
Haemonchus contortus Adopt Isolate-Specific Life History Strategies to Optimize Fitness and Overcome Obstacles in Their Environment: Experimental Evidence

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Design
2.3. Haemonchus Contortus Isolates
2.4. Sheep
2.5. Haemonchus Contortus Experiment
2.6. Pathogenicity
2.7. Faecal Nematode Egg Counts (FEC)
2.8. Egg-L3 Larvae Development
2.9. Establishment
2.10. Fertility
2.11. Parasite Fitness
2.12. Statistical Analyses
2.13. RNA Extraction and Double Strand cDNA Synthesis on H. contortus Adult Males
2.14. cDNA-AFLP Experiments
2.15. Transcriptomic Expression Profiles
3. Results
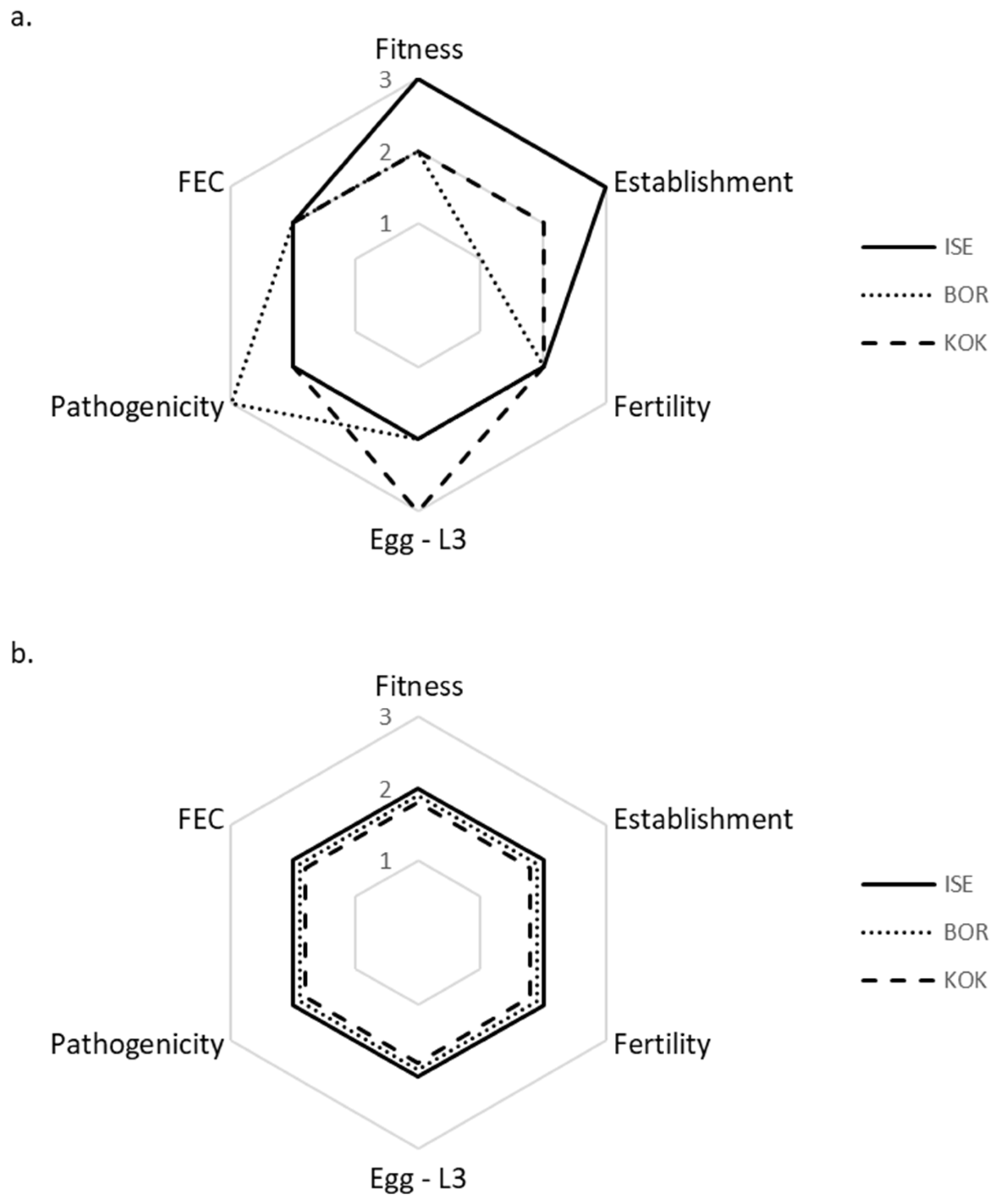
3.1. Inter-Isolate Comparison of Life History Strategies and Fitness in Susceptible Lambs × Spring Conditions
3.2. Inter-Isolate Comparison of Life History Strategies and Fitness in Resistant Rams × Spring Conditions
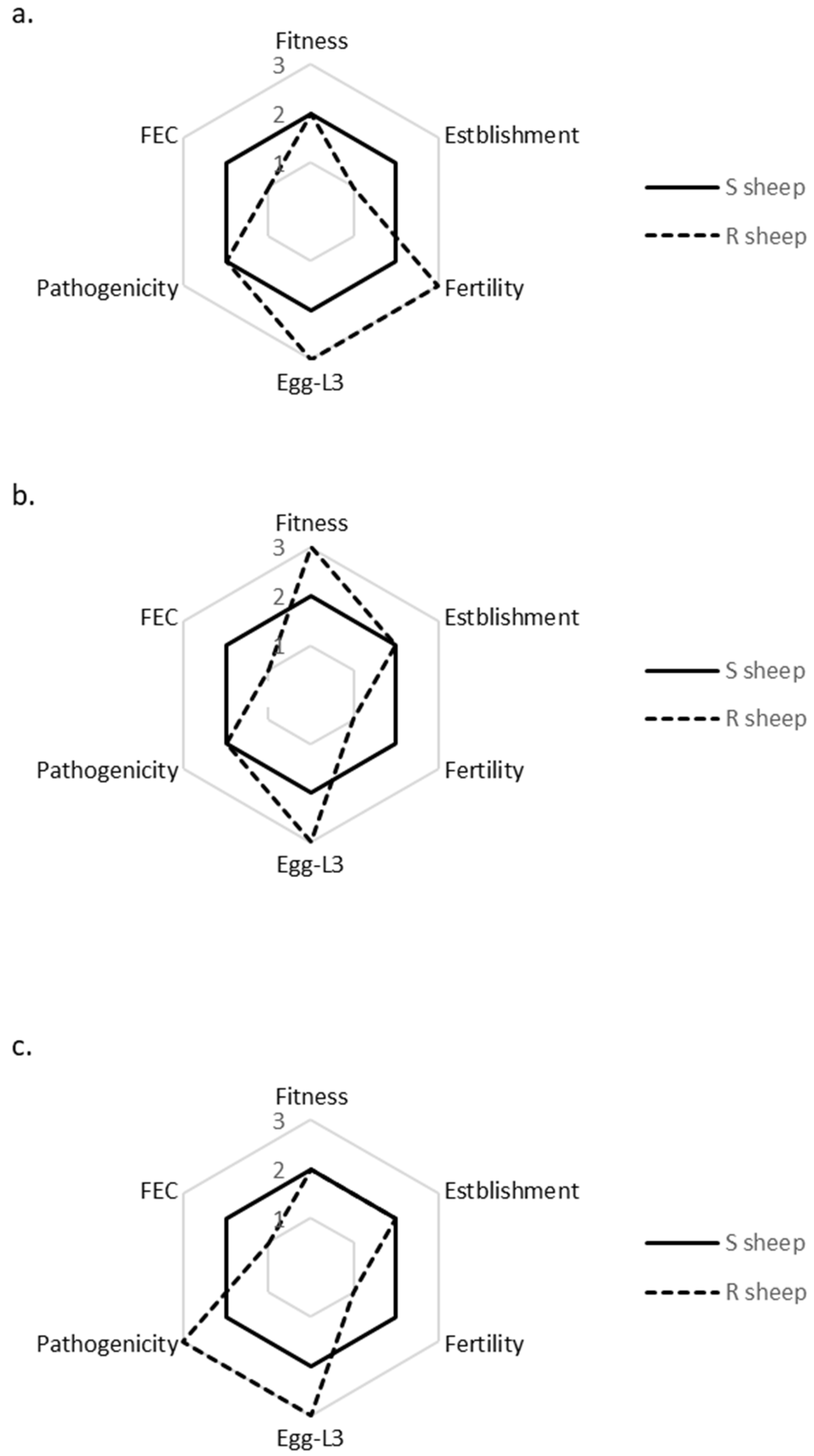
3.3. Intra-Isolate Comparison of Life History Strategies and Fitness in Susceptible vs. Resistant Sheep × Spring Conditions
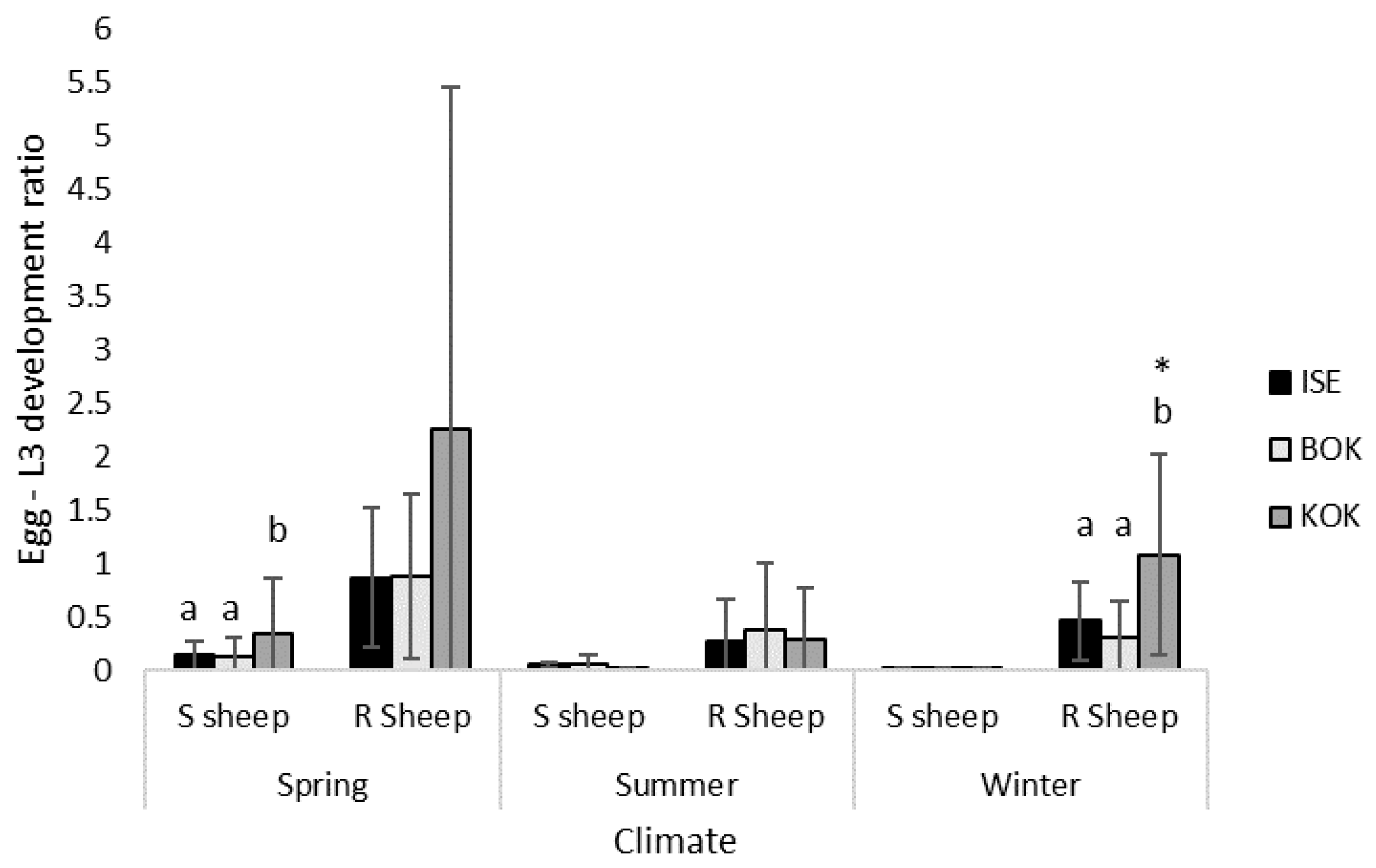
3.4. Isolate Egg-L3 Development under Different Climatic Conditions
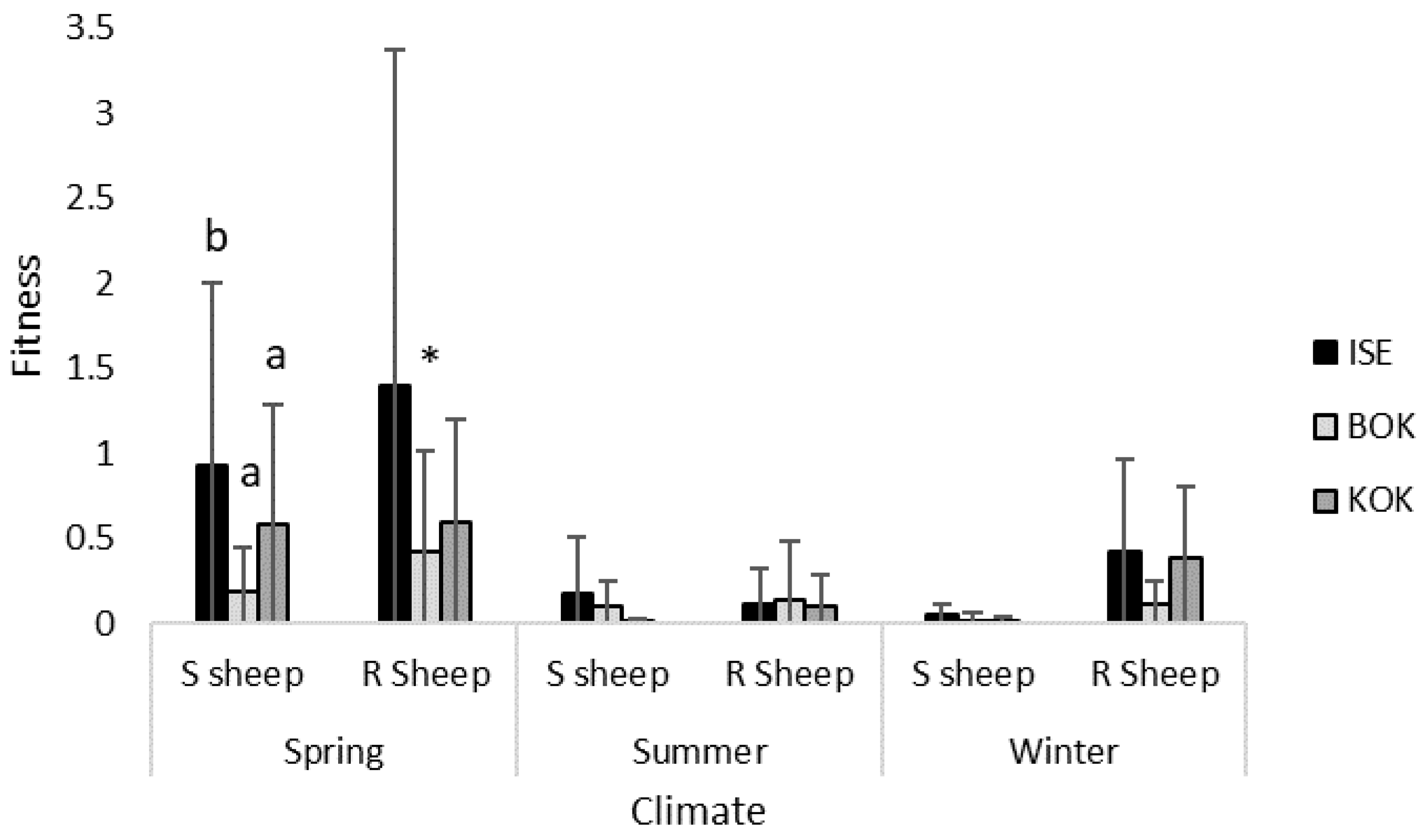
3.5. Isolate Experimental Fitness under Different Climatic Conditions
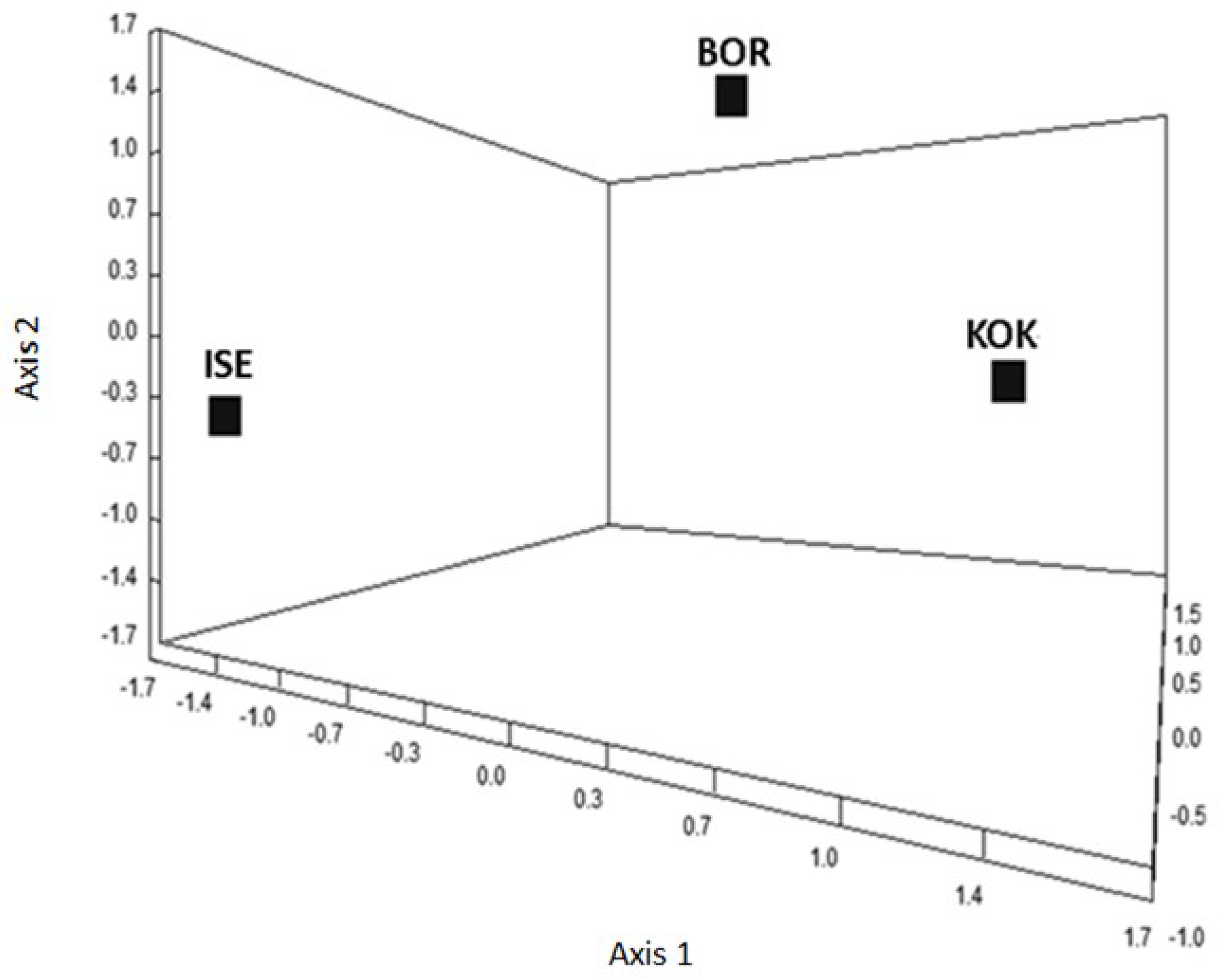
3.6. Genetic Analyses of the H. contortus Isolates
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilleard, J.S.; Kotze, A.C.; Leathwick, D.; Nisbet, A.J.; McNeilly, T.N.; Besier, B. A journey through 50 years of research relevant to the control of gastrointestinal nematodes in ruminant livestock and thoughts on future directions. Int. J. Parasitol. 2021, 51, 1133–1151. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, R.M. Biology, Epidemiology, Diagnosis, and Management of Anthelmintic Resistance in Gastrointestinal Nematodes of Livestock. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Sallé, G.; Doyle, S.R.; Cortet, J.; Cabaret, J.; Berriman, M.; Holroyd, N.; Cotton, J.A. The global diversity of Haemonchus contortus is shaped by human intervention and climate. Nat. Commun. 2019, 10, 4811. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.R.; Wall, R. Climate change and parasitic disease: Farmer mitigation? Trends Parasitol. 2009, 25, 308–313. [Google Scholar] [CrossRef]
- Sweeney, T.; Hanrahan, J.P.; Ryan, M.T.; Good, B. Immunogenomics of gastrointestinal nematode infection in ruminants—Breeding for resistance to produce food sustainably and safely. Parasite Immunol. 2016, 38, 569–586. [Google Scholar] [CrossRef]
- Viney, M.; Cable, J. Macroparasite life histories. Curr. Biol. 2011, 21, R767–R774. [Google Scholar] [CrossRef]
- Ekino, T.; Kirino, H.; Kanzaki, N.; Shinya, R. Ultrastructural plasticity in the plant-parasitic nematode, Bursaphelenchus xylophilus. Sci. Rep. 2020, 10, 11576. [Google Scholar] [CrossRef]
- Poulin, R. Evolutionary Ecology of Parasites—From Individuals to Communities, 1st ed.; Chapman and Hall: London, UK, 1998; pp. 46–65. [Google Scholar]
- Fisher, R.A. The Genetical Theory of Natural Selection; Oxford Clarendon Press: Oxford, UK, 1930. [Google Scholar]
- Haldane, J.B.S. The Causes of Evolution; London Longmans: London, UK, 1932. [Google Scholar]
- Hartl, D.L.; Clark, A.G. Principles of Population Genetics, 2nd ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 1989; pp. 147–155. [Google Scholar]
- Levins, R. Evolution in Changing Environments; Princeton University Press: Princeton, NJ, USA, 1968. [Google Scholar]
- Callow, P. The cost of reproduction: A physiological approach. Biol. Rev. 1979, 54, 23–40. [Google Scholar] [CrossRef]
- Chylinski, C.; Lherminé, E.; Coquille, M.; Cabaret, J. Desiccation tolerance of gastrointestinal nematode third-stage larvae: Exploring the effects on survival and fitness. Parasitol. Res. 2014, 113, 2789–2796. [Google Scholar] [CrossRef]
- Aumont, G.; Gruner, L.; Hostache, G. Comparison of the resistance to sympatric and allopatric isolates of Haemonchus contortus of Black Belly sheep in Guadeloupe (FWI) and of INRA 401 sheep in France. Vet. Parasitol. 2003, 116, 139–150. [Google Scholar] [CrossRef]
- Troell, K.; Tingstedt, C.; Höglund, J. Phenotypic characterization of Haemonchus contortus: A study of isolates from Sweden and Kenya in experimentally infected sheep. Parasitology 2006, 132, 403–409. [Google Scholar] [CrossRef]
- Gasnier, N. Isolement du Nématode Teladorsagia circumcincta au Sein des Fermes Caprines. Influence sur la Variabilité Morphologique, Ecologique et Génétique. Ph.D. Thesis, Université F. Rabelais, Tours, France, 1994. [Google Scholar]
- Kelly, J.D.; Whitlock, H.V.; Thompson, H.G.; Hall, C.A.; Martin, I.C.; Le Jambre, L.F. Physiological characteristics of free-living and parasitic stages of strains of Haemonchus contortus, susceptible or resistant to benzimidazole anthelmintics. Res. Vet. Sci. 1978, 25, 376–385. [Google Scholar] [CrossRef]
- Maingi, N.; Scott, M.E.; Prichard, R.K. Effect of selection pressure for thiabendazole resistance on fitness of Haemonchus contortus in sheep. Parasitology 1990, 100, 327–335. [Google Scholar] [CrossRef]
- Scott, E.W.; Armour, J. Effect of development of resistance to benzimidazoles, salicylanilides and ivermectin on the pathogenicity and survival of Haemonchus contortus. Vet. Rec. 1991, 128, 346–349. [Google Scholar] [CrossRef]
- Echevarria, F.A.; Armour, J.; Borba, M.F.; Duncan, J.L. Survival and development of ivermectin-resistant or susceptible strains of Haemonchus contortus under field and laboratory conditions. Res. Vet. Sci. 1993, 54, 133–139. [Google Scholar] [CrossRef]
- Chehresa, A.; Beech, R.N.; Scott, M.E. Life-history variation among lines isolated from a laboratory population of Heligmosomoides polygyrus bakeri. Int. J. Parasitol. 1997, 27, 541–551. [Google Scholar] [CrossRef]
- Dobson, C.; Owen, M.E. Influence of serial passage on the infectivity and immunogenicity of Nematospiroides dubius in mice. Int. J. Parasitol. 1977, 7, 463–466. [Google Scholar] [CrossRef]
- Chylinski, C.; Cortet, J.; Sallé, G.; Jacquiet, P.; Cabaret, J. Storage of gastrointestinal nematode infective larvae for species preservation and experimental infections. Parasitol. Res. 2015, 114, 715–720. [Google Scholar] [CrossRef]
- Hoekstra, R.; Borgsteede, F.H.; Boersema, J.H.; Roos, M.H. Selection for high levamisole resistance in Haemonchus contortus monitored with an egg-hatch assay. Int. J. Parasitol. 1997, 27, 1395–1400. [Google Scholar] [CrossRef]
- Roos, M.H.; Otsen, M.; Hoekstra, R.; Veenstra, J.G.; Lenstra, J.A. Genetic analysis of inbreeding of two strains of the parasitic nematode Haemonchus contortus. Int. J. Parasitol. 2004, 34, 109–115. [Google Scholar] [CrossRef]
- Terefe, G.; Lacroux, C.; Andreoletti, O.; Grisez, C.; Prevot, F.; Bergeaud, J.P.; Penicaud, J.; Rouillon, V.; Gruner, L.; Brunel, J.C.; et al. Immune response to Haemonchus contortus infection in susceptible (INRA 401) and resistant (Barbados Black Belly) breeds of lambs. Parasite Immunol. 2007, 29, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Koski, K.G.; Scott, M.E. Gastrointestinal nematodes, nutrition and immunity: Breaking the negative spiral. Ann. Rev. Nutr. 2001, 21, 297–321. [Google Scholar] [CrossRef] [PubMed]
- Raynaud, J.P. Etude de l’efficacité d’une technique de coproscopie quantitative pour le diagnostic de routine et le contrôle des infestations parasitaires des bovins, ovins, équins et porcins. Ann. Parasitol. Hum. Comp. 1970, 45, 321–342. [Google Scholar] [CrossRef] [PubMed]
- Fleming, M.W. Size of inoculum dose regulates in part worm burdens, fecundity, and lengths in ovine Haemonchus contortus infections. J. Parasitol. 1988, 74, 975–978. [Google Scholar] [CrossRef] [PubMed]
- Rossanigo, C.E. Rôle de L’eau et de la Température sur les Taux de Développement des Nematodes Parasites du Tractus Digestif des Ruminants. Ph.D. Thesis, Université Montpellier, Montpellier, France, 1992. [Google Scholar]
- Baermann, G. Eine Einfach Methode zur Auffindung von Ankylostomun—(Nematoda)—Larven in Erdproben; Welteureden Batavia Geneesk. Lab. Feestbundel; Javasche Boekhandel & Drukkerij: Batavia, Germany, 1917; pp. 41–47. [Google Scholar]
- Gaba, S.; Chadoeuf, J.; Monestiez, P.; Sauve, C.; Cortet, J.; Cabaret, J. Estimation of abomasum strongyle nematode infections in sheep at necropsy: Tentative proposals for a simplified technique. Vet. Parasitol. 2006, 140, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Cabaret, J.; Ouhelli, H. Fertility of parasitic strongyles in the digestive system of sheep under natural conditions. Rev. Méd. Vétérinaire 1984, 135, 627–633. [Google Scholar]
- Chylinski, C.; Schmidt, E.; Gruner, L.; Cabaret, J. Serial passage in resistant sheep drives the infectivity and fitness of Teladorsagia circumcincta in susceptible lambs: Experimental evidence. Parasitol. Int. 2022, 89, 102586. [Google Scholar] [CrossRef]
- Lebart, L.; Morineau, A.; Fénelon, J.P. Traitement des Données Statistiques; Méthodes et Programmes; Dunod: Paris, France, 1982. [Google Scholar]
- Blanquart, F.; Kaltz, O.; Nuismer, S.L.; Gandon, S. A practical guide to measuring local adaptation. Ecol. Lett. 2013, 16, 1195–1205. [Google Scholar] [CrossRef]
- de Almeida, A.M. Barbados Blackbelly: The Caribbean ovine genetic resource. Trop. Anim. Health Prod. 2018, 50, 239–250. [Google Scholar] [CrossRef]
- Greer, A.W.; Sykes, A.R. Are faecal egg counts approaching their “sell-by” date? N. Z. Soc. Anim. Prod. 2012, 72, 199–204. [Google Scholar]
- Sabatini, G.A.; de Almeida Borges, F.; Claerebout, E.; Gianechini, L.S.; Höglund, J.; Kaplan, R.M.; Lopes, W.D.Z.; Mitchell, S.; Rinaldi, L.; von Samson-Himmelstjerna, G.; et al. Practical guide to the diagnostics of ruminant gastrointestinal nematodes, liver fluke and lungworm infection: Interpretation and usability of results. Parasites Vectors 2023, 16, 58. [Google Scholar] [CrossRef]
- Morgan, E.R.; Cavill, L.; Curry, G.E.; Wood, R.M.; Mitchell, E.S. Effects of aggregation and sample size on composite faecal egg counts in sheep. Vet. Parasitol. 2005, 131, 79–87. [Google Scholar] [CrossRef]
- Rossanigo, C.E.; Gruner, L. Accuracy of two methods for counting eggs of sheep nematode parasites. Vet. Parasitol. 1991, 39, 115–121. [Google Scholar] [CrossRef]
- Stear, M.J.; Bishop, S.C. The curvilinear relationship between worm length and fecundity of Teladorsagia circumcincta. Int. J. Parasitol. 1999, 29, 777–780. [Google Scholar] [CrossRef]
- Harvey, S.C.; Orbidans, H.E. All eggs are not equal: The maternal environment affects progeny reproduction and developmental fate in Caenorhabditis elegans. PLoS ONE 2011, 6, e25840. [Google Scholar] [CrossRef]
- Otsen, M.; Hoekstra, R.; Plas, M.E.; Buntjer, J.B.; Lenstra, J.A.; Roos, M.H. Amplified fragment length polymorphism analysis of genetic diversity of Haemonchus contortus during selection for drug resistance. Int. J. Parasitol. 2001, 31, 1138–1143. [Google Scholar] [CrossRef]
- Xu, M.; Molento, M.; Blackhall, W.; Ribeiro, P.; Beech, R.; Prichard, R. Ivermectin resistance in nematodes may be caused by alteration of P-glycoprotein homolog. Mol. Biochem Parasitol. 1998, 91, 327–335. [Google Scholar] [CrossRef]
- Dicker, A.J.; Nath, M.; Yaga, R.; Nisbet, A.J.; Lainson, F.A.; Gilleard, J.S.; Skuce, P.J. Teladorsagia circumcincta: The transcriptomic response of a multi-drug-resistant isolate to ivermectin exposure in vitro. Exp. Parasitol. 2011, 127, 351–356. [Google Scholar] [CrossRef]
- El-Abdellati, A.; De Graef, J.; Van Zeveren, A.; Donnan, A.; Skuce, P.; Walsh, T.; Wolstenholme, A.; Tait, A.; Vercruysse, J.; Claerebout, E.; et al. Altered avr-14B gene transcription patterns in ivermectin-resistant isolates of the cattle parasites, Cooperia oncophora and Ostertagia ostertagi. Int. J. Parasitol. 2011, 41, 951–957. [Google Scholar] [CrossRef]
- Williamson, S.M.; Storey, B.; Howell, S.; Harper, K.M.; Kaplan, R.M.; Wolstenholme, A.J. Candidate anthelmintic resistance-associated gene expression and sequence polymorphisms in a triple-resistant field isolate of Haemonchus contortus. Mol. Biochem. Parasitol. 2011, 180, 99–105. [Google Scholar] [CrossRef]
- Kahn, L.P.; Woodgate, R.G. Integrated parasite management: Products for adoption by the Australian sheep industry. Vet. Parasitol. 2012, 186, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Maqbool, I.; Wani, Z.A.; Shahardar, R.A.; Allaie, I.M.; Shah, M.M. Integrated parasite management with special reference to gastro-intestinal nematodes. J. Parasit Dis. 2017, 41, 1–8. [Google Scholar] [CrossRef] [PubMed]
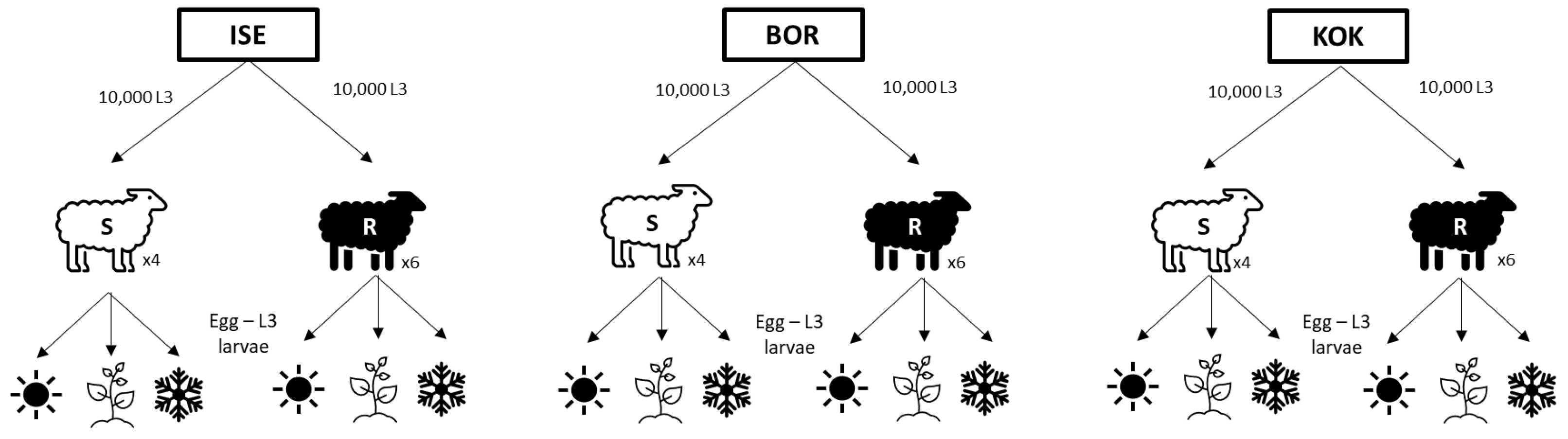
 (×4) and resistant
(×4) and resistant  sheep (×6) using an infective dose of 10,000 L3 larvae; eggs excreted from the sheep were collected a cultured under conditions reflecting summer
sheep (×6) using an infective dose of 10,000 L3 larvae; eggs excreted from the sheep were collected a cultured under conditions reflecting summer  , spring
, spring  and winter
and winter  .
(×4) and resistant sheep (×6) using an infective dose of 10,000 L3 larvae; eggs excreted from the sheep were collected a cultured under conditions reflecting summer , spring and winter .
.
(×4) and resistant sheep (×6) using an infective dose of 10,000 L3 larvae; eggs excreted from the sheep were collected a cultured under conditions reflecting summer , spring and winter .






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Country of Origin | Years Maintained in Lab (INRAE) | Anthelmintic Resistance Status | Selection of Anthelmintic Resistance | Prior Experience of Resistant Sheep | Line History Reference |
|---|---|---|---|---|---|---|
| ISE | NA (inbred isolate) | 25+ | Susceptible | NA | No | [26] |
| BOR | Netherlands | 10 | Levamisole | Experimentally | No | [25] |
| KOK | South Africa | 18 | Levamisole, Avermectin, Benzimidazole | On field | No | Donated by J. Van Wyk, 2000 |
| Primer Name | 5′-3′ |
|---|---|
| M + 0 | GATGAGTCCTGAGTAA |
| H + 0 | GACTGCGTACCAGCTT |
| H + A | GACTGCGTACCAGCTTA |
| H + T | GACTGCGTACCAGCTTT |
| M12 | GATGAGTCCTGAGTAAAC |
| M14 | GATGAGTCCTGAGTAAAT |
| M15 | GATGAGTCCTGAGTAACA |
| M16 | GATGAGTCCTGAGTAACC |
| M17 | GATGAGTCCTGAGTAACG |
| M18 | GATGAGTCCTGAGTAACT |
| M19 | GATGAGTCCTGAGTAAGA |
| M20 | GATGAGTCCTGAGTAAGC |
| M23 | GATGAGTCCTGAGTAATA |
| M24 | GATGAGTCCTGAGTAATC |
| M25 | GATGAGTCCTGAGTAATG |
| M26 | GATGAGTCCTGAGTAATT |
| Susceptible Sheep | Resistant Sheep | |||||
|---|---|---|---|---|---|---|
| ISE | BOR | KOK | ISE | BOR | KOK | |
| Fitness | ||||||
| Spring | 0.93 (±1.07) | 0.19 (±0.26) | 0.58 (±0.70) | 1.40 (±1.97) | 0.42 (±0.59) | 0.59 (±0.61) |
| Summer | 0.18 (±0.33) | 0.10 (±0.15) | 0.01 (±0.02) | 0.12 (±0.21) | 0.14 (±0.35) | 0.10 (±0.19) |
| Winter | 0.05 (±0.07) | 0.02 (±0.05) | 0.02 (±0.02) | 0.42 (±0.55) | 0.11 (±0.14) | 0.38 (±0.42) |
| Establishment | 4314 (±2532) | 119 (±83) | 1263 (±814) | 468 (±632) | 170 (±270) | 683 (±582) |
| Fertility | 17 (±16) | 148 ±205 | 162 (±183) | 74 (±97) | 8 (±6) | 36 (±70) |
| Egg-L3 | ||||||
| Spring | 0.14 (±0.14) | 0.12 (±0.19) | 0.35 (±0.52) | 0.87 (±0.66) | 0.88 (±0.77) | 2.25 (±3.21) |
| Summer | 0.05 (±0.02) | 0.06 (±0.09) | 0.01 (±0.02) | 0.27 (±0.40) | 0.38 (±0.62) | 0.29 (±0.48) |
| Winter | 0.01 (±0.02) | 0.01 (±0.01) | 0.01 (±0.01) | 0.46 (±0.37) | 0.31 (±0.34) | 1.08 (±0.94) |
| Pathogenicity | 19 (±6) | 27 (±4) | 24 (±2) | 27 (±5) | 30 (±1) | 28 (±1) |
| FEC | 11,299 (±10,166) | 2940 (±5156) | 4290 (±3581) | 1765 (±2233) | 257 (±395) | 517 (±684) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chylinski, C.; Cortet, J.; Cabaret, J.; Blanchard, A. Haemonchus contortus Adopt Isolate-Specific Life History Strategies to Optimize Fitness and Overcome Obstacles in Their Environment: Experimental Evidence. Animals 2023, 13, 1759. https://doi.org/10.3390/ani13111759
Chylinski C, Cortet J, Cabaret J, Blanchard A. Haemonchus contortus Adopt Isolate-Specific Life History Strategies to Optimize Fitness and Overcome Obstacles in Their Environment: Experimental Evidence. Animals. 2023; 13(11):1759. https://doi.org/10.3390/ani13111759
Chicago/Turabian StyleChylinski, Caroline, Jacques Cortet, Jacques Cabaret, and Alexandra Blanchard. 2023. "Haemonchus contortus Adopt Isolate-Specific Life History Strategies to Optimize Fitness and Overcome Obstacles in Their Environment: Experimental Evidence" Animals 13, no. 11: 1759. https://doi.org/10.3390/ani13111759