
Automatically Identifying Sickness Behavior in Grazing Lambs with an Acceleration Sensor

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site and Design
2.2. Animals and Management
2.3. Measurements
2.3.1. Infection Status
2.3.2. Herbage Mass
2.3.3. Behavior
3. Results
3.1. Rectal Temperature
3.2. Herbage Mass
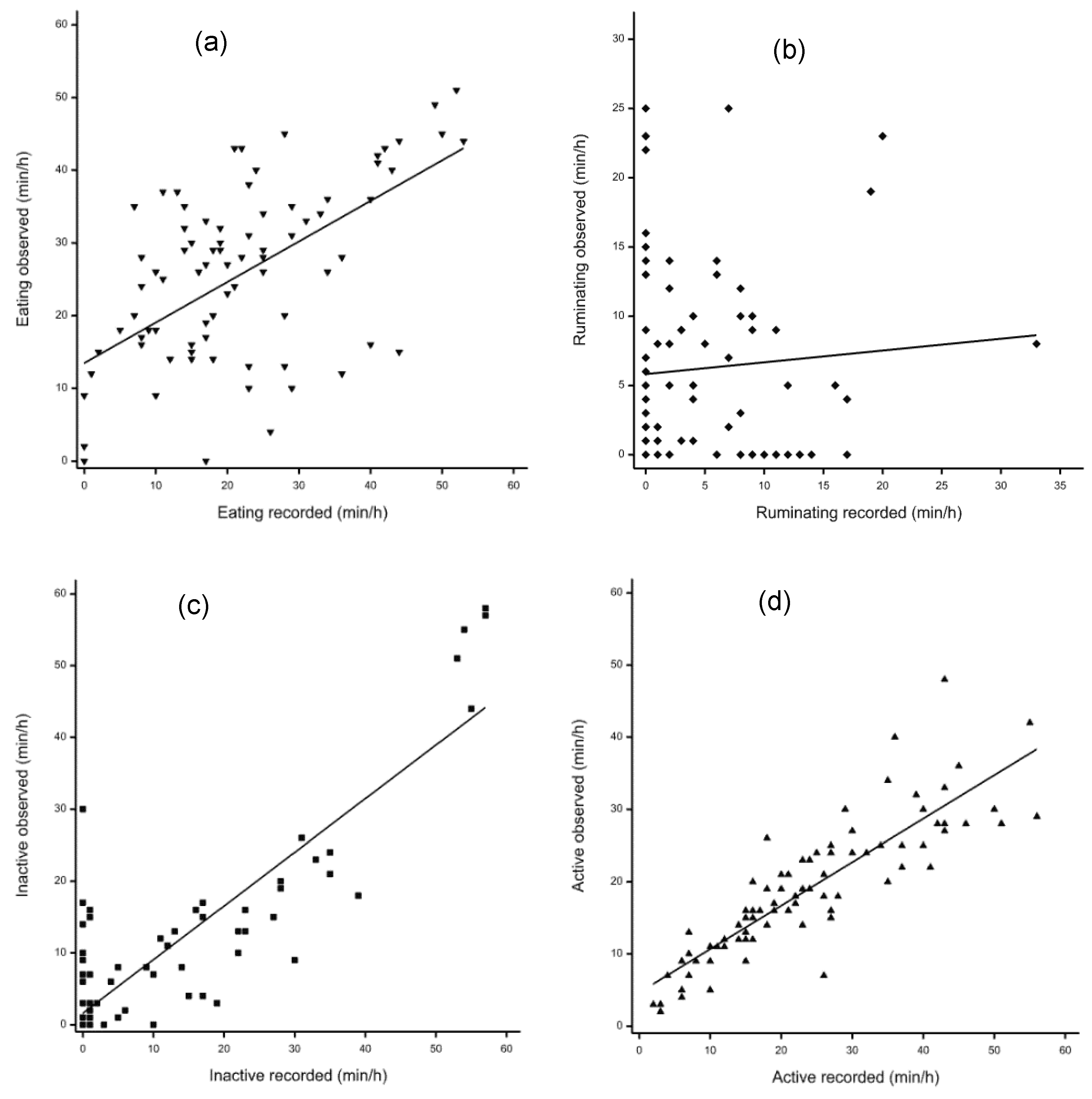
3.3. Validation for the Data between Visual Observation and Sensor Detection
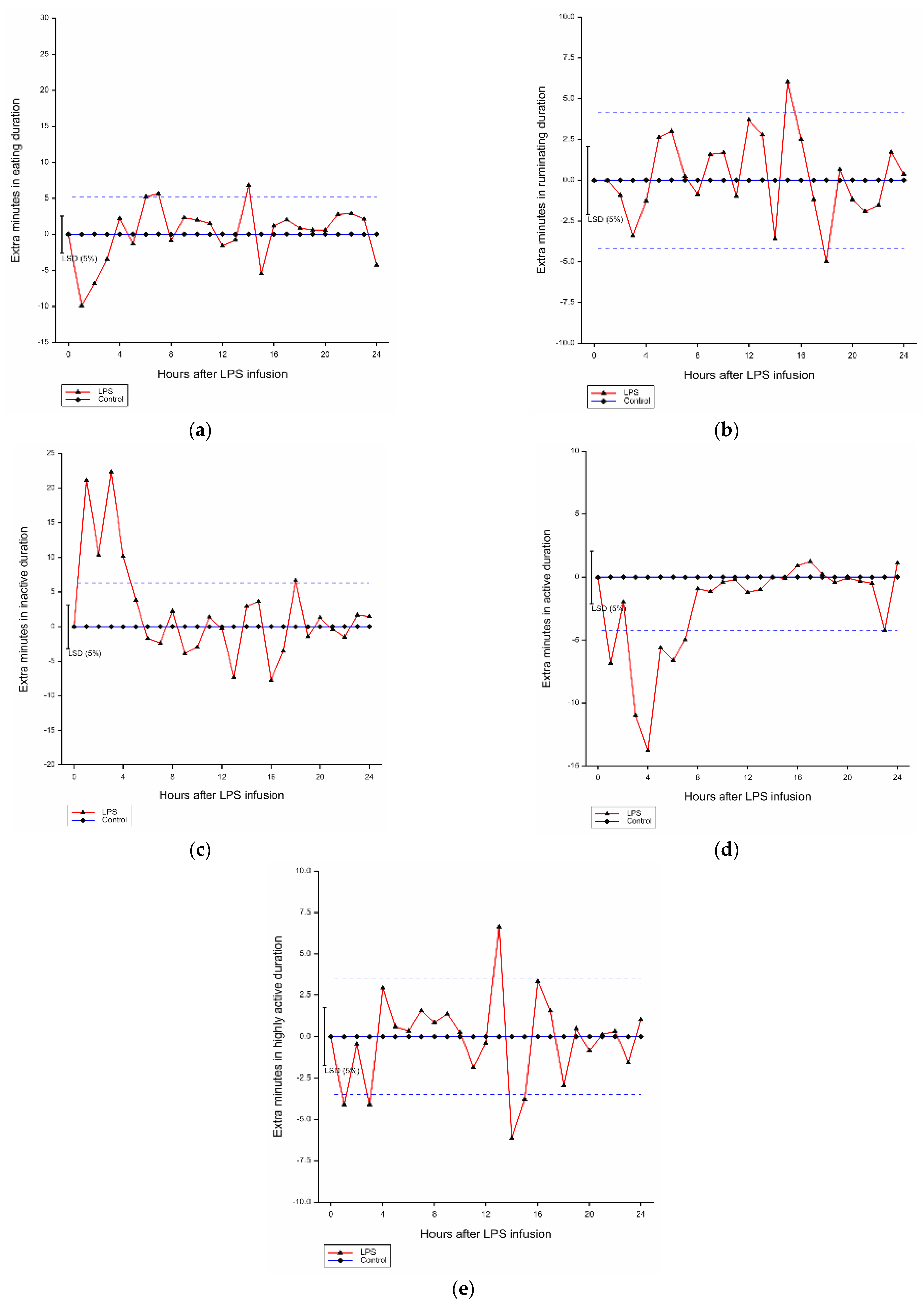
3.4. Effects of LPS Challenge on Activity Patterns
3.4.1. Eating Duration
3.4.2. Ruminating Duration
3.4.3. Inactive Duration
3.4.4. Active Duration
3.4.5. Highly Active Duration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steiger, M.; Senn, M.; Altreuther, G.; Werling, D.; Sutter, F.; Kreuzer, M.; Langhans, W. Effect of a prolonged low-dose lipopolysaccharide infusion on feed intake and metabolism in heifers. J. Anim. Sci. 1999, 77, 2523–2532. [Google Scholar] [CrossRef] [PubMed]
- Kopf, B.S.; Langhans, W.; Geary, N.; Asarian, L. Serotonin 2C receptor signaling in a diffuse neuronal network is necessary for LPS anorexia. Brain Res. 2010, 1306, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Breen, G.; Barrell, G. Effects of pregnancy on the fever response of sheep to a Gram-negative pyrogen. Proc. N. Z. Soc. Anim. Prod. 2002, 62, 337–339. [Google Scholar]
- Fisher, R.E.; Karrow, N.A.; Quinton, M.; Finegan, E.J.; Miller, S.P.; Atkinson, J.L.; Boermans, H.J. Endotoxin exposure during late pregnancy alters ovine offspring febrile and hypothalamic–pituitary–adrenal axis responsiveness later in life. Stress 2010, 13, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Plessers, E.; Wyns, H.; Watteyn, A.; Pardon, B.; De Backer, P.; Croubels, S. Characterization of an intravenous lipopolysaccharide inflammation model in calves with respect to the acute-phase response. Vet. Immunol. Immunopathol. 2015, 163, 46–56. [Google Scholar] [CrossRef]
- Waldron, M.; Nishida, T.; Nonnecke, B.; Overton, T. Effect of lipopolysaccharide on indices of peripheral and hepatic metabolism in lactating cows. J. Dairy Sci. 2003, 86, 3447–3459. [Google Scholar] [CrossRef]
- Werling, D.; Sutter, F.; Arnold, M.; Kun, G.; Tooten, P.; Gruys, E.; Kreuzer, M.; Langhans, W. Characterisation of the acute phase response of heifers to a prolonged low dose infusion of lipopolysaccharide. Res. Vet. Sci. 1996, 61, 252–257. [Google Scholar] [CrossRef]
- McMahon, C.; Buxton, D.; Elsasser, T.; Gunter, D.; Sanders, L.; Steele, B.; Sartin, J. Neuropeptide Y restores appetite and alters concentrations of GH after central administration to endotoxic sheep. J. Endocrinol. 1999, 161, 333–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartin, J.; Marks, D.; McMahon, C.; Daniel, J.; Levasseur, P.; Wagner, C.; Whitlock, B.; Steele, B. Central role of the melanocortin-4 receptors in appetite regulation after endotoxin. J. Anim. Sci. 2008, 86, 2557–2567. [Google Scholar] [CrossRef]
- Aditya, S.; Humer, E.; Pourazad, P.; Khiaosa-Ard, R.; Huber, J.; Zebeli, Q. Intramammary infusion of Escherichia coli lipopolysaccharide negatively affects feed intake, chewing, and clinical variables, but some effects are stronger in cows experiencing subacute rumen acidosis. J. Dairy Sci. 2017, 100, 1363–1377. [Google Scholar] [CrossRef] [Green Version]
- Siivonen, J.; Taponen, S.; Hovinen, M.; Pastell, M.; Lensink, B.J.; Pyörälä, S.; Hänninen, L. Impact of acute clinical mastitis on cow behaviour. Appl. Anim. Behav. Sci. 2011, 132, 101–106. [Google Scholar] [CrossRef]
- Müller, R.; Schrader, L. A new method to measure behavioural activity levels in dairy cows. Appl. Anim. Behav. Sci. 2003, 83, 247–258. [Google Scholar] [CrossRef]
- Fan, B.; Bryant, R.; Greer, A. Behavioral Fingerprinting: Acceleration Sensors for Identifying Changes in Livestock Health. J 2022, 5, 435–454. [Google Scholar] [CrossRef]
- Fogarty, E.S.; Swain, D.L.; Cronin, G.; Trotter, M. Autonomous on-animal sensors in sheep research: A systematic review. Comput. Electron. Agric. 2018, 150, 245–256. [Google Scholar] [CrossRef]
- Wolfger, B.; Timsit, E.; Pajor, E.; Cook, N.; Barkema, H.; Orsel, K. Accuracy of an ear tag-attached accelerometer to monitor rumination and feeding behavior in feedlot cattle. J. Anim. Sci. 2015, 93, 3164–3168. [Google Scholar] [CrossRef] [Green Version]
- Bikker, J.P.; van Laar, H.; Rump, P.; Doorenbos, J.; van Meurs, K.; Griffioen, G.M.; Dijkstra, J. Technical note: Evaluation of an ear-attached movement sensor to record cow feeding behavior and activity. J. Dairy Sci. 2014, 97, 2974–2979. [Google Scholar] [CrossRef]
- Borchers, M.R.; Chang, Y.M.; Tsai, I.C.; Wadsworth, B.A.; Bewley, J.M. A validation of technologies monitoring dairy cow feeding, ruminating, and lying behaviors. J. Dairy Sci. 2016, 99, 7458–7466. [Google Scholar] [CrossRef]
- Reynolds, M.A.; Borchers, M.R.; Davidson, J.A.; Bradley, C.M.; Bewley, J.M. Technical note: An evaluation of technology-recorded rumination and feeding behaviors in dairy heifers. J. Dairy Sci. 2019, 102, 6555–6558. [Google Scholar] [CrossRef]
- Dolecheck, K.A.; Silvia, W.J.; Heersche, G.; Chang, Y.M.; Ray, D.L.; Stone, A.E.; Wadsworth, B.A.; Bewley, J.M. Behavioral and physiological changes around estrus events identified using multiple automated monitoring technologies. J. Dairy Sci. 2015, 98, 8723–8731. [Google Scholar] [CrossRef]
- Rutten, C.J.; Kamphuis, C.; Hogeveen, H.; Huijps, K.; Nielen, M.; Steeneveld, W. Sensor data on cow activity, rumination, and ear temperature improve prediction of the start of calving in dairy cows. Comput. Electron. Agric. 2017, 132, 108–118. [Google Scholar] [CrossRef]
- VanValin, K.R.; Carmichael-Wyatt, R.N.; Deters, E.L.; Messersmith, E.M.; Heiderscheit, K.J.; Hochmuth, K.G.; Jackson, T.D.; Peschel, J.M.; Johnson, A.K.; Hansen, S.L. Dietary zinc concentration and lipopolysaccharide injection affect circulating trace minerals, acute phase protein response, and behavior as evaluated by an ear-tag–based accelerometer in beef steers. J. Anim. Sci. 2021, 99, skab278. [Google Scholar] [CrossRef] [PubMed]
- Borderas, T.; De Passillé, A.; Rushen, J. Behavior of dairy calves after a low dose of bacterial endotoxin. J. Anim. Sci. 2008, 86, 2920–2927. [Google Scholar] [CrossRef]
- Pereira, G.M.; Heins, B.J.; Endres, M.I. Technical note: Validation of an ear-tag accelerometer sensor to determine rumination, eating, and activity behaviors of grazing dairy cattle. J. Dairy Sci. 2018, 101, 2492–2495. [Google Scholar] [CrossRef] [Green Version]
- Zambelis, A.; Wolfe, T.; Vasseur, E. Technical note: Validation of an ear-tag accelerometer to identify feeding and activity behaviors of tiestall-housed dairy cattle. J. Dairy Sci. 2019, 102, 4536–4540. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, P.P. Automatic registration of grazing behaviour in dairy cows using 3D activity loggers. Appl. Anim. Behav. Sci. 2013, 148, 179–184. [Google Scholar] [CrossRef]
- Elsasser, T.; Caperna, T.; Rumsey, T. Endotoxin administration decreases plasma insulin-like growth factor (IGF)-I and IGF-binding protein-2 in Angus× Hereford steers independent of changes in nutritional intake. J. Endocrinol. 1995, 144, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro Filho, H.M.N.; Setelich, E.A.; Crestani, S.; Dias, K.M.; Mantovani, C.; Valenti, J. Relationship between diurnal grazing time and herbage intake in dairy cows in rotational grazing. Ciência Rural 2011, 41, 2010–2013. [Google Scholar] [CrossRef] [Green Version]
- Allden, W.; McDWhittaker, I. The determinants of herbage intake by grazing sheep: The interrelationship of factors influencing herbage intake and availability. Aust. J. Agric. Res. 1970, 21, 755–766. [Google Scholar] [CrossRef]
- González, L.A.; Tolkamp, B.J.; Coffey, M.P.; Ferret, A.; Kyriazakis, I. Changes in Feeding Behavior as Possible Indicators for the Automatic Monitoring of Health Disorders in Dairy Cows. J. Dairy Sci. 2008, 91, 1017–1028. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, Y.; Kikusui, T.; Mori, Y. Changes in the behavioral parameters following the lipopolysaccharide administration in goats. J. Vet. Med. Sci. 1995, 57, 1041–1044. [Google Scholar] [CrossRef] [Green Version]
- Szyszka, O.; Tolkamp, B.; Edwards, S.; Kyriazakis, I. The effects of acute versus chronic health challenges on the behavior of beef cattle. J. Anim. Sci. 2012, 90, 4308–4318. [Google Scholar] [CrossRef]
- Nordgreen, J.; Munsterhjelm, C.; Aae, F.; Popova, A.; Boysen, P.; Ranheim, B.; Heinonen, M.; Raszplewicz, J.; Piepponen, P.; Lervik, A.; et al. The effect of lipopolysaccharide (LPS) on inflammatory markers in blood and brain and on behavior in individually-housed pigs. Physiol. Behav. 2018, 195, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Owen-Ashley, N.T.; Turner, M.; Hahn, T.P.; Wingfield, J.C. Hormonal, behavioral, and thermoregulatory responses to bacterial lipopolysaccharide in captive and free-living white-crowned sparrows (Zonotrichia leucophrys gambelii). Horm. Behav. 2006, 49, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Ferreira Mello, B.S.; Monte, A.S.; McIntyre, R.S.; Soczynska, J.K.; Custódio, C.S.; Cordeiro, R.C.; Chaves, J.H.; Mendes Vasconcelos, S.M.; Nobre Júnior, H.V.; Florenço de Sousa, F.C.; et al. Effects of doxycycline on depressive-like behavior in mice after lipopolysaccharide (LPS) administration. J. Psychiatr. Res. 2013, 47, 1521–1529. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, Q.; Yuan, L.; Wang, S.; Liu, L.; Yang, X.; Li, G.; Liu, D. The effects of curcumin on depressive-like behavior in mice after lipopolysaccharide administration. Behav. Brain Res. 2014, 274, 282–290. [Google Scholar] [CrossRef]
- Bian, H.; Wang, G.; Huang, J.; Liang, L.; Zheng, Y.; Wei, Y.; Wang, H.; Xiao, L.; Wang, H. Dihydrolipoic acid protects against lipopolysaccharide-induced behavioral deficits and neuroinflammation via regulation of Nrf2/HO-1/NLRP3 signaling in rat. J. Neuroinflammation 2020, 17, 166. [Google Scholar] [CrossRef]
- Rodrigues, F.T.S.; de Souza, M.R.M.; Lima, C.N.d.C.; da Silva, F.E.R.; Costa, D.V.d.S.; dos Santos, C.C.; Miyajima, F.; de Sousa, F.C.F.; Vasconcelos, S.M.M.; Barichello, T.; et al. Major depression model induced by repeated and intermittent lipopolysaccharide administration: Long-lasting behavioral, neuroimmune and neuroprogressive alterations. J. Psychiatr. Res. 2018, 107, 57–67. [Google Scholar] [CrossRef]
- Harden, L.M.; du Plessis, I.; Poole, S.; Laburn, H.P. Interleukin-6 and leptin mediate lipopolysaccharide-induced fever and sickness behavior. Physiol. Behav. 2006, 89, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Harden, L.M.; du Plessis, I.; Roth, J.; Loram, L.C.; Poole, S.; Laburn, H.P. Differences in the relative involvement of peripherally released interleukin (IL)-6, brain IL-1β and prostanoids in mediating lipopolysaccharide-induced fever and sickness behavior. Psychoneuroendocrinology 2011, 36, 608–622. [Google Scholar] [CrossRef]
- Trieu, L.L.; Bailey, D.W.; Cao, H.; Son, T.C.; Scobie, D.R.; Trotter, M.G.; Hume, D.E.; Sutherland, B.L.; Tobin, C.T. Potential of Accelerometers and GPS Tracking to Remotely Detect Perennial Ryegrass Staggers in Sheep. Smart Agric. Technol. 2022, 2, 100040. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Eating | Ruminating | Inactive | Active | |
|---|---|---|---|---|
| Visual observation | 25.39 ± 1.35 | 6.18 ± 0.73 | 9.33 ± 1.47 | 19.08 ± 1.03 |
| Sensor recordings | 21.38 ± 1.43 | 4.23 ± 0.66 | 10.36 ± 1.70 | 23.99 ± 1.48 |
| p-value for t-test | 0.002 | 0.043 | 0.23 | <0.001 |
| r | 0.59 | 0.08 | 0.87 | 0.86 |
| p-value for r | <0.001 | 0.48 | <0.001 | <0.001 |
| R2 | 0.35 (p < 0.001) | 0.006 (p = 0.48) | 0.75 (p < 0.001) | 0.75 (p < 0.001) |
| Slope (SEM, p) | 0.56 (0.08, p < 0.001) | 0.085 (0.12, p = 0.48) | 0.75 (0.05, p < 0.001) | 0.60 (0.04, p < 0.001) |
| Intercept (SEM, p) | 13.47 (2.12, p < 0.001) | 5.82 (0.90, p < 0.001) | 1.60 (0.88, p = 0.07) | 4.65 (1.06, p < 0.001) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, B.; Bryant, R.H.; Greer, A.W. Automatically Identifying Sickness Behavior in Grazing Lambs with an Acceleration Sensor. Animals 2023, 13, 2086. https://doi.org/10.3390/ani13132086
Fan B, Bryant RH, Greer AW. Automatically Identifying Sickness Behavior in Grazing Lambs with an Acceleration Sensor. Animals. 2023; 13(13):2086. https://doi.org/10.3390/ani13132086
Chicago/Turabian StyleFan, Bowen, Racheal H. Bryant, and Andrew W. Greer. 2023. "Automatically Identifying Sickness Behavior in Grazing Lambs with an Acceleration Sensor" Animals 13, no. 13: 2086. https://doi.org/10.3390/ani13132086