
Anthelmintic Treatment and the Stability of Parasite Distribution in Ruminants
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
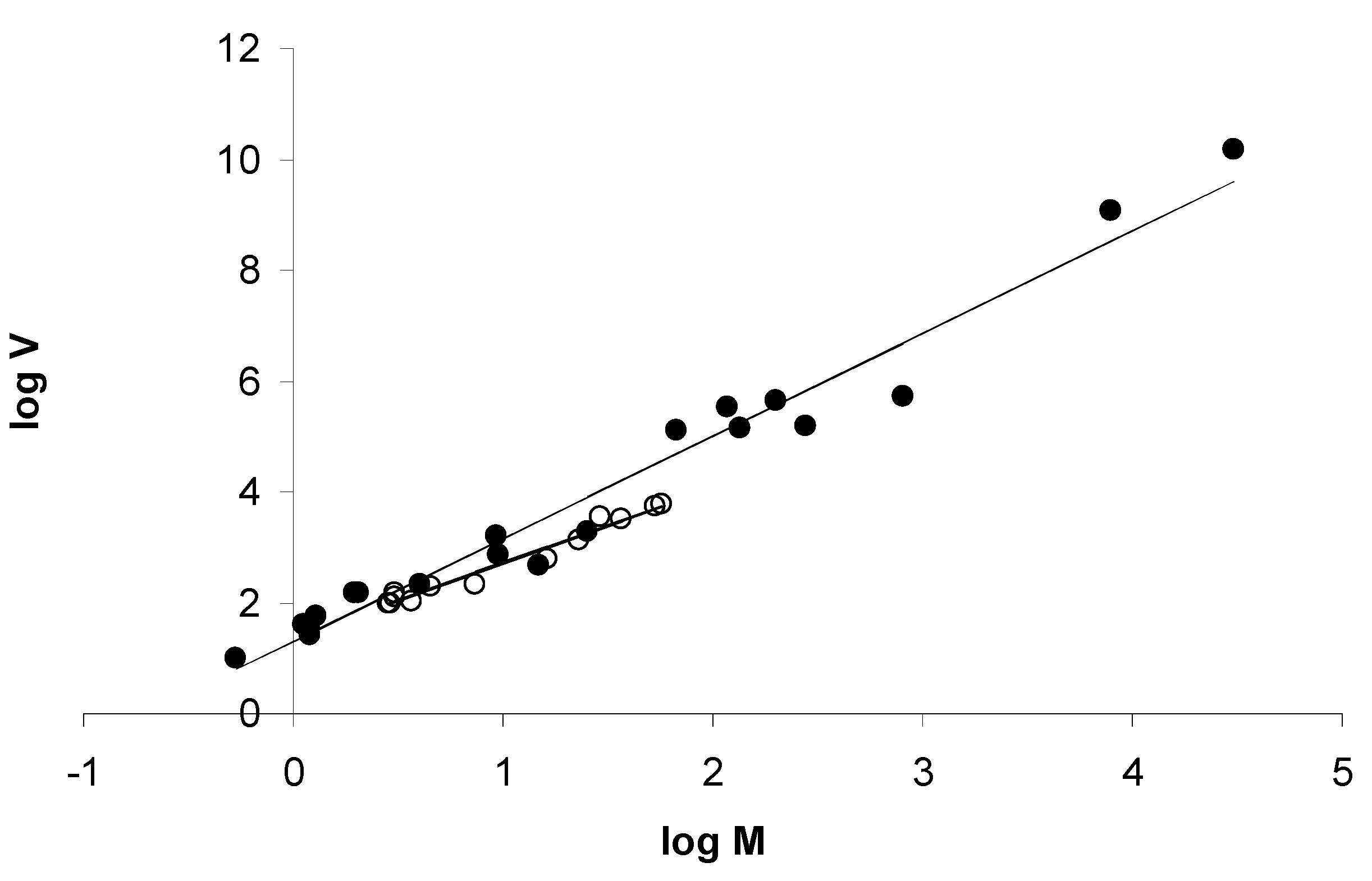
3.1. Overall Fit to Taylor’s Power Law
3.2. Immunity: Nematodirus and Marshallagia in Sheep and Antelopes
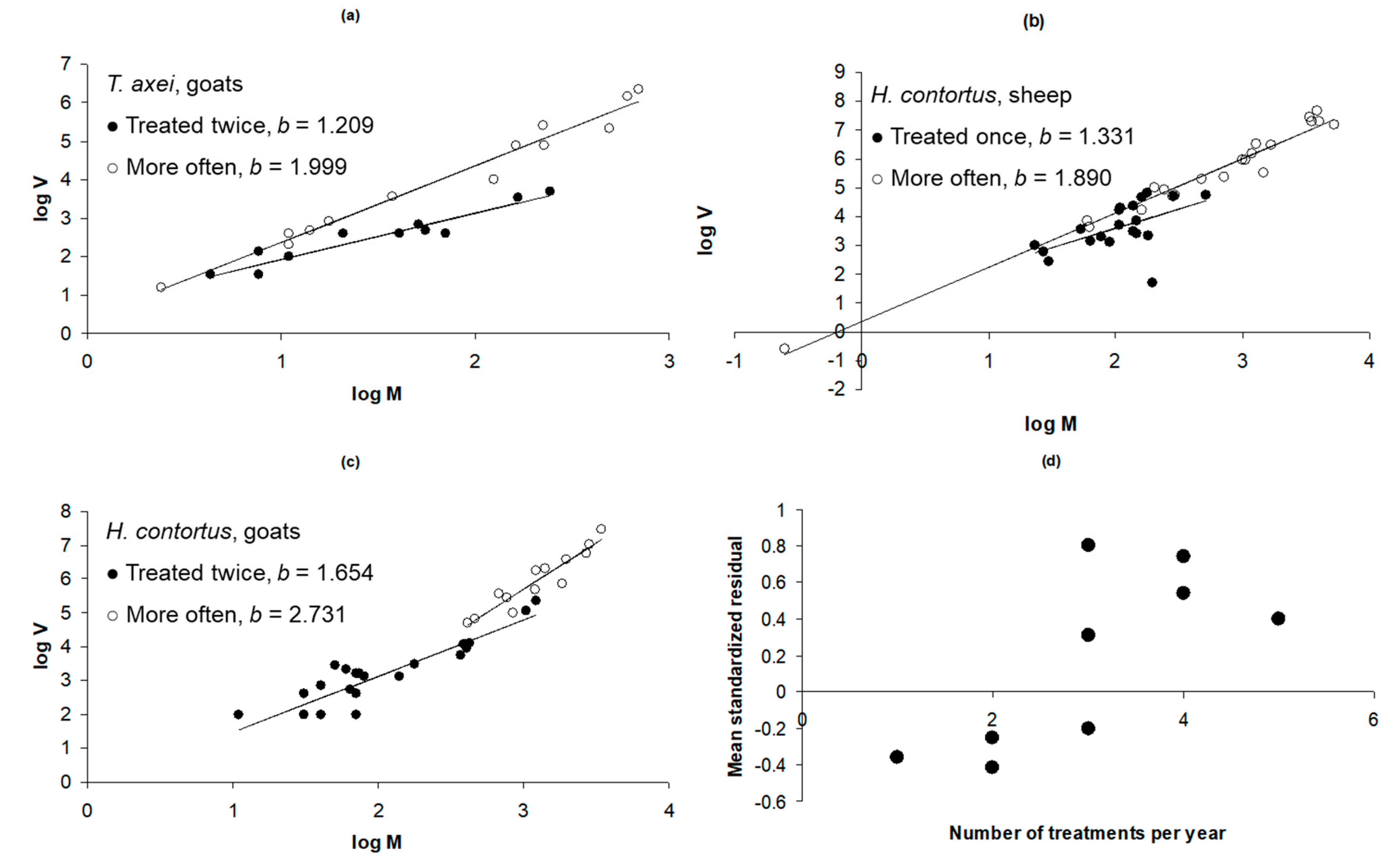
3.3. Frequency of Treatment: Haemonchus and Trichostrongylus in Lambs and Goats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eisler, Z.; Bartos, I.; Kertesz, J. Fluctuation scaling in complex systems: Taylor’s law and beyond. Adv. Phys. 2008, 57, 89–142. [Google Scholar] [CrossRef] [Green Version]
- Fairfield Smith, H. An empirical law describing heterogeneity in the yields of agricultural crops. J. Agric. Sci. 1938, 28, 1–23. [Google Scholar] [CrossRef]
- Taylor, L.R. Aggregation, variance and the mean. Nature 1961, 189, 732–735. [Google Scholar] [CrossRef]
- Barger, I.A. The statistical distribution of trichostrongylid nematodes in grazing lambs. Int. J. Parasitol. 1985, 15, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Gaba, S.; Ginot, V.; Cabaret, J. Modelling macroparasite aggregation using a nematode-sheep system: The Weibull distribution as an alternative to the Negative Binomial distribution? Parasitology 2005, 131, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.M.; May, R.M. Regulation and stability of host-parasite population interactions. 1. Regulatory processes. J. Anim. Ecol. 1978, 47, 219–247. [Google Scholar] [CrossRef]
- Quinnell, R.J.; Medley, G.F.; Keymer, A.E. The regulation of gastrointestinal helminth populations. Philos. Trans. R. Soc. Lond. B 1990, 330, 191–201. [Google Scholar]
- Medley, G.F.; Guyatt, H.L.; Bundy, D.A.P. A quantitative framework for evaluating the effect of community treatment on the morbidity due to ascariasis. Parasitology 1993, 106, 211–221. [Google Scholar] [CrossRef]
- Churcher, T.S.; Ferguson, N.M.; Basanez, M.G. Density dependence and overdispersion in the transmission of helminth parasites. Parasitology 2005, 131, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Morgan, E.R.; Cavill, L.; Curry, G.E.; Wood, R.M.; Mitchell, E.S.E. Effects of aggregation and sample size on composite faecal egg counts in sheep. Vet. Parasitol. 2005, 131, 79–87. [Google Scholar] [CrossRef]
- Churcher, T.S.; Basanez, M.G. Density dependence and the spread of anthelmintic resistance. Evolution 2008, 62, 528–537. [Google Scholar] [CrossRef]
- Charlier, J.; Morgan, E.R.; Rinalid, L.; Van Dijk, J.; Demeler, J.; Hoglund, J.; Hertzberg, H.; Van Ranst, B.; Hendrickx, G.; Vercruysse, J.; et al. Practices to optimise gastrointestinal nematode control on sheep, goat and cattle farms in Europe using targeted (selective) treatments. Vet. Rec. 2014, 175, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Michel, J.E. Strategies for the use of anthelmintics in livestock and their implications for the development of drug resistance. Parasitology 1985, 90, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Van Wyk, J.A.; Bath, G.F. The FAMACHA system for managing haemonchosis in sheep and goats by clinically identifying individual animals for treatment. Vet. Res. 2002, 33, 509–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrill, A.; Forbes, M.R. Random parasite encounters coupled with condition-linked immunity of hosts generate parasite aggregation. Int. J. Parasitol. 2012, 42, 701–706. [Google Scholar] [CrossRef]
- Johnson, P.T.J.; Hoverman, J.T. Heterogeneous hosts: How variation in host size, behaviour and immunity affects parasite aggregation. J. Anim. Ecol. 2014, 83, 1103–1112. [Google Scholar] [CrossRef]
- Tinsley, R.; Vineer, H.R.; Grainger-Wood, R.; Morgan, E.R. Heterogeneity in helminth infections: Factors influencing aggregation in a simple host-parasite system. Parasitology 2020, 147, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Grenfell, B.T.; Wilson, K.; Isham, V.S.; Boyd, H.E.G.; Dietz, K. Modelling patterns of parasite aggregation in natural populations: Trichostrongylid nematode-ruminant interactions as a case study. Parasitology 1995, 111, S135–S151. [Google Scholar] [CrossRef] [Green Version]
- Galvani, A.P. Immunity, heterogeneity, and aggregation of helminth parasites. J. Parasitol. 2003, 89, 232–241. [Google Scholar] [CrossRef]
- Cornell, S.J.; Isham, V.S.; Grenfell, B.T. Stochastic and spatial dynamics of nematode parasites in farmed ruminants. Proc. R. Soc. B 2004, 271, 1243–1250. [Google Scholar] [CrossRef]
- Stromberg, B.E. Environmental factors influencing transmission. Vet. Parasitol. 1997, 72, 247–256. [Google Scholar] [CrossRef]
- Heinzmann, D.; Barbour, A.D.; Torgerson, P.R. Compound processes as models for clumped parasite data. Math. Biosci. 2009, 222, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Vineer, H.R.; Morgan, E.R.; Hertzberg, H.; Bartley, D.J.; Bosco, A.; Charlier, J.; Chartier, C.; Claerebout, E.; de Waal, T.; Hendrickx, G.; et al. Increasing importance of anthelmintic resistance in European livestock: Creation and meta-analysis of an open database. Parasite 2020, 27, 69. [Google Scholar] [CrossRef]
- Vercruysse, J.; Charlier, J.; Van Dijk, J.; Morgan, E.R.; Geary, T.; von Samson-Himmelstjerna, G.; Claerebout, E. Control of helminth ruminant infections by 2030. Parasitology 2018, 145, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- Poulin, R.; Morand, S. Parasite body size and interspecific variation in levels of aggregation among nematodes. J. Parasitol. 2000, 86, 642–647. [Google Scholar] [CrossRef]
- Filipe, J.A.N.; Kyriazakis, I.; McFarland, C.; Morgan, E.R. Novel epidemiological model of gastrointestinal nematode infection to assess grazing cattle resilience by integrating host growth, parasite, grass and environmental dynamics. Int. J. Parasitol. 2023, 53, 133–155. [Google Scholar] [CrossRef]
- Grenfell, B.; Harwood, J. (Meta)population dynamics of infectious diseases. Trends Ecol. Evol. 1997, 12, 395–399. [Google Scholar] [CrossRef]
- Benton, T.G.; Beckerman, A.P. Population dynamics in a noisy world: Lessons from a mite experimental system. Adv. Ecol. Res. 2005, 37, 143–181. [Google Scholar]
- Morand, S.; Krasnov, B. Why apply ecological laws to epidemiology? Trends Parasitol. 2008, 24, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Blaxter, M.; Koutsovoulos, G. The evolution of parasitism in Nematoda. Nature 2015, 142, S26–S39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, E.R.; Shaikenov, B.; Torgerson, P.R.; Medley, G.F.; Milner-Gulland, E.J. Helminths of saiga antelopes in Kazakhstan: Implications for conservation and livestock production. J. Wildl. Dis. 2005, 41, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Morgan, E.R.; Torgerson, P.R.; Shaikenov, B.S.; Usenbayev, A.E.; Moore, A.B.M.; Medley, G.F.; Milner-Gulland, E.J. Agricultural restructuring and gastrointestinal parasitism in domestic ruminants on the rangelands of Kazakhstan. Vet. Parasitol. 2006, 139, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Nowosad, B.; Gruner, L.; Skalska, M.; Fudalewicz-Niemczyk, W.; Molenda, K.; Kornas, S. Genetic difference in natural resistance to gastrointestinal nematodes in Polish long-wool, Blackface and Weisses Alpenschaf sheep. Acta Parassitol. 2003, 48, 131–134. [Google Scholar]
- Gasnier, N.; Cabaret, J.; Chartier, C.; Reche, B. Species diversity in gastrointestinal nematode communities of dairy goats: Species-area and species-climate relationships. Vet. Res. 1997, 28, 55–64. [Google Scholar]
- Cabaret, J.; Gasnier, N.; Jacquiet, P. Faecal egg counts are representative of digestive-tract strongyle worm burdens in sheep and goats. Parasite 1998, 5, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Shaw, D.J.; Dobson, A.P. Patterns of macroparasite abundance and aggregation in wildlife populations: A quantitative review. Parasitology 1995, 111, S111–S133. [Google Scholar] [CrossRef] [PubMed]
- Shaw, D.J.; Grenfell, B.T.; Dobson, A.P. Patterns of macroparasite aggregation in wildlife host populations. Parasitology 1998, 117, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Claerebout, E.; Vercruysse, J. The immune response and the evaluation of acquired immunity against gastrointestinal nematodes in cattle: A review. Parasitology 2000, 120, S25–S42. [Google Scholar] [CrossRef]
- Boag, B.; Hackett, C.A.; Topham, P.B. The use of Taylor’s Power Law to describe the aggregated distribution of gastro-intestinal nematodes of sheep. Int. J. Parasitol. 1992, 22, 267–270. [Google Scholar] [CrossRef]
- Denwood, M.J.; Stear, M.J.; Matthews, L.; Reid, S.W.J.; Toft, N.; Innocent, G.T. The distribution of the pathogenic nematode Nematodirus battus in lambs is zero-inflated. Parasitology 2008, 135, 1225–1235. [Google Scholar] [CrossRef]
- Israf, D.A.; Jackson, F.; Stevenson, L.M.; Jones, D.G.; Jackson, E.; Huntley, J.F.; Coop, R.L. Persistence of immunity to Nematodirus battus infection in lambs. Vet. Parasitol. 1997, 71, 39–52. [Google Scholar] [CrossRef] [Green Version]
- Boag, B.; Thomas, R.J. Epidemiological studies on gastrointestinal nematode parasites of sheep—Seasonal number of generations and succession of species. Res. Vet. Sci. 1977, 22, 62–67. [Google Scholar] [CrossRef]
- Boag, B.; Lello, J.; Fenton, A.; Tompkins, D.M.; Hudson, P.J. Patterns of parasite aggregation in the wild European rabbit (Oryctolagus cuniculus). Int. J. Parasitol. 2001, 31, 1421–1428. [Google Scholar] [CrossRef] [PubMed]
- Boag, B.; Hernandez, A.D.; Cattadori, I.M. Observations on the epidemiology and interactions between myxomatosis, coccidiosis and helminth parasites in a wild rabbit population in Scotland. Eur. J. Wildl. Res. 2013, 59, 557–562. [Google Scholar] [CrossRef]
- Krasnov, B.R.; Stanko, M.; Miklisova, D.; Morand, S. Host specificity, parasite community size and the relation between abundance and its variance. Evol. Ecol. 2006, 20, 75–91. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Ives, A.R. Species interactions can explain Taylor’s power law for ecological time series. Nature 2003, 422, 6927. [Google Scholar] [CrossRef]
- Jimenez, J.J.; Rossi, J.P.; Lavelle, P. Spatial distribution of earthworms in acid-soil savannas of the eastern plains of Colombia. Appl. Soil Ecol. 2001, 17, 267–278. [Google Scholar] [CrossRef]
- He, F.; Gaston, K.J. Occupancy, spatial variance and the abundance of species. Am. Nat. 2003, 162, 366–375. [Google Scholar] [CrossRef] [Green Version]
- Collyer, B.S.; Anderson, R.M. Probability distributions of helminth parasite burdens within the human host population following repeated rounds of mass drug administration and their impact on the transmission breakpoint. J. R. Soc. Interface 2021, 18, 177. [Google Scholar] [CrossRef]
- Werkman, M.; Wright, J.E.; Truscott, J.E.; Oswald, W.E.; Halliday, K.E.; Papaiakovou, M.; Farrell, S.H.; Pullan, R.L.; Anderson, R.M. The impact of community-wide, mass drug administration on aggregation of soil-transmitted helminth infection in human host populations. Parasites Vectors 2020, 13, 290. [Google Scholar] [CrossRef]
- Engen, S.; Lande, R.; Saether, B.E. A general model for analyzing Taylor’s spatial scaling laws. Ecology 2008, 89, 2612–2622. [Google Scholar] [CrossRef] [Green Version]
- Woolhouse, M.E.J.; Taylor, L.H.; Haydon, D.T. Population biology of multihost pathogens. Science 2001, 292, 1109–1112. [Google Scholar] [CrossRef] [PubMed]
- Keeling, M.; Grenfell, B. Stochastic dynamics and a power law for measles variability. Philos. Trans. R. Soc. Lond. B 1999, 354, 769–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballantyne, F.; Kerkhoff, A.J. Reproductive correlation and mean-variance scaling of reproductive output for a forest model. J. Theor. Biol. 2005, 235, 373–380. [Google Scholar] [CrossRef]
- Ballantyne, F.; Kerkhoff, A.J. The observed range for temporal mean-variance scaling exponents can be explained by reproductive correlation. Oikos 2007, 116, 174–180. [Google Scholar] [CrossRef]
- Morand, S.; Guegan, J.F. Distribution and abundance of parasite nematodes: Ecological specialisation, phylogenetic constraint or simply epidemiology? Oikos 2000, 88, 563–573. [Google Scholar] [CrossRef]
- Charlier, J.; Bartley, D.J.; Sotiraki, S.; Martinez-Valladares, M.; Claerebout, E.; Von Samson-Himmelstjerna, G.; Thamsborg, S.M.; Hoste, H.; Morgan, E.R.; Rinaldi, L. Anthelmintic resistance in ruminants: Challenges and solutions. Adv. Parasitol. 2022, 115, 171–227. [Google Scholar]
- Coles, G.C. Sustainable use of anthelmintics in grazing animals. Vet. Rec. 2002, 151, 165–169. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Factor | Host | Climate | Age | Sex | Method | Parasite | Treatment | Tracer | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Categories (n) | 5 | 3 | 3 | 3 | 2 | 5 | 5 | 3 | |||||||
| Sheep | 129 | Temp | 202 | Juv | 180 | M | 42 | PM | 234 | Haemonchus | 98 | N/yr | Perm | 289 | |
| Goat | 81 | Trop | 36 | Ad | 128 | F | 74 | FEC | 91 | T. axei | 91 | Trac | 33 | ||
| Cattle | 22 | Arid | 87 | Mix | 17 | Mix | 209 | Marshallagia | 25 | Expmtl | 3 | ||||
| Deer | 66 | Other trichostrongylids | 33 | ||||||||||||
| Saiga | 27 | Trichuris | 51 |
| Taxon | Intercept, a | 95% CI | Slope, b | 95% CI |
|---|---|---|---|---|
| Haemonchus | −0.515 | −0.955–−0.075 | 2.057 | 1.884–2.229 |
| T. axei | 0.172 | −0.056–0.400 | 1.956 | 1.833–2.079 |
| Marshallagia | 1.077 | 0.796–1.358 | 1.937 | 1.736–2.137 |
| Trichuris | 0.987 | 0.791–1.184 | 1.886 | 1.763–2.010 |
| Nematodirus | 1.130 | 0.887–1.373 | 1.847 | 1.699–1.995 |
| Other trichostrongyles | 1.058 | 0.759–1.357 | 1.750 | 1.508–1.993 |
| Treatment Frequency | 0 | 1 | 2 | 3 | 4 |
|---|---|---|---|---|---|
| Host species | Deer | Sheep | Goats | Goats | Goats, cattle |
| a (intercept) | 1.305 | 0.215 | 0.713 | 0.375 | 0.529 |
| b (slope) | 1.517 | 1.970 | 1.209 | 2.029 | 1.889 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morgan, E.R.; Segonds-Pichon, A.; Ferté, H.; Duncan, P.; Cabaret, J. Anthelmintic Treatment and the Stability of Parasite Distribution in Ruminants. Animals 2023, 13, 1882. https://doi.org/10.3390/ani13111882
Morgan ER, Segonds-Pichon A, Ferté H, Duncan P, Cabaret J. Anthelmintic Treatment and the Stability of Parasite Distribution in Ruminants. Animals. 2023; 13(11):1882. https://doi.org/10.3390/ani13111882
Chicago/Turabian StyleMorgan, Eric R., Anne Segonds-Pichon, Hubert Ferté, Patrick Duncan, and Jacques Cabaret. 2023. "Anthelmintic Treatment and the Stability of Parasite Distribution in Ruminants" Animals 13, no. 11: 1882. https://doi.org/10.3390/ani13111882