
Managing Reproduction in Hyperprolific Sow Herds
 ,
,  , and
, and 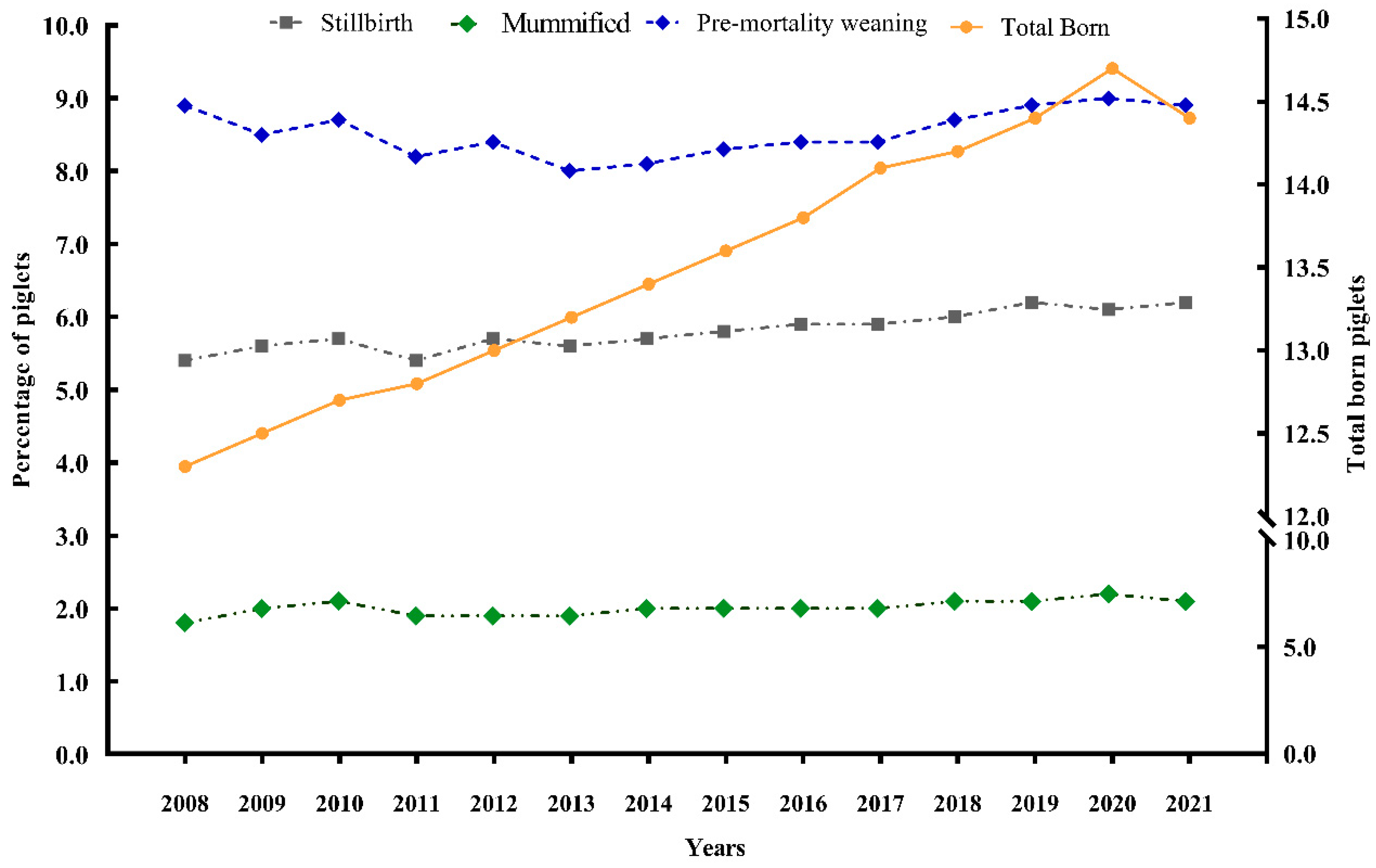{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Birth Traits, Growth, and Body Development of Replacement Gilt
3. Controlling the Gestation Phase to Avoid Reproductive Failures
4. Farrowing Traits of Hyperprolific Sows and Their Consequences
5. Neonatal Losses: Incidence of Stillbirths and Low Viability Piglets
6. Colostrum Yield
7. Female Health, Reproductive Performance, and Longevity
8. Strategies and Tools to Optimize Performance at Farrowing
9. Managing Lactating and Weaned Sows
10. Management of Hyperprolific Litters in the Lactational Period
10.1. Number of Piglets and Functional Teats
10.2. Cross-Fostering Protocols
10.3. Pre-Weaning Litter Socialization
10.4. Nurse Sows and Supplementary Feeding
10.5. Increased Weaning Age
11. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tokach, M.D.; Menegat, M.B.; Gourley, K.M.; Goodband, R.D. Review: Nutrient Requirements of the Modern High-Producing Lactating Sow, with an Emphasis on Amino Acid Requirements. Animal 2019, 13, 2967–2977. [Google Scholar] [CrossRef] [PubMed]
- Danish Pig Research Centre. Combination of Key Figures of Production Performance in Danish Swine Herds between 2000 and 2020 [in Danish: Landsgennemsnit for Produktitivet i Svineproduktionen Fra 2000 Til 2020]; SEGES, Danish Pig Research Centre (DPRC): Copenhagen, Denmark, 2020. [Google Scholar]
- Bortolozzo, F.P.; Bernardi, M.L.; Kummer, R.; Wentz, I. Growth, Body State and Breeding Performance in Gilts and Primiparous Sows. Soc. Reprod. Fertil. Suppl. 2009, 66, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Magnabosco, D.; Cunha, E.C.P.; Bernardi, M.L.; Wentz, I.; Bortolozzo, F.P. Impact of the Birth Weight of Landrace x Large White Dam Line Gilts on Mortality, Culling and Growth Performance until Selection for Breeding Herd. Acta Sci. Vet. 2015, 43, 1274. [Google Scholar]
- Da Silva, C.L.A.; van den Brand, H.; Laurenssen, B.F.A.; Broekhuijse, M.L.W.J.; Knol, E.F.; Kemp, B.; Soede, N.M. Relationships between Ovulation Rate and Embryonic and Placental Characteristics in Multiparous Sows at 35 Days of Pregnancy. Animal 2016, 10, 1192–1199. [Google Scholar] [CrossRef] [PubMed]
- Magnabosco, D.; Bernardi, M.L.; Wentz, I.; Cunha, E.C.P.; Bortolozzo, F.P. Low Birth Weight Affects Lifetime Productive Performance and Longevity of Female Swine. Livest. Sci. 2016, 184, 119–125. [Google Scholar] [CrossRef]
- Patterson, J.; Bernardi, M.L.; Allerson, M.; Hanson, A.; Holden, N.; Bruner, L.; Pinilla, J.C.; Foxcroft, G. Associations among Individual Gilt Birth Weight, Litter Birth Weight Phenotype, and the Efficiency of Replacement Gilt Production. J. Anim. Sci. 2020, 98, skaa331. [Google Scholar] [CrossRef]
- Moroni, J.L.; Tsoi, S.; Wenger, I.I.; Tran, C.; Plastow, G.S.; Charagu, P.; Dyck, M.K. The Influence of Litter Birth Weight Phenotype on Embryonic and Placental Development at Day 30 of Gestation in Multiparous Purebred Large White Sows. Anim. Reprod. Sci. 2022, 244, 107035. [Google Scholar] [CrossRef]
- Patterson, J.; Foxcroft, G. Gilt Management for Fertility and Longevity. Animals 2019, 9, 434. [Google Scholar] [CrossRef]
- Patterson, J.; Foxcroft, G.; Holden, N.; Allerson, M.; Hanson, A.; Triemert, E.; Bruner, L.; Pinilla, J.C. 117 A Low Litter Birth Weight Phenotype Reduces the Retention Rate of Potential Replacement Gilts. J. Anim. Sci. 2018, 96, 62. [Google Scholar] [CrossRef]
- Smit, M.N.; Spencer, J.D.; Almeida, F.R.C.L.; Patterson, J.L.; Chiarini-Garcia, H.; Dyck, M.K.; Foxcroft, G.R. Consequences of a Low Litter Birth Weight Phenotype for Postnatal Lean Growth Performance and Neonatal Testicular Morphology in the Pig. Animal 2013, 7, 1681–1689. [Google Scholar] [CrossRef]
- Tummaruk, P. Effects of Season, Outdoor Climate and Photo Period on Age at First Observed Estrus in Landrace × Yorkshire Crossbred Gilts in Thailand. Livest. Sci. 2012, 144, 163–172. [Google Scholar] [CrossRef]
- Knox, R.V.; Arend, L.S.; Buerkley, A.L.; Patterson, J.L.; Foxcroft, G.R. Effects of Physical or Fenceline Boar Exposure and Exogenous Gonadotropins on Puberty Induction and Subsequent Fertility in Gilts. J. Anim. Sci. 2021, 99, skab348. [Google Scholar] [CrossRef] [PubMed]
- Newton, E.A.; Mahan, D.C. Effect of Feed Intake during Late Development on Pubertal Onset and Resulting Body Composition in Crossbred Gilts. J. Anim. Sci. 1992, 70, 3774–3780. [Google Scholar] [CrossRef] [PubMed]
- Prunier, A.; Bonneau, M.; Etienne, M. Effects of Age and Live Weight on the Sexual Development of Gilts and Boars Fed Two Planes of Nutrition. Reprod. Nutr. Dev. 1987, 27, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Beltranena, E.; Aherne, F.X.; Foxcroft, G.R. Innate Variability in Sexual Development Irrespective of Body Fatness in Gilts. J. Anim. Sci. 1993, 71, 471–480. [Google Scholar] [CrossRef]
- Gaughan, J.B.; Cameron, R.D.A.; Dryden, G.M.L.; Young, B.A. Effect of Body Composition at Selection on Reproductive Development in Large White Gilts. J. Anim. Sci. 1997, 75, 1764–1772. [Google Scholar] [CrossRef]
- Sankarganesh, D.; Kirkwood, R.; Angayarkanni, J.; Achiraman, S.; Archunan, G. Pig Pheromones and Behaviors: A Review. Theriogenology 2021, 175, 1–6. [Google Scholar] [CrossRef]
- Beltranena, E.; Aherne, F.X.; Foxcroft, G.R.; Kirkwood, R.N. Effects of Pre- and Postpubertal Feeding on Production Traits at First and Second Estrus in Gilts. J. Anim. Sci. 1991, 69, 886–893. [Google Scholar] [CrossRef]
- Foxcroft, G.; Beltranena, E.; Patterson, J.; Williams, N. The Biological Basis for Implementing Effective Replacement Gilt Management. In Proceedings of the Allen D. Leman Swine Pre-Conference Reproduction Workshop, Saint Paul, MN, USA, 17–20 September 2005; pp. 5–25. [Google Scholar]
- Filha, W.S.A.; Bernardi, M.L.; Wentz, I.; Bortolozzo, F.P. Reproductive Performance of Gilts According to Growth Rate and Backfat Thickness at Mating. Anim. Reprod. Sci. 2010, 121, 139–144. [Google Scholar] [CrossRef]
- Kummer, R.; Bernardi, M.L.; Schenkel, A.C.; Amaral Filha, W.S.; Wentz, I.; Bortolozzo, F.P. Reproductive Performance of Gilts with Similar Age but with Different Growth Rate at the Onset of Puberty Stimulation. Reprod. Domest. Anim. 2009, 44, 255–259. [Google Scholar] [CrossRef]
- Kirkwood, R.N.; Thacker, P.A. Management of Replacement Breeding Animals. Vet. Clin. N. Am. Food Anim. Pract. 1992, 8, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.; Patterson, J.; Foxcroft, G.; Pettitt, M. Non-Negotiable Aspects of Gilt Development. Adv. Pork Prod. 2005, 16, 281–289. [Google Scholar]
- Theil, P.K.; Krogh, U.; Bruun, T.S.; Feyera, T. Feeding the Modern Sow to Sustain High Productivity. Mol. Reprod. Dev. 2022. [Google Scholar] [CrossRef]
- Kummer, R.; Bernardi, M.L.; Wentz, I.; Bortolozzo, F.P. Reproductive Performance of High Growth Rate Gilts Inseminated at an Early Age. Anim. Reprod. Sci. 2006, 96, 47–53. [Google Scholar] [CrossRef]
- Clowes, E.J.; Aherne, F.X.; Schaefer, A.L.; Foxcroft, G.R.; Baracos, V.E. Parturition Body Size and Body Protein Loss during Lactation Influence Performance during Lactation and Ovarian Function at Weaning in First-Parity Sows. J. Anim. Sci. 2003, 81, 1517–1528. [Google Scholar] [CrossRef]
- Mallmann, A.L.; Oliveira, G.S.; Ulguim, R.R.; Mellagi, A.P.G.; Bernardi, M.L.; Orlando, U.A.D.; Goncalves, M.A.D.; Cogo, R.J.; Bortolozzo, F.P. Impact of Feed Intake in Early Gestation on Maternal Growth and Litter Size According to Body Reserves at Weaning of Young Parity Sows. J. Anim. Sci. 2020, 98, skaa075. [Google Scholar] [CrossRef] [PubMed]
- Lents, C.A.; Supakorn, C.; DeDecker, A.E.; Phillips, C.E.; Boyd, R.D.; Vallet, J.L.; Rohrer, G.A.; Foxcroft, G.R.; Flowers, W.L.; Trottier, N.L.; et al. Dietary Lysine-to-Energy Ratios for Managing Growth and Pubertal Development in Replacement Gilts. Appl. Anim. Sci. 2020, 36, 701–714. [Google Scholar] [CrossRef]
- Johnson, R.K.; Trenhaile-Grannemann, M.D.; Moreno, R.; Ciobanu, D.C.; Miller, P.S. Effects of Restricting Energy during the Gilt Development Period on Growth and Reproduction of Lines Differing in Lean Growth Rate: Responses in Reproductive Performance and Longevity. J. Anim. Sci. 2022, 100, skab352. [Google Scholar] [CrossRef]
- Bruun, T.S.; Bache, J.K.; Amdi, C. The Effects of Long- or Short-Term Increased Feed Allowance Prior to First Service on Litter Size in Gilts. Transl. Anim. Sci. 2021, 5, txab005. [Google Scholar] [CrossRef]
- Lucia, T.; Dial, G.D.; Marsh, W.E. Lifetime Reproductive Performance in Female Pigs Having Distinct Reasons for Removal. Livest. Prod. Sci. 2000, 63, 213–222. [Google Scholar] [CrossRef]
- De Hollander, C.A.; Knol, E.F.; Heuven, H.C.M.; Van Grevenhof, E.M. Interval from Last Insemination to Culling: II. Culling Reasons from Practise and the Correlation with Longevity. Livest. Sci. 2015, 181, 25–30. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, X.; Mo, D.; Chen, Q.; Chen, Y. Analysis of Reasons for Sow Culling and Seasonal Effects on Reproductive Disorders in Southern China. Anim. Reprod. Sci. 2015, 159, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Mellagi, A.P.G.; Will, K.J.; Quirino, M.; Bustamante-Filho, I.C.; Ulguim, R.D.R.; Bortolozzo, F.P. Update on Artificial Insemination: Semen, Techniques, and Sow Fertility. Mol. Reprod. Dev. 2022, 1–11. [Google Scholar] [CrossRef]
- Galli, M.C.; Boyle, L.A.; Mazzoni, C.; Contiero, B.; Stefani, A.; Bertazzo, V.; Mereghetti, F.; Gottardo, F. Can We Further Reduce the Time Pregnant Sows Spend in Gestation Stalls? Livest. Sci. 2022, 264, 105049. [Google Scholar] [CrossRef]
- Perry, J.; Heap, R.; Burton, R.; Gadsby, J. Endocrinology of Blastocyst and Its Role in the Establishment of Pregnancy. J. Reprod. Fertil. Suppl. 1976, 25, 85–104. [Google Scholar]
- Knox, R.; Salak-Johnson, J.; Hopgood, M.; Greiner, L.; Connor, J. Effect of Day of Mixing Gestating Sows on Measures of Reproductive Performance and Animal Welfare. J. Anim. Sci. 2014, 92, 1698–1707. [Google Scholar] [CrossRef] [PubMed]
- Cunha, E.C.P.; de Alcantara Menezes, T.; Bernardi, M.L.; Mellagi, A.P.G.; da Rosa Ulguim, R.; Wentz, I.; Bortolozzo, F.P. Reproductive Performance, Offspring Characteristics, and Injury Scores According to the Housing System of Gestating Gilts. Livest. Sci. 2018, 210, 59–67. [Google Scholar] [CrossRef]
- Bampi, D.; Borstnez, K.K.; Dias, C.P.; Costa, O.A.D.; Moreira, F.; Peripolli, V.; Oliveira Júnior, J.M.; Schwegler, E.; Rauber, L.P.; Bianchi, I. Evaluation of Reproductive and Animal Welfare Parameters of Swine Females of Different Genetic Lines Submitted to Different Reproductive Management and Housing Systems during Pregnancy. Arq. Bras. Med. Vet. Zootec. 2020, 72, 1675–1682. [Google Scholar] [CrossRef]
- Stevens, B.; Karlen, G.M.; Morrison, R.; Gonyou, H.W.; Butler, K.L.; Kerswell, K.J.; Hemsworth, P.H. Effects of Stage of Gestation at Mixing on Aggression, Injuries and Stress in Sows. Appl. Anim. Behav. Sci. 2015, 165, 40–46. [Google Scholar] [CrossRef]
- Magoga, J.; Vier, C.E.; Mallmann, A.L.; Mellagi, A.P.G.; Cogo, R.J.; Bortolozzo, F.P.; da Rosa Ulguim, R. Reproductive Performance of Gilts and Weaned Sows Grouped at Different Days after Insemination. Trop. Anim. Health Prod. 2023, 55, 31. [Google Scholar] [CrossRef]
- Leal, D.F.; Muro, B.B.D.; Nichi, M.; Almond, G.W.; Viana, C.H.C.; Vioti, G.; Carnevale, R.F.; Garbossa, C.A.P. Effects of Post-Insemination Energy Content of Feed on Embryonic Survival in Pigs: A Systematic Review. Anim. Reprod. Sci. 2019, 205, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Langendijk, P. Latest Advances in Sow Nutrition during Early Gestation. Animals 2021, 11, 1720. [Google Scholar] [CrossRef] [PubMed]
- Salak-Johnson, J.L. Social Status and Housing Factors Affect Reproductive Performance of Pregnant Sows in Groups. Mol. Reprod. Dev. 2017, 84, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Mallmann, A.L.; Betiolo, F.B.; Camilloti, E.; Mellagi, A.P.G.; Ulguim, R.R.; Wentz, I.; Bernardi, M.L.; Gonçalves, M.A.D.; Kummer, R.; Bortolozzo, F.P. Two Different Feeding Levels during Late Gestation in Gilts and Sows under Commercial Conditions: Impact on Piglet Birth Weight and Female Reproductive Performance. J. Anim. Sci. 2018, 96, 4209–4219. [Google Scholar] [CrossRef]
- Gonçalves, M.A.D.; Dritz, S.S.; Tokach, M.D.; Piva, J.H.; de Rouchey, J.M.; Woodworth, J.C.; Goodband, R.D. Fact Sheet—Impact of Increased Feed Intake during Late Gestation on Reproductive Performance of Gilts and Sows. J. Swine Health Prod. 2016, 24, 264–266. [Google Scholar]
- De Oliveira Araújo, V.; de Oliveira, R.A.; Vieira, M.D.F.A.; Silveira, H.; da Silva Fonseca, L.; Alves, L.K.S.; Guimarães, E.B.B.; Schinckel, A.P.; Garbossa, C.A.P. Bump Feed for Gestating Sows Is Really Necessary? Livest. Sci. 2020, 240, 104184. [Google Scholar] [CrossRef]
- Moreira, R.H.R.; Perez Palencia, J.Y.; Moita, V.H.C.; Caputo, L.S.S.; Saraiva, A.; Andretta, I.; Ferreira, R.A.; de Abreu, M.L.T. Variability of Piglet Birth Weights: A Systematic Review and Meta-analysis. J. Anim. Physiol. Anim. Nutr. 2020, 104, 657–666. [Google Scholar] [CrossRef]
- Iida, R.; Piñeiro, C.; Koketsu, Y. Removal of Sows in Spanish Breeding Herds Due to Lameness: Incidence, Related Factors and Reproductive Performance of Removed Sows. Prev. Vet. Med. 2020, 179, 105002. [Google Scholar] [CrossRef]
- Kikuti, M.; Preis, G.M.; Deen, J.; Pinilla, J.C.; Corzo, C.A. Sow Mortality in a Pig Production System in the Midwestern USA: Reasons for Removal and Factors Associated with Increased Mortality. Vet. Rec. 2022, 192, e2539. [Google Scholar] [CrossRef]
- Schwertz, C.I.; Bianchi, R.M.; Cecco, B.S.; Pavarini, S.P.; Driemeier, D. Causes of Death of Sows in Three Brazilian Pig Farms. Pesqui. Vet. Bras. 2021, 41, e06857. [Google Scholar] [CrossRef]
- Iida, R.; Piñeiro, C.; Koketsu, Y. Incidences and Risk Factors for Prolapse Removal in Spanish Sow Herds. Prev. Vet. Med. 2019, 163, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Chipman, A.; Rademacher, C.; Johnson, C.; Stalder, K.; Johnson, A.; Keating, A.; Patience, J.; Gabler, N.; Linhares, D.; Schwartz, K. Pelvic Organ Prolapse: An Industry-Wide Collaboration to Identify Putative Contributing Factors; Iowa State University: Ames, IA, USA, 2018. [Google Scholar]
- Pierozan, C.R.; Callegari, M.A.; Dias, C.P.; de Souza, K.L.; Gasa, J.; da Silva, C.A. Herd-Level Factors Associated with Non-Productive Days and Farrowing Rate in Commercial Pig Farms in Two Consecutive Years. Livest. Sci. 2021, 244, 104312. [Google Scholar] [CrossRef]
- Agriness: Relatório Anual Do Desempenho da Produção de Suínos. 2022. Available online: https://melhores.agriness.com/ (accessed on 10 January 2023).
- Baxter, E.M.; Schmitt, O.; Pedersen, L.J. Managing the Litter from Hyperprolific Sows. In The Suckling and Weaned Piglet; Wageningen Academic Publishers: Wageningen, The Netherlands, 2020; pp. 71–106. [Google Scholar] [CrossRef]
- Van Dijk, A.J.; Van Rens, B.T.T.M.; Van Der Lende, T.; Taverne, M.A.M. Factors Affecting Duration of the Expulsive Stage of Parturition and Piglet Birth Intervals in Sows with Uncomplicated, Spontaneous Farrowings. Theriogenology 2005, 64, 1573–1590. [Google Scholar] [CrossRef] [PubMed]
- Björkman, S.; Oliviero, C.; Kauffold, J.; Soede, N.M.; Peltoniemi, O.A.T. Prolonged Parturition and Impaired Placenta Expulsion Increase the Risk of Postpartum Metritis and Delay Uterine Involution in Sows. Theriogenology 2018, 106, 87–92. [Google Scholar] [CrossRef]
- Oliviero, C.; Heinonen, M.; Valros, A.; Peltoniemi, O. Environmental and Sow-Related Factors Affecting the Duration of Farrowing. Anim. Reprod. Sci. 2010, 119, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Martínez-Burnes, J.; Trujillo-Ortega, M.E.; Alonso-Spilsbury, M.L.; Ramírez-Necoechea, R.; López, A. Effect of Oxytocin Treatment in Sows on Umbilical Cord Morphology, Meconium Staining, and Neonatal Mortality of Piglets. Am. J. Vet. Res. 2002, 63, 1571–1574. [Google Scholar] [CrossRef]
- Martínez-Burnes, J.; Muns, R.; Barrios-García, H.; Villanueva-García, D.; Domínguez-Oliva, A.; Mota-Rojas, D. Parturition in Mammals: Animal Models, Pain and Distress. Animals 2021, 11, 2960. [Google Scholar] [CrossRef] [PubMed]
- Tummaruk, P.; Sang-Gassanee, K. Effect of Farrowing Duration, Parity Number and the Type of Anti-Inflammatory Drug on Postparturient Disorders in Sows: A Clinical Study. Trop. Anim. Health Prod. 2013, 45, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Peltoniemi, O.; Oliviero, C.; Yun, J.; Grahofer, A.; Björkman, S. Management Practices to Optimize the Parturition Process in the Hyperprolific Sow. J. Anim. Sci. 2020, 98, S96. [Google Scholar] [CrossRef]
- Oliviero, C.; Kothe, S.; Heinonen, M.; Valros, A.; Peltoniemi, O. Prolonged Duration of Farrowing Is Associated with Subsequent Decreased Fertility in Sows. Theriogenology 2013, 79, 1095–1099. [Google Scholar] [CrossRef] [PubMed]
- Peltoniemi, O.A.T.; Oliviero, C. Housing, Management and Environment during Farrowing and Early Lactation. In The Gestating and Lactating Sow; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015; pp. 231–252. [Google Scholar] [CrossRef]
- Borges, V.F.; Bernardi, M.L.; Bortolozzo, F.P.; Wentz, I. Risk Factors for Stillbirth and Foetal Mummification in Four Brazilian Swine Herds. Prev. Vet. Med. 2005, 70, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Herpin, P.; Le Dividich, J.; Hulin, J.C.; Fillaut, M.; De Marco, F.; Bertin, R. Effects of the Level of Asphyxia during Delivery on Viability at Birth and Early Postnatal Vitality of Newborn Pigs. J. Anim. Sci. 1996, 74, 2067–2075. [Google Scholar] [CrossRef] [PubMed]
- Panzardi, A.; Bernardi, M.L.; Mellagi, A.P.; Bierhals, T.; Bortolozzo, F.P.; Wentz, I. Newborn Piglet Traits Associated with Survival and Growth Performance until Weaning. Prev. Vet. Med. 2013, 110, 206–213. [Google Scholar] [CrossRef]
- Baxter, E.M.; Jarvis, S.; D’Eath, R.B.; Ross, D.W.; Robson, S.K.; Farish, M.; Nevison, I.M.; Lawrence, A.B.; Edwards, S.A. Investigating the Behavioural and Physiological Indicators of Neonatal Survival in Pigs. Theriogenology 2008, 69, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Ash, M. Management of the Farrowing and Lactating Sow. In Current Therapy in Theriogenology, 2nd ed.; WB Saunders Company: Philadelphia, PA, USA, 1986; pp. 931–934. [Google Scholar]
- Declerck, I.; Sarrazin, S.; Dewulf, J.; Maes, D. Sow and Piglet Factors Determining Variation of Colostrum Intake between and within Litters. Animal 2017, 11, 1336–1343. [Google Scholar] [CrossRef] [PubMed]
- Quesnel, H.; Farmer, C.; Devillers, N. Colostrum Intake: Influence on Piglet Performance and Factors of Variation. Livest. Sci. 2012, 146, 105–114. [Google Scholar] [CrossRef]
- Declerck, I.; Dewulf, J.; Piepers, S.; Decaluwé, R.; Maes, D. Sow and Litter Factors Influencing Colostrum Yield and Nutritional Composition. J. Anim. Sci. 2015, 93, 1309–1317. [Google Scholar] [CrossRef]
- Quesnel, H. Colostrum Production by Sows: Variability of Colostrum Yield and Immunoglobulin G Concentrations. Animal 2011, 5, 1546–1553. [Google Scholar] [CrossRef]
- Machado, A.P.; Otto, M.A.; Bernardi, M.L.; Wentz, I.; Bortolozzo, F.P. Factors Influencing Colostrum Yield by Sows. Arq. Bras. Med. Vet. Zootec. 2016, 68, 553–561. [Google Scholar] [CrossRef]
- Friendship, R.M.; O’Sullivan, T.L. Sow Health. In The Gestating and Lactating Sow; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015; pp. 409–422. ISBN 9789086868032. [Google Scholar]
- Klopfenstein, C.; Farmer, C.; Martineau, G. Diseases of the Ammary Glands and Lactation Problems Diseases of Swine; Iowa State University Press: Ames, IA, USA, 2000; p. 833. [Google Scholar]
- Pearodwong, P.; Muns, R.; Tummaruk, P. Prevalence of Constipation and Its Influence on Post-Parturient Disorders in Tropical Sows. Trop. Anim. Health Prod. 2016, 48, 525–531. [Google Scholar] [CrossRef]
- Einarsson, S. Agalactia in Sows. In Current Therapy in Theriogenology; Morrow, D.A., Ed.; Saunders: Philadelphia, PA, USA, 1986; Volume 2, pp. 935–937. [Google Scholar]
- Egli, P.T.; Schüpbach-Regula, G.; Nathues, H.; Ulbrich, S.E.; Grahofer, A. Influence of the Farrowing Process and Different Sow and Piglet Traits on Uterine Involution in a Free Farrowing System. Theriogenology 2022, 182, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mellagi, A.P.G.; Heim, G.; Bernardi, M.L.; Bortolozzo, F.P.; Wentz, I. Caracterização e Desempenho Reprodutivo de Fêmeas Suínas Submetidas à Intervenção Obstétrica Manual. Ciência Rural. 2009, 39, 1478–1484. [Google Scholar] [CrossRef]
- Iida, R.; Koketsu, Y. Climatic Factors Associated with Peripartum Pig Deaths during Hot and Humid or Cold Seasons. Prev. Vet. Med. 2014, 115, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, M.S.; Matias, D.N.; Poor, A.P.; Dutra, M.C.; Moreno, L.Z.; Parra, B.M.; Silva, A.P.S.; Matajira, C.E.C.; de Moura Gomes, V.T.; Barbosa, M.R.F.; et al. Causes of Sow Mortality and Risks to Post-Mortem Findings in a Brazilian Intensive Swine Production System. Animals 2022, 12, 1804. [Google Scholar] [CrossRef]
- Vearick, G.; Mellagi, A.P.G.; Bortolozzo, F.P.; Wentz, I.; Bernardi, M.L. Causes of Mortality in Swine Female. Arch. Vet. Sci. 2008, 13, 126–132. [Google Scholar] [CrossRef]
- Supakorn, C.; Christianson, M.I.; Howard, J.; Gray, K.A.; Stalder, K.J. Heritability Estimates for Sow Prolapse. Livest. Sci. 2019, 227, 111–113. [Google Scholar] [CrossRef]
- Ross, J.W. Identification of Putative Factors Contributing to Pelvic Organ Prolapse in Sows (Grant# 17-224) II. Industry Summary 2019. Available online: https://porkcheckoff.org/research/identification-putative-factors-contributing-pelvic-organ-prolapse-sows/ (accessed on 10 January 2023).
- Supakorn, C.; Stock, J.; Stalder, K.; Hostetler, C. Prolapse Incidence in Swine Breeding Herds Is a Cause for Concern. Open J. Vet. Med. 2017, 7, 85–97. [Google Scholar]
- Kiefer, Z.E.; Koester, L.R.; Showman, L.; Studer, J.M.; Chipman, A.L.; Keating, A.F.; Schmitz-Esser, S.; Ross, J.W. Vaginal Microbiome and Serum Metabolite Differences in Late Gestation Commercial Sows at Risk for Pelvic Organ Prolapse. Sci. Rep. 2021, 11, 6189. [Google Scholar] [CrossRef]
- Kiefer, Z.E.; Studer, J.M.; Chipman, A.L.; Adur, M.K.; Mainquist-Whigham, C.; Gabler, N.K.; Keating, A.F.; Ross, J.W. Circulating Biomarkers Associated with Pelvic Organ Prolapse Risk in Late Gestation Sows. J. Anim. Sci. 2021, 99, skab207. [Google Scholar] [CrossRef]
- Jiarpinitnun, P.; Loyawatananan, S.; Sangratkanjanasin, P.; Kompong, K.; Nuntapaitoon, M.; Muns, R.; De Rensis, F.; Tummaruk, P. Administration of Carbetocin after the First Piglet Was Born Reduced Farrowing Duration but Compromised Colostrum Intake in Newborn Piglets. Theriogenology 2019, 128, 23–30. [Google Scholar] [CrossRef]
- Hill, S.V.; del Rocio Amezcua, M.; Ribeiro, E.S.; O’Sullivan, T.L.; Friendship, R.M. Defining the Effect of Oxytocin Use in Farrowing Sows on Stillbirth Rate: A Systematic Review with a Meta-Analysis. Animals 2022, 12, 1795. [Google Scholar] [CrossRef] [PubMed]
- Vongsariyavanich, S.; Sundaraketu, P.; Sakulsirajit, R.; Suriyapornchaikul, C.; Therarachatamongkol, S.; Boonraungrod, N.; Pearodwong, P.; Tummaruk, P. Effect of Carbetocin Administration during the Mid-Period of Parturition on Farrowing Duration, Newborn Piglet Characteristics, Colostrum Yield and Milk Yield in Hyperprolific Sows. Theriogenology 2021, 172, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Kamphues, J.; Tabeling, R.; Schwier, S. Die Kotqualität von Sauen Unter Dem Einfluss Verschiedener Fütterungs-Und Haltungsbedingungen [Feces Quality of Sows under the Influence of Different Feeding and Housing Conditions]. Dtsch. Tierarztl. Wochenschr. 2020, 107, 380. [Google Scholar]
- Theil, P.K. Transition Feeding of Sows. In The Gestating and Lactating Sow; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015; pp. 147–172. ISBN 9789086868032. [Google Scholar]
- Oliviero, C.; Kokkonen, T.; Heinonen, M.; Sankari, S.; Peltoniemi, O. Feeding Sows with High Fibre Diet around Farrowing and Early Lactation: Impact on Intestinal Activity, Energy Balance Related Parameters and Litter Performance. Res. Vet. Sci. 2009, 86, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Guillemet, R.; Hamard, A.; Quesnel, H.; Père, M.C.; Etienne, M.; Dourmad, J.Y.; Meunier-Salaün, M.C. Dietary Fibre for Gestating Sows: Effects on Parturition Progress, Behaviour, Litter and Sow Performance. Animal 2007, 1, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, Y.; Feng, B.; Xuan, Y.; Che, L.; Fang, Z.; Lin, Y.; Xu, S.; Li, J.; Feng, B.; Wu, D. Inclusion of Purified Dietary Fiber during Gestation Improved the Reproductive Performance of Sows. J. Anim. Sci. Biotechnol. 2020, 11, 47. [Google Scholar] [CrossRef]
- Feyera, T.; Højgaard, C.K.; Vinther, J.; Bruun, T.S.; Theil, P.K. Dietary Supplement Rich in Fiber Fed to Late Gestating Sows during Transition Reduces Rate of Stillborn Piglets. J. Anim. Sci. 2017, 95, 5430–5438. [Google Scholar] [CrossRef]
- Wu, J.; Xiong, Y.; Zhong, M.; Li, Y.; Wan, H.; Wu, D.; Liu, Q. Effects of Purified Fibre-Mixture Supplementation of Gestation Diet on Gut Microbiota, Immunity and Reproductive Performance of Sows. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1144–1154. [Google Scholar] [CrossRef]
- Omari, M.; Plöntzke, J.; Röblitz, S. A Pharmacokinetic-Pharmacodynamic Model for Single Dose Administration of Dexamethasone in Dairy Cows; ZIB Report 19-53; Zuse Institute Berlin: Berlin, Germany, 2019. [Google Scholar]
- Will, K.J.; Magoga, J.; De Conti, E.R.; da Rosa Ulguim, R.; Mellagi, A.P.G.; Bortolozzo, F.P. Relationship between Dexamethasone Treatment around Parturition of Primiparous Sows and Farrowing Performance and Newborn Piglet Traits. Theriogenology 2023, 198, 256–263. [Google Scholar] [CrossRef]
- Ward, S.A.; Kirkwood, R.N.; Plush, K.J. Administering Dexamethasone to Prepartum Sows: Effects on Sow and Piglet Performance. Livest. Sci. 2020, 239, 104171. [Google Scholar] [CrossRef]
- Farmer, C.; Edwards, S.A. Review: Improving the Performance of Neonatal Piglets. Animal 2022, 16, 100350. [Google Scholar] [CrossRef] [PubMed]
- Eissen, J.J.; Apeldoorn, E.J.; Kanis, E.; Verstegen, M.W.A.; De Greef, K.H. The Importance of a High Feed Intake during Lactation of Primiparous Sows Nursing Large Litters. J. Anim. Sci. 2003, 81, 594–603. [Google Scholar] [CrossRef]
- Muro, B.B.D.; Carnevale, R.F.; Leal, D.F.; Almond, G.W.; Monteiro, M.S.; Poor, A.P.; Schinckel, A.P.; Garbossa, C.A.P. The Importance of Optimal Body Condition to Maximise Reproductive Health and Perinatal Outcomes in Pigs. Nutr. Res. Rev. 2022. [Google Scholar] [CrossRef]
- Decaluwé, R.; Maes, D.; Declerck, I.; Cools, A.; Wuyts, B.; De Smet, S.; Janssens, G.P.J. Changes in Back Fat Thickness during Late Gestation Predict Colostrum Yield in Sows. Animal 2013, 7, 1999–2007. [Google Scholar] [CrossRef] [PubMed]
- Kemp, B.; da Silva, C.L.A.; Soede, N.M. Recent Advances in Pig Reproduction: Focus on Impact of Genetic Selection for Female Fertility. Reprod. Domest. Anim. 2018, 53, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Zak, L.J.; Cosgrove, J.R.; Aherne, F.X.; Foxcroft, G.R. Pattern of Feed Intake and Associated Metabolic and Endocrine Changes Differentially Affect Postweaning Fertility in Primiparous Lactating Sows. J. Anim. Sci. 1997, 75, 208–216. [Google Scholar] [CrossRef]
- Arend, L.S.; Vinas, R.F.; Silva, G.S.; Lower, A.J.; Connor, J.F.; Knox, R. V Effects of Nursing a Large Litter and Ovarian Response to Gonadotropins at Weaning on Subsequent Fertility in First Parity Sows. J. Anim. Sci. 2023, 101, skac398. [Google Scholar] [CrossRef]
- Patterson, J.; Zimmerman, P.; Dyck, M.; Foxcroft, G. Effect of Skip-a-Heat Breeding on Subsequent Reproductive Performance in 1st Parity Sows. Adv. Pork Prod. 2006, 17. [Google Scholar]
- Vinsky, M.D.; Novak, S.; Dixon, W.T.; Dyck, M.K.; Foxcroft, G.R. Nutritional Restriction in Lactating Primiparous Sows Selectively Affects Female Embryo Survival and Overall Litter Development. Reprod. Fertil. Dev. 2006, 18, 347–355. [Google Scholar] [CrossRef]
- Schenkel, A.C.; Bernardi, M.L.; Bortolozzo, F.P.; Wentz, I. Body Reserve Mobilization during Lactation in First Parity Sows and Its Effect on Second Litter Size. Livest. Sci. 2010, 132, 165–172. [Google Scholar] [CrossRef]
- Soede, N.M.; Kemp, B. Best Practices in the Lactating and Weaned Sow to Optimize Reproductive Physiology and Performance. In The Gestating and Lactating Sow; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015; pp. 377–407. ISBN 9789086868032. [Google Scholar]
- Poleze, E.; Bernardi, M.L.; Amaral Filha, W.S.; Wentz, I.; Bortolozzo, F.P. Consequences of Variation in Weaning-to-Estrus Interval on Reproductive Performance of Swine Females. Livest. Sci. 2006, 103, 124–130. [Google Scholar] [CrossRef]
- Gianluppi, R.D.F.; Lucca, M.S.; Mellagi, A.P.G.; Bernardi, M.L.; Orlando, U.A.D.; Ulguim, R.R.; Bortolozzo, F.P. Effects of Different Amounts and Type of Diet during Weaning-to-Estrus Interval on Reproductive Performance of Primiparous and Multiparous Sows. Animal 2020, 14, 1906–1915. [Google Scholar] [CrossRef] [PubMed]
- Knauer, M.; Cassady, J. Effects of Preweaning Factors on Sow Lifetime Productivity-NPB#11-146; National Pork Board: Des Moines, IA, USA, 2016; pp. 11–14. [Google Scholar]
- Muns, R.; Nuntapaitoon, M.; Tummaruk, P. Non-Infectious Causes of Pre-Weaning Mortality in Piglets. Livest. Sci. 2016, 184, 46–57. [Google Scholar] [CrossRef]
- Alexopoulos, J.G.; Lines, D.S.; Hallett, S.; Plush, K.J. A Review of Success Factors for Piglet Fostering in Lactation. Animals 2018, 8, 38. [Google Scholar] [CrossRef]
- Le Dividich, J.; Martineau, G.P.; Madec, F.; Orgeur, P. Saving and Rearing Underprivileged and Supernumerary Piglets, and Improving Their Health at Weaning. In Weaning the Pig, Concepts and Consequences; Wageningen Pers.: Wageningen, The Netherlands, 2003; pp. 361–383. [Google Scholar]
- Balzani, A.; Cordell, H.J.; Edwards, S.A. Relationship of Sow Udder Morphology with Piglet Suckling Behavior and Teat Access. Theriogenology 2016, 86, 1913–1920. [Google Scholar] [CrossRef]
- Wiegert, J.G.; Knauer, M.T. 98 Sow Functional Teat Number Impacts Colostrum Intake and Piglet Throughput. J. Anim. Sci. 2018, 96, 51–52. [Google Scholar] [CrossRef]
- Obermier, D.; Eickhoff, M.; Mote, B.E.; Uitermarkt, A.; Frobose, H.; Borg, B. 57 The Impact of Functional Teat Number on Piglet Survival and Sow Efficiency. J. Anim. Sci. 2021, 99, 149. [Google Scholar] [CrossRef]
- Lundeheim, N.; Chalkias, H.; Rydhmer, L. Genetic Analysis of Teat Number and Litter Traits in Pigs. Acta Agric. Scand. A Anim. Sci. 2013, 63, 121–125. [Google Scholar] [CrossRef]
- Heim, G.; Mellagi, A.P.G.; Bierhals, T.; de Souza, L.P.; de Fries, H.C.C.; Piuco, P.; Seidel, E.; Bernardi, M.L.; Wentz, I.; Bortolozzo, F.P. Effects of Cross-Fostering within 24h after Birth on Pre-Weaning Behaviour, Growth Performance and Survival Rate of Biological and Adopted Piglets. Livest. Sci. 2012, 150, 121–127. [Google Scholar] [CrossRef]
- Souza, L.P.; Fries, H.C.C.; Heim, G.; Faccin, J.E.; Hernig, L.F.; Marimon, B.T.; Bernardi, M.L.; Bortolozzo, F.P.; Wentz, I. Behaviour and Growth Performance of Low-Birth-Weight Piglets Cross-Fostered in Multiparous Sows with Piglets of Higher Birth Weights. Arq. Bras. Med. Vet. Zootec. 2014, 66, 510–518. [Google Scholar] [CrossRef]
- Vande Pol, K.D.; Bautista, R.O.; Harper, H.; Shull, C.M.; Brown, C.B.; Ellis, M. Effect of Rearing Cross-Fostered Piglets in Litters of Either Uniform or Mixed Birth Weights on Preweaning Growth and Mortality. Transl. Anim. Sci. 2021, 5, txab030. [Google Scholar] [CrossRef] [PubMed]
- Bierhals, T.; Magnabosco, D.; Ribeiro, R.R.; Perin, J.; da Cruz, R.A.; Bernardi, M.L.; Wentz, I.; Bortolozzo, F.P. Influence of Pig Weight Classification at Cross-Fostering on the Performance of the Primiparous Sow and the Adopted Litter. Livest. Sci. 2012, 146, 115–122. [Google Scholar] [CrossRef]
- Earnhardt, A.L. The Genetics of Functional Teats in Swine. Master’s Thesis, North Carolina State University, Raleigh, NC, USA, 2019. [Google Scholar]
- Vande Pol, K.D.; Bautista, R.O.; Olivo, A.; Harper, H.; Shull, C.M.; Brown, C.B.; Ellis, M. Effect of Rearing Cross-Fostered Piglets in Litters of Differing Size Relative to Sow Functional Teat Number on Preweaning Growth and Mortality. Transl. Anim. Sci. 2021, 5, txab193. [Google Scholar] [CrossRef] [PubMed]
- Hessel, E.F.; Reiners, K.; Van Den Weghe, H.F.A. Socializing Piglets before Weaning: Effects on Behavior of Lactating Sows, Pre- and Postweaning Behavior, and Performance of Piglets. J. Anim. Sci. 2006, 84, 2847–2855. [Google Scholar] [CrossRef]
- North, L.; Stewart, A.H. The Effect of Mixing Litters Pre Weaning on the Performance of Piglets Pre and Post Weaning. Proc. Br. Soc. Anim. Sci. 2000, 2000, 135. [Google Scholar] [CrossRef]
- Salazar, L.C.; Ko, H.L.; Yang, C.H.; Llonch, L.; Manteca, X.; Camerlink, I.; Llonch, P. Early Socialisation as a Strategy to Increase Piglets’ Social Skills in Intensive Farming Conditions. Appl. Anim. Behav. Sci. 2018, 206, 25–31. [Google Scholar] [CrossRef]
- Camerlink, I.; Farish, M.; D’eath, R.B.; Arnott, G.; Turner, S.P. Long Term Benefits on Social Behaviour after Early Life Socialization of Piglets. Animals 2018, 8, 192. [Google Scholar] [CrossRef]
- Van Nieuwamerongen, S.E.; Soede, N.M.; van der Peet-Schwering, C.M.C.; Kemp, B.; Bolhuis, J.E. Development of Piglets Raised in a New Multi-Litter Housing System vs. Conventional Single-Litter Housing until 9 Weeks of Age. J. Anim. Sci. 2015, 93, 5442–5454. [Google Scholar] [CrossRef]
- Kobek-Kjeldager, C.; Moustsen, V.A.; Theil, P.K.; Pedersen, L.J. Managing Large Litters: Selected Measures of Performance in 10 Intermediate Nurse Sows and Welfare of Foster Piglets. Appl. Anim. Behav. Sci. 2020, 233, 105149. [Google Scholar] [CrossRef]
- Garrido-Mantilla, J.; Culhane, M.R.; Torremorell, M. Transmission of Influenza A Virus and Porcine Reproductive and Respiratory Syndrome Virus Using a Novel Nurse Sow Model: A Proof of Concept. Vet. Res. 2020, 51, 42. [Google Scholar] [CrossRef]
- Moreira, L.P.; Menegat, M.B.; Barros, G.P.; Bernardi, M.L.; Wentz, I.; Bortolozzo, F.P. Effects of Colostrum, and Protein and Energy Supplementation on Survival and Performance of Low-Birth-Weight Piglets. Livest. Sci. 2017, 202, 188–193. [Google Scholar] [CrossRef]
- Kummer, A.; Baroncello, E.; Moreira, L.; Bortolozzo, F.; Wentz, I. Efeitos Do Fornecimento Oral de Suplementos Nutricionais Na Sobrevivência e Crescimento de Leitões de Baixo Peso Ao Nascer. Acta Sci. Vet. 2015, 43, 1336. [Google Scholar]
- Viott, R.C.; Menezes, T.A.; Mellagi, A.P.G.; Bernardi, M.L.; Wentz, I.; Bortolozzo, F.P. Performance of Low Birth-Weight Piglets upon Protein-Energy and/or Colostrum Supplementation. Arq. Bras. Med. Vet. Zootec. 2018, 70, 1293–1300. [Google Scholar] [CrossRef]
- Kobek-Kjeldager, C.; Moustsen, V.A.; Theil, P.K.; Pedersen, L.J. Effect of Litter Size, Milk Replacer and Housing on Production Results of Hyper-Prolific Sows. Animal 2020, 14, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Pluske, J.R.; Hampson, D.J.; Williams, I.H. Factors Influencing the Structure and Function of the Small Intestine in the Weaned Pig: A Review. Livest. Prod. Sci. 1997, 51, 215–236. [Google Scholar] [CrossRef]
- Salak-Johnson, J.L.; Webb, S.R.; Salak-Johnson, J.L.; Webb, S.R. Short- and Long-Term Effects of Weaning Age on Pig Innate Immune Status. Open. J. Anim. Sci. 2018, 8, 137–150. [Google Scholar] [CrossRef]
- Faccin, J.E.G.; Laskoski, F.; Hernig, L.F.; Kummer, R.; Lima, G.F.R.; Orlando, U.A.D.; Goncalves, M.A.D.; Mellagi, A.P.G.; Ulguim, R.R.; Bortolozzo, F.P. Impact of Increasing Weaning Age on Pig Performance and Belly Nosing Prevalence in a Commercial Multisite Production System. J. Anim. Sci. 2020, 98, skaa031. [Google Scholar] [CrossRef] [PubMed]
- Faccin, J.E.G.; Tokach, M.D.; Allerson, M.W.; Woodworth, J.C.; Derouchey, J.M.; Dritz, S.S.; Bortolozzo, F.P.; Goodband, R.D. Relationship between Weaning Age and Antibiotic Usage on Pig Growth Performance and Mortality. J. Anim. Sci. 2020, 98, skaa363. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bortolozzo, F.P.; Zanin, G.P.; Ulguim, R.d.R.; Mellagi, A.P.G. Managing Reproduction in Hyperprolific Sow Herds. Animals 2023, 13, 1842. https://doi.org/10.3390/ani13111842
Bortolozzo FP, Zanin GP, Ulguim RdR, Mellagi APG. Managing Reproduction in Hyperprolific Sow Herds. Animals. 2023; 13(11):1842. https://doi.org/10.3390/ani13111842
Chicago/Turabian StyleBortolozzo, Fernando Pandolfo, Gabriela Piovesan Zanin, Rafael da Rosa Ulguim, and Ana Paula Gonçalves Mellagi. 2023. "Managing Reproduction in Hyperprolific Sow Herds" Animals 13, no. 11: 1842. https://doi.org/10.3390/ani13111842