes in skin of zebu (tharparkar) and crossbred (Karan fries) cattle during different seasons under tropical climatic conditions. J. Therm. Biol. 2017, 63, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Ahbara, A.M.; Musa, H.H.; Robert, C.; Abebe, A.; Al-Jumaili, A.S.; Kebede, A.; Latairish, S.; Agoub, M.O.; Clark, E.; Hanotte, O.; et al. Natural adaptation and human selection of northeast African sheep genomes. Genomics 2022, 114, 110448. [Google Scholar] [CrossRef] [PubMed]
- Groenen, M.A.; Archibald, A.L.; Uenishi, H.; Tuggle, C.K.; Takeuchi, Y.; Rothschild, M.F.; Rogel-Gaillard, C.; Park, C.; Milan, D.; Megens, H.J.; et al. Analyses of pig genomes provide insight into porcine demography and evolution. Nature 2012, 491, 393–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.K.; Kim, K.S.; Ranyuk, M.; Babaev, E.; Voloshina, I.; Bayarlkhagva, D.; Chong, J.R.; Ishiguro, N.; Yu, L.; Min, M.S.; et al. Asia-wide phylogeography of wild boar (sus scrofa) based on mitochondrial DNA and y-chromosome: Revising the migration routes of wild boar in Asia. PLoS ONE 2020, 15, e238049. [Google Scholar] [CrossRef]
- Wang, X.; Tsai, T.; Deng, F.; Wei, X.; Chai, J.; Knapp, J.; Apple, J.; Maxwell, C.V.; Lee, J.A.; Li, Y.; et al. Longitudinal investigation of the swine gut microbiome from birth to market reveals stage and growth performance associated bacteria. Microbiome 2019, 7, 109. [Google Scholar] [CrossRef] [Green Version]
- Kenney, S.P.; Meng, X.J. An SH3 binding motif within the nucleocapsid protein of porcine reproductive and respiratory syndrome virus interacts with the host cellular signaling proteins stami, txk, fyn, hck, and cortactin. Virus Res. 2015, 204, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.D.; Zhang, S.J.; Ding, Y.Y.; Feng, Y.F.; Zhu, H.Y.; Huang, L.; Wu, T.; Zhou, J.; Yin, Z.J. Association between adsl, gars-airs-gart, dgat1, and decr1 expression levels and pork meat quality traits. Genet. Mol. Res. 2015, 14, 14823–14830. [Google Scholar] [CrossRef]
- Liu, Z.; Xu, R.; Zhang, H.; Wang, D.; Wang, J.; Wu, K. A unique 15-bp indel in the first intron of bmpr1b regulates its expression in taihu pigs. BMC Genom. 2022, 23, 799. [Google Scholar] [CrossRef] [PubMed]
- Ropka-Molik, K.; Bereta, A.; Zukowski, K.; Tyra, M.; Piorkowska, K.; Zak, G.; Oczkowicz, M. Screening for candidate genes related with histological microstructure, meat quality and carcass characteristic in pig based on rna-seq data. Asian Australas. J. Anim. Sci. 2018, 31, 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.; Li, S.; Ding, R.; Yang, M.; Zheng, E.; Yang, H.; Gu, T.; Xu, Z.; Cai, G.; Wu, Z.; et al. Meta-analysis of genome-wide association studies for loin muscle area and loin muscle depth in two duroc pig populations. PLoS ONE 2019, 14, e218263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbeck, Y.E.; Gulevich, R.G. Neuropeptides as facilitators of domestication. Cell Tissue Res. 2019, 375, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The variant call format and vcftools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [Green Version]
- Szpiech, Z.A.; Hernandez, R.D. Selscan: An efficient multithreaded program to perform ehh-based scans for positive selection. Mol. Biol. Evol. 2014, 31, 2824–2827. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. Dnasp 6: Dna sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Yang, B.; Cui, L.; Perez-Enciso, M.; Traspov, A.; Crooijmans, R.P.M.A.; Zinovieva, N.; Schook, L.B.; Archibald, A.; Gatphayak, K.; Knorr, C.; et al. Genome-wide SNP data unveils the globalization of domesticated pigs. Genet. Sel. Evol. 2017, 49, 71. [Google Scholar] [CrossRef] [Green Version]
- Wang, J. A Comparative Study on the Domestication Patterns of Chinese and Western Domestic Pigs; China Agricultural University: Beijing, China, 2017. (In Chinese) [Google Scholar]
- Yang, K.; Wang, J.; Wang, K.; Luo, Y.; Tang, Q.; Liu, X.; Fang, M. Integrated analysis of mirna-mrna network reveals different regulatory patterns in the endometrium of meishan and duroc sows during mid-late gestation. Animals 2020, 10, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prochazka, R.; Blaha, M.; Nemcova, L. Signaling pathways regulating fsh- and amphiregulin-induced meiotic resumption and cumulus cell expansion in the pig. Reproduction 2012, 144, 535–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Tello, J.; Schofield, Z.; Kiu, R.; Dalby, M.J.; van Sinderen, D.; Le Gall, G.; Sferruzzi-Perri, A.N.; Hall, L.J. Maternal gut microbiota bifidobacterium promotes placental morphogenesis, nutrient transport and fetal growth in mice. Cell. Mol. Life Sci. 2022, 79, 386. [Google Scholar] [CrossRef]
- Sun, J.; Guo, Y.; Fan, Y.; Wang, Q.; Zhang, Q.; Lai, D. Decreased expression of idh1 by chronic unpredictable stress suppresses proliferation and accelerates senescence of granulosa cells through ros activated mapk signaling pathways. Free Radic. Biol. Med. 2021, 169, 122–136. [Google Scholar] [CrossRef] [PubMed]
- Franca, M.M.; Mendonca, B.B. Genetics of ovarian insufficiency and defects of folliculogenesis. Best Pract. Res. Clin. Endoc. Metab. 2022, 36, 101594. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Kerouanton, B.; Chothani, S.; Zhang, S.; Chen, Y.; Mantri, C.K.; Hock, D.H.; Lim, R.; Nadkarni, R.; Huynh, V.T.; et al. Coding and non-coding roles of mocci (c15orf48) coordinate to regulate host inflammation and immunity. Nat. Commun. 2021, 12, 2130. [Google Scholar] [CrossRef]
- Ceyhan, Y.; Zhang, M.; Sandoval, C.G.; Agoulnik, A.I.; Agoulnik, I.U. Expression patterns and the roles of phosphatidylinositol phosphatases in testisdagger. Biol. Reprod. 2022, 107, 902–915. [Google Scholar]
- Hanzu, F.A.; Gasa, R.; Bulur, N.; Lybaert, P.; Gomis, R.; Malaisse, W.J.; Beauwens, R.; Sener, A. Expression of tmem16a and slc4a4 in human pancreatic islets. Cell. Physiol. Biochem. 2012, 29, 61–64. [Google Scholar] [CrossRef]
- Jiang, J.; Ma, L.; Prakapenka, D.; VanRaden, P.M.; Cole, J.B.; Da, Y. A large-scale genome-wide association study in u.s. Holstein cattle. Front. Genet. 2019, 10, 412. [Google Scholar] [CrossRef]
- Grzeskowiak, L.; Saliu, E.M.; Martinez-Vallespin, B.; Aschenbach, J.R.; Brockmann, G.A.; Fulde, M.; Hartmann, S.; Kuhla, B.; Lucius, R.; Metges, C.C.; et al. Dietary fiber and its role in performance, welfare, and health of pigs. Anim. Health Res. Rev. 2022, 23, 165–193. [Google Scholar] [CrossRef]
- Kim, M.; Roura, E.; Choi, Y.; Kim, J. Transcriptomic analysis of the porcine gut in response to heat stress and dietary soluble fiber from beet pulp. Genes 2022, 13, 1456. [Google Scholar] [CrossRef]
- Li, J. Study of Porcine CACNA2D1 Gene; Zhejiang University: Hangzhou, China, 2007. (In Chinese) [Google Scholar]
- Akit, H.; Collins, C.; Fahri, F.; Hung, A.; D’Souza, D.; Leury, B.; Dunshea, F. Dietary lecithin decreases skeletal muscle col1a1 and col3a1 gene expression in finisher gilts. Animals 2016, 6, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, T.F.; Braga, M.A.; Verardo, L.L.; Santos, G.C.; Silva, F.A.; Gomes, V.J.; Irano, N.; Dos, S.D. Functional analysis of litter size and number of teats in pigs: From gwas to post-gwas. Theriogenology 2022, 193, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Farmanullah, F.; Hosseini, S.M.; Liang, A.; Hua, G.; Rehman, Z.U.; Talpur, H.S.; Salim, M.; Ahmad, S.; Abulaiti, A.; Khan, M.; et al. Adaptive molecular evolution of akt3 gene for positive diversifying selection in mammals. Biomed Res. Int. 2020, 2020, 2584627. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Tian, S.; Jin, L.; Zhou, G.; Li, Y.; Zhang, Y.; Wang, T.; Yeung, C.K.; Chen, L.; Ma, J.; et al. Genomic analyses identify distinct patterns of selection in domesticated pigs and tibetan wild boars. Nat. Genet. 2013, 45, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Zhang, L.; Biswas, S.; Schugar, R.C.; Brown, J.M.; Byzova, T.; Podrez, E. Akt3 inhibits adipogenesis and protects from diet-induced obesity via wnk1/sgk1 signaling. JCI Insight 2017, 2, e95687. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; An, J.; Lian, L.; Qu, L.; Zheng, J.; Xu, G.; Yang, N. Polymorphisms in akt3, figf, prkag3, and tgf-beta genes are associated with myofiber characteristics in chickens. Poult. Sci. 2013, 92, 325–330. [Google Scholar] [CrossRef]
- Jung, W.; Yoo, I.; Han, J.; Kim, M.; Lee, S.; Cheon, Y.; Hong, M.; Jeon, B.Y.; Ka, H. Expression of caspases in the pig endometrium throughout the estrous cycle and at the maternal-conceptus interface during pregnancy and regulation by steroid hormones and cytokines. Front. Vet. Sci. 2021, 8, 641916. [Google Scholar] [CrossRef]
- Petrelli, S.; Buglione, M.; Maselli, V.; Troiano, C.; Larson, G.; Frantz, L.; Manin, A.; Ricca, E.; Baccigalupi, L.; Wright, D.; et al. Population genomic, olfactory, dietary, and gut microbiota analyses demonstrate the unique evolutionary trajectory of feral pigs. Mol. Ecol. 2022, 31, 220–237. [Google Scholar] [CrossRef]
- Gao, N.; Lu, M.; Echeverri, F.; Laita, B.; Kalabat, D.; Williams, M.E.; Hevezi, P.; Zlotnik, A.; Moyer, B.D. Voltage-gated sodium channels in taste bud cells. BMC Neurosci. 2009, 10, 20. [Google Scholar] [CrossRef] [Green Version]
- Haehner, A.; Hummel, T.; Heinritz, W.; Krueger, S.; Meinhardt, M.; Whitcroft, K.L.; Sabatowski, R.; Gossrau, G. Mutation in na(v) 1.7 causes high olfactory sensitivity. Eur. J. Pain 2018, 22, 1767–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, R.Y.; Ham, J.R.; Lee, M.K. Esculetin prevents non-alcoholic fatty liver in diabetic mice fed high-fat diet. Chem. Biol. Interact. 2016, 260, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Busnelli, M.; Manzini, S.; Parolini, C.; Escalante-Alcalde, D.; Chiesa, G. Lipid phosphate phosphatase 3 in vascular pathophysiology. Atherosclerosis 2018, 271, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Luo, Y.; Zhang, L.; Li, J.; Zhang, B.; Xing, S.; Zhu, Y.; Gao, F.; Zhou, G. Effects of cysteamine supplementation on the intestinal expression of amino acid and peptide transporters and intestinal health in finishing pigs. Anim. Sci. J. 2017, 88, 314–321. [Google Scholar] [CrossRef]
- Gotoh, S.; Miyauchi, Y.; Moore, R.; Negishi, M. Glucose elicits serine/threonine kinase vrk1 to phosphorylate nuclear pregnane x receptor as a novel hepatic gluconeogenic signal. Cell. Signal. 2017, 40, 200–209. [Google Scholar] [CrossRef]
- Bosma, P.J.; Wits, M.; Oude-Elferink, R.P. Gene therapy for progressive familial intrahepatic cholestasis: Current progress and future prospects. Int. J. Mol. Sci. 2020, 22, 273. [Google Scholar] [CrossRef]
- Puccinelli, E.; Gervasi, P.G.; Trivella, M.G.; Vornoli, A.; Viglione, F.; Pelosi, G.; Parodi, O.; Sampietro, T.; Puntoni, M. Modulation of lipid homeostasis in response to continuous or intermittent high-fat diet in pigs. Animal 2015, 9, 1000–1007. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.L.; Zheng, W.; Ren, P.; Hu, H.; Tong, X.; Zhang, S.P.; Li, X.; Wang, H.; Jiang, J.C.; Jin, J.; et al. Biallelic mutations in mos cause female infertility characterized by human early embryonic arrest and fragmentation. Embo Mol. Med. 2021, 13, e14887. [Google Scholar] [CrossRef]
- Fei, C.F.; Zhou, L.Q. Gene mutations impede oocyte maturation, fertilization, and early embryonic development. Bioessays 2022, 44, e2200007. [Google Scholar] [CrossRef]
- Celozzi, S.; Battini, M.; Prato-Previde, E.; Mattiello, S. Humans and goats: Improving knowledge for a better relationship. Animals 2022, 12, 774. [Google Scholar] [CrossRef]
- Samet, L.E.; Vaterlaws-Whiteside, H.; Harvey, N.D.; Upjohn, M.M.; Casey, R.A. Exploring and developing the questions used to measure the human-dog bond: New and existing themes. Animals 2022, 12, 805. [Google Scholar] [CrossRef] [PubMed]
- Norkett, R.; Modi, S.; Birsa, N.; Atkin, T.A.; Ivankovic, D.; Pathania, M.; Trossbach, S.V.; Korth, C.; Hirst, W.D.; Kittler, J.T. Disc1-dependent regulation of mitochondrial dynamics controls the morphogenesis of complex neuronal dendrites. J. Biol. Chem. 2016, 291, 613–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; O’Brien, D.P.; Mhlanga-Mutangadura, T.; Olby, N.J.; Taylor, J.F.; Schnabel, R.D.; Katz, M.L.; Johnson, G.S. A rare homozygous mfsd8 single-base-pair deletion and frameshift in the whole genome sequence of a Chinese crested dog with neuronal ceroid lipofuscinosis. BMC Vet. Res. 2015, 10, 960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Derks, M.F.; Groenen, M.A.; Zhao, Y.; Bosse, M. Distinct traces of mixed ancestry in western commercial pig genomes following gene flow from Chinese indigenous breeds. Front. Genet. 2022, 13, 1070783. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Zhang, H.; Long, H.; Zhang, D.; Yang, X.; Liu, D.; E, G. Genome-Wide Selection Signal Analysis to Investigate Wide Genomic Heredity Divergence between Eurasian Wild Boar and Domestic Pig. Animals 2023, 13, 2158. https://doi.org/10.3390/ani13132158
Wu X, Zhang H, Long H, Zhang D, Yang X, Liu D, E G. Genome-Wide Selection Signal Analysis to Investigate Wide Genomic Heredity Divergence between Eurasian Wild Boar and Domestic Pig. Animals. 2023; 13(13):2158. https://doi.org/10.3390/ani13132158
Chicago/Turabian StyleWu, Xinming, Haoyuan Zhang, Haoyuan Long, Dongjie Zhang, Xiuqin Yang, Di Liu, and Guangxin E. 2023. "Genome-Wide Selection Signal Analysis to Investigate Wide Genomic Heredity Divergence between Eurasian Wild Boar and Domestic Pig" Animals 13, no. 13: 2158. https://doi.org/10.3390/ani13132158