
Is Oral Vaccination against Escherichia coli Influenced by Zinc Oxide?
 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
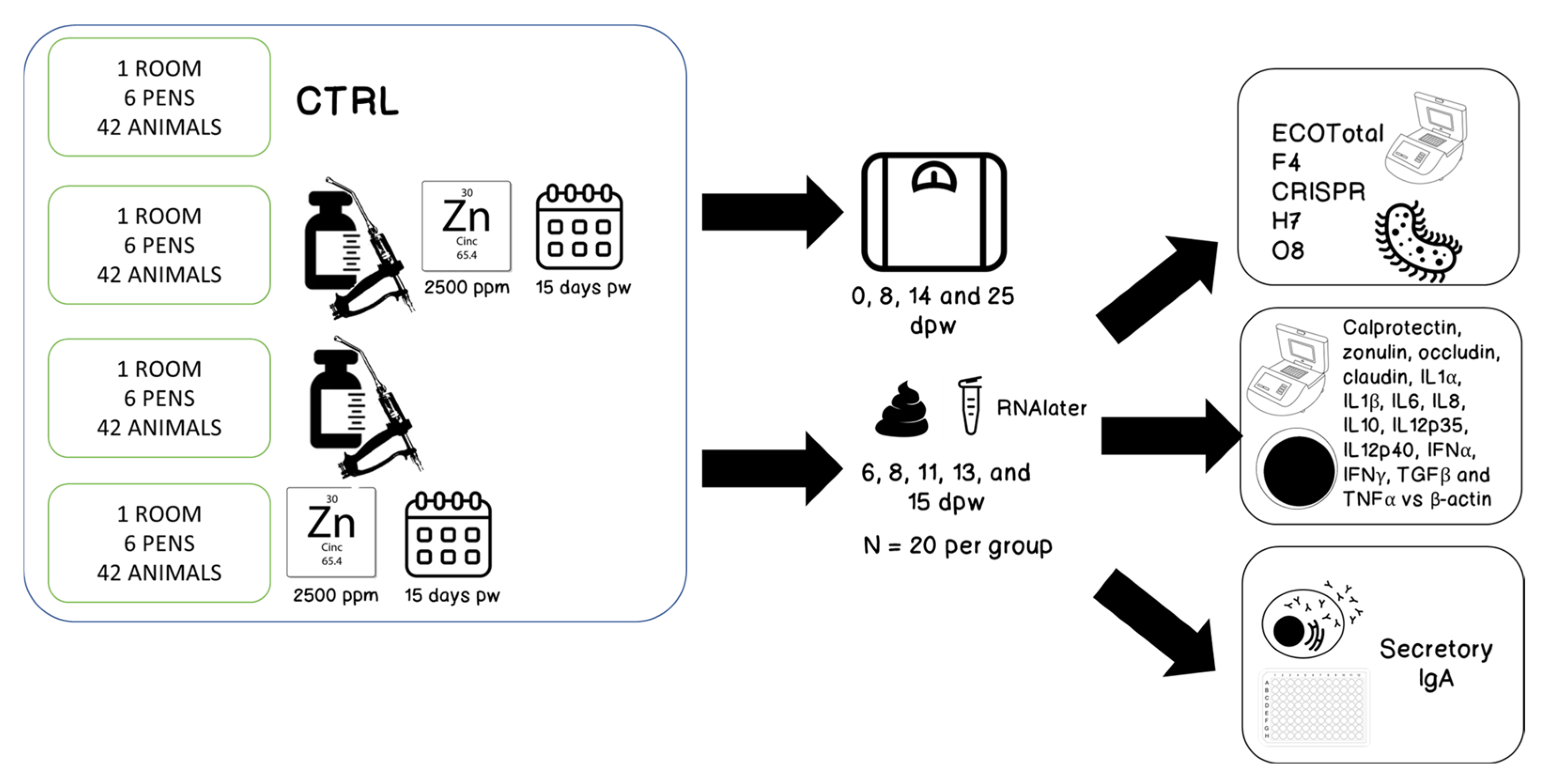
2. Materials and Methods
2.1. Animals
2.2. Vaccine and Vaccination
2.3. Feeds
2.4. Weighing of Animals
2.5. Faecal Sampling
2.6. Isolation of Nucleic Acids
2.7. Quantification of E. coli
2.8. Gene Expression for Intestinal Integrity and Immune Stimulation
2.9. Secretory IgA Quantification
2.10. Statistical Analysis
3. Results
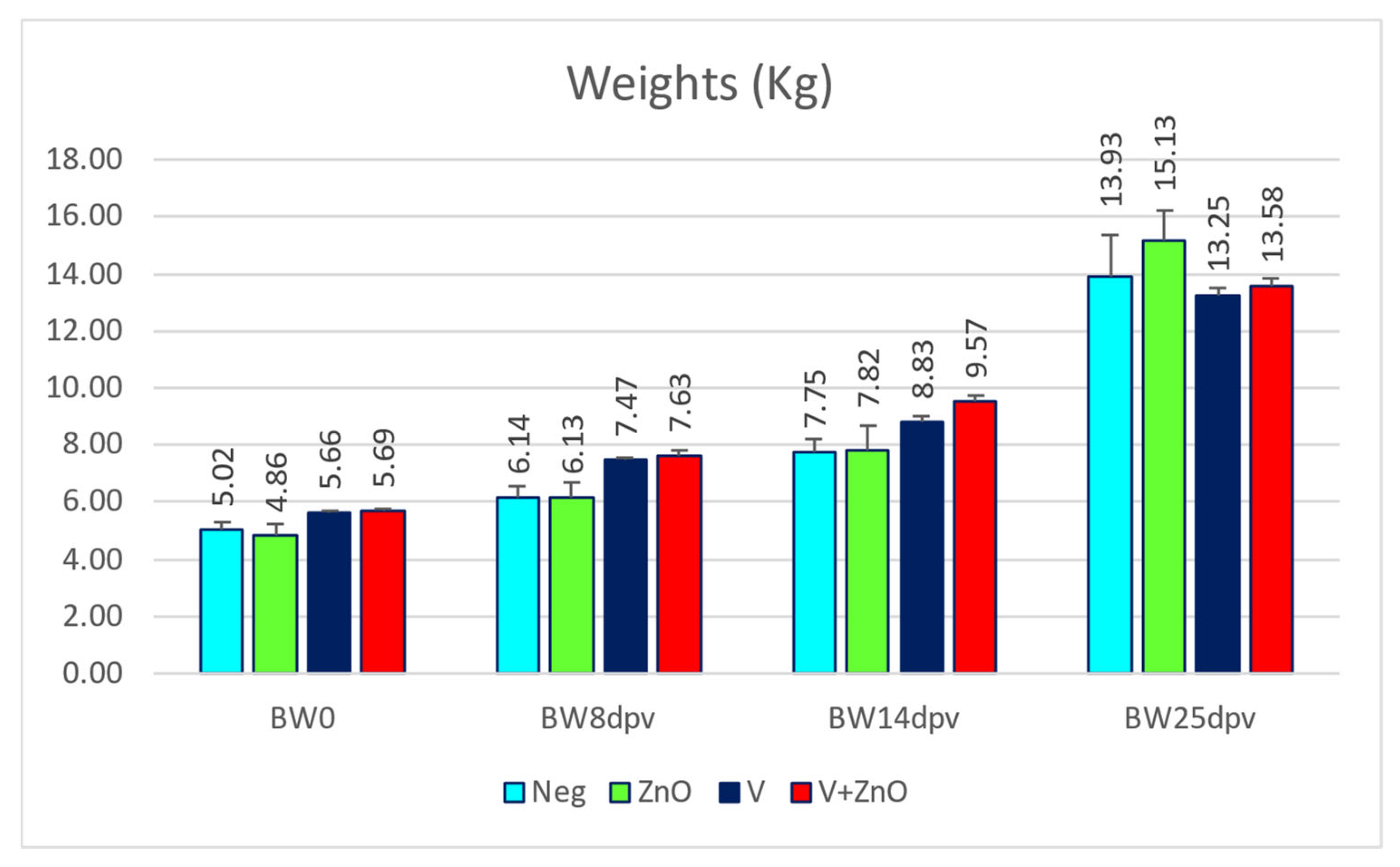
3.1. Weights and Growth
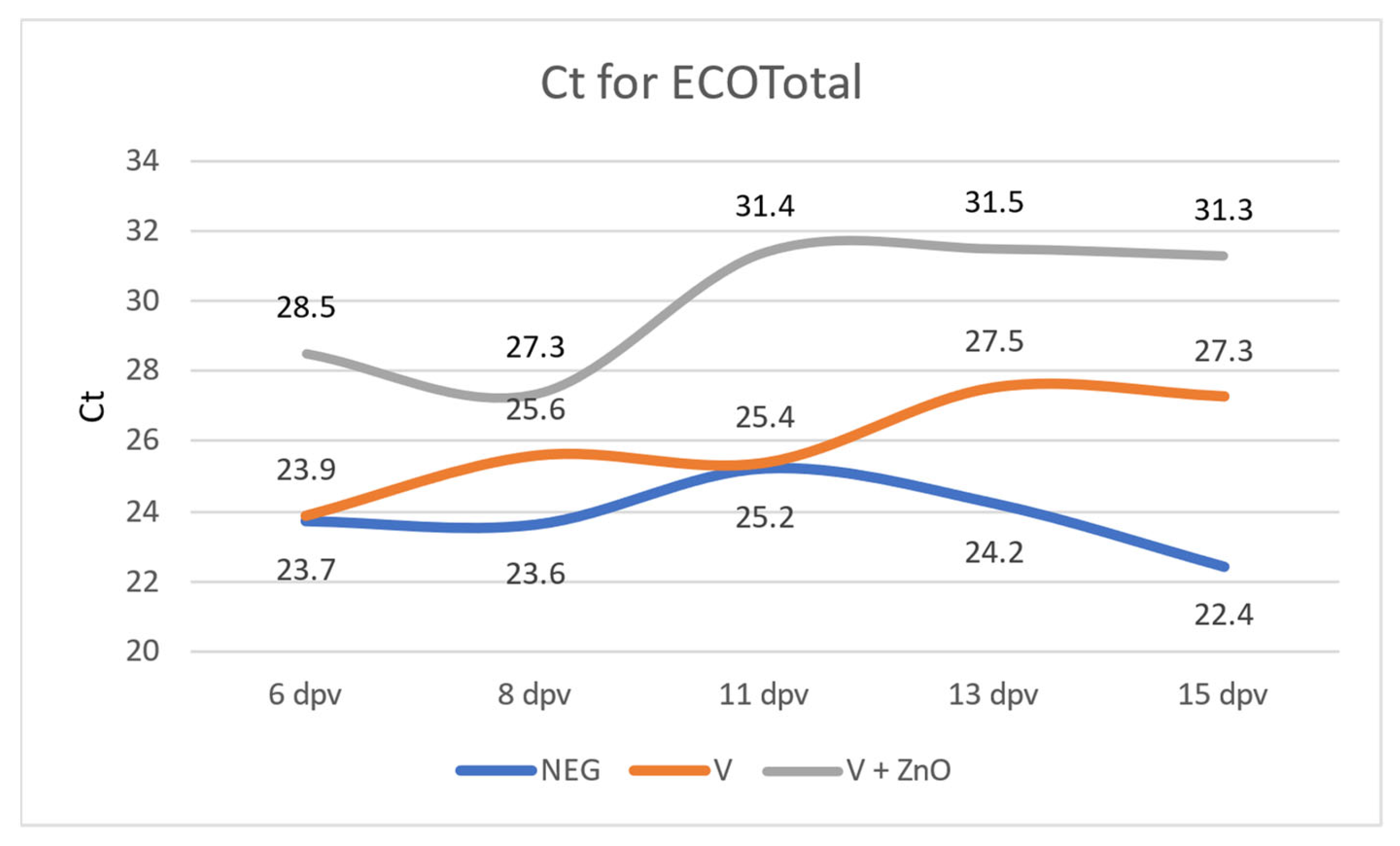
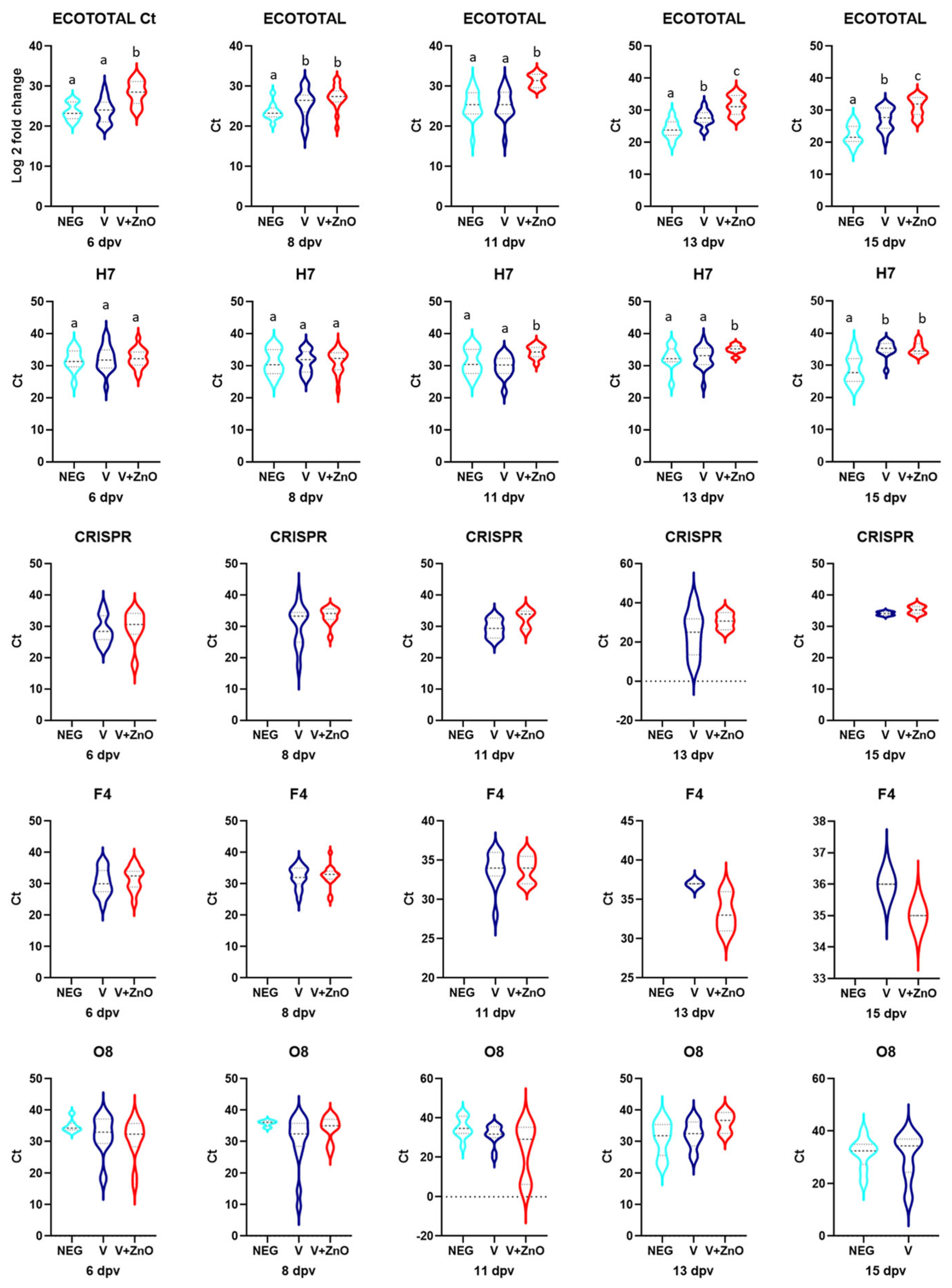
3.2. Quantification of E. coli
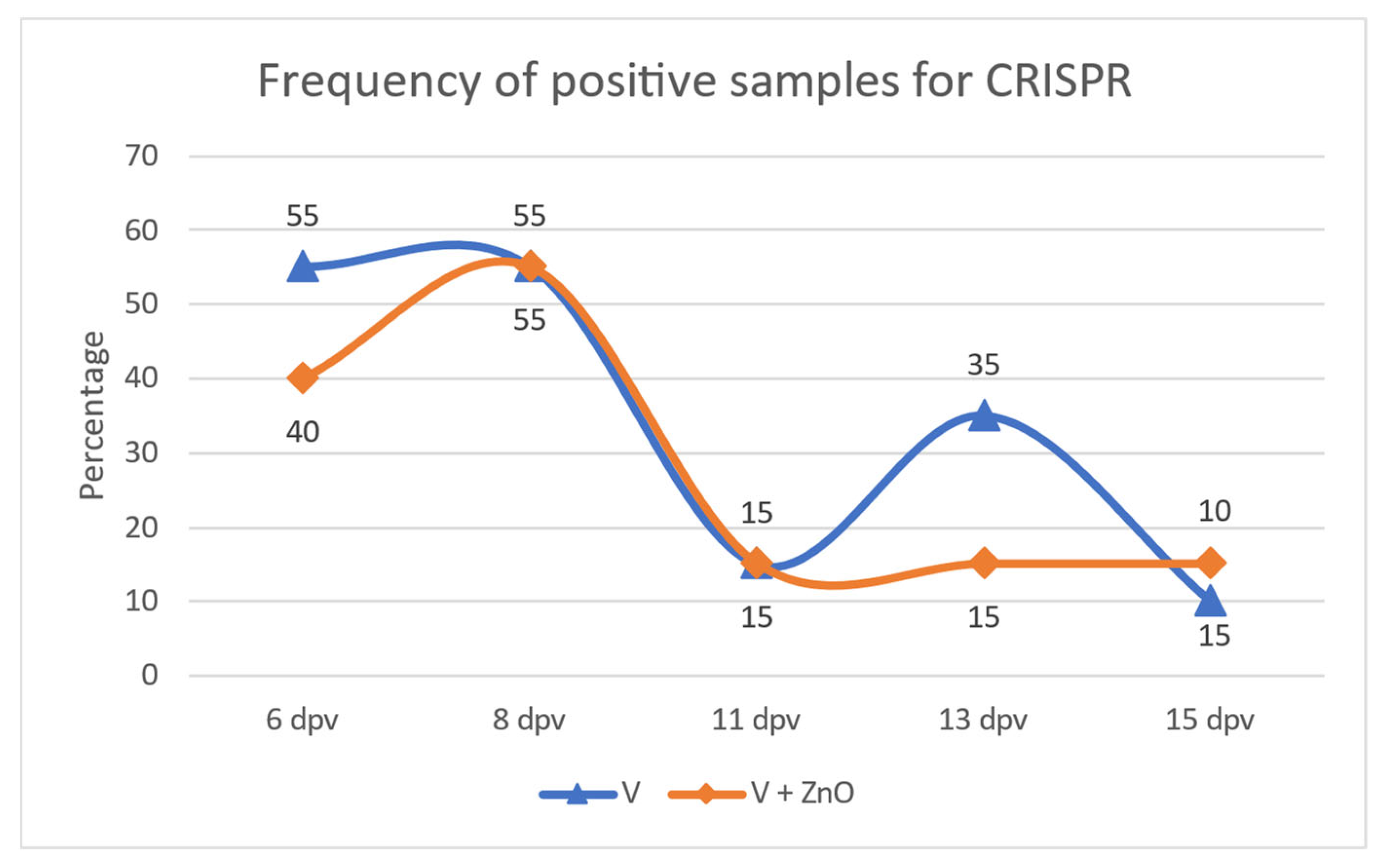
3.2.1. Frequency of Positive Samples
3.2.2. Quantification of E. coli Genes
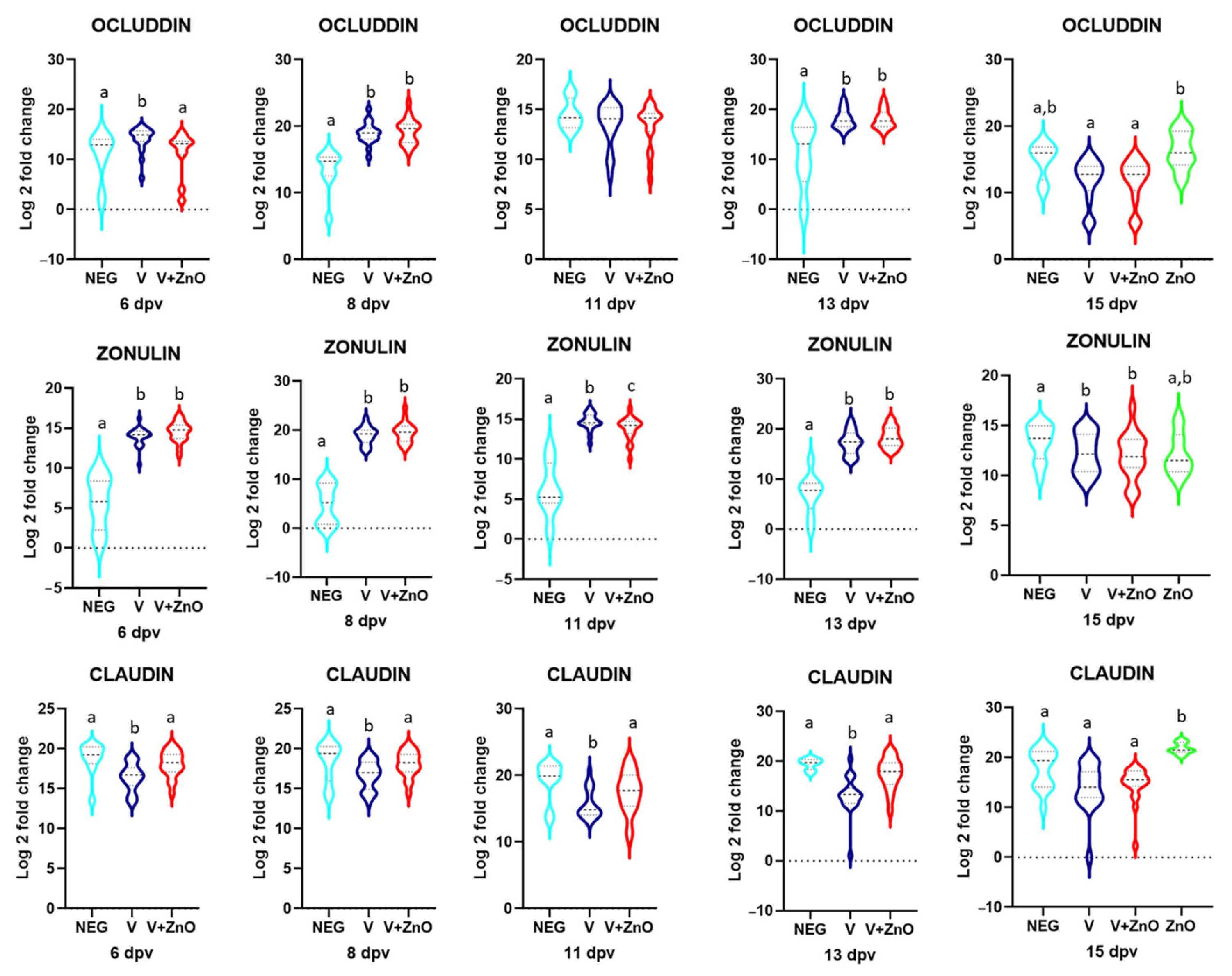
3.3. Intestinal Integrity
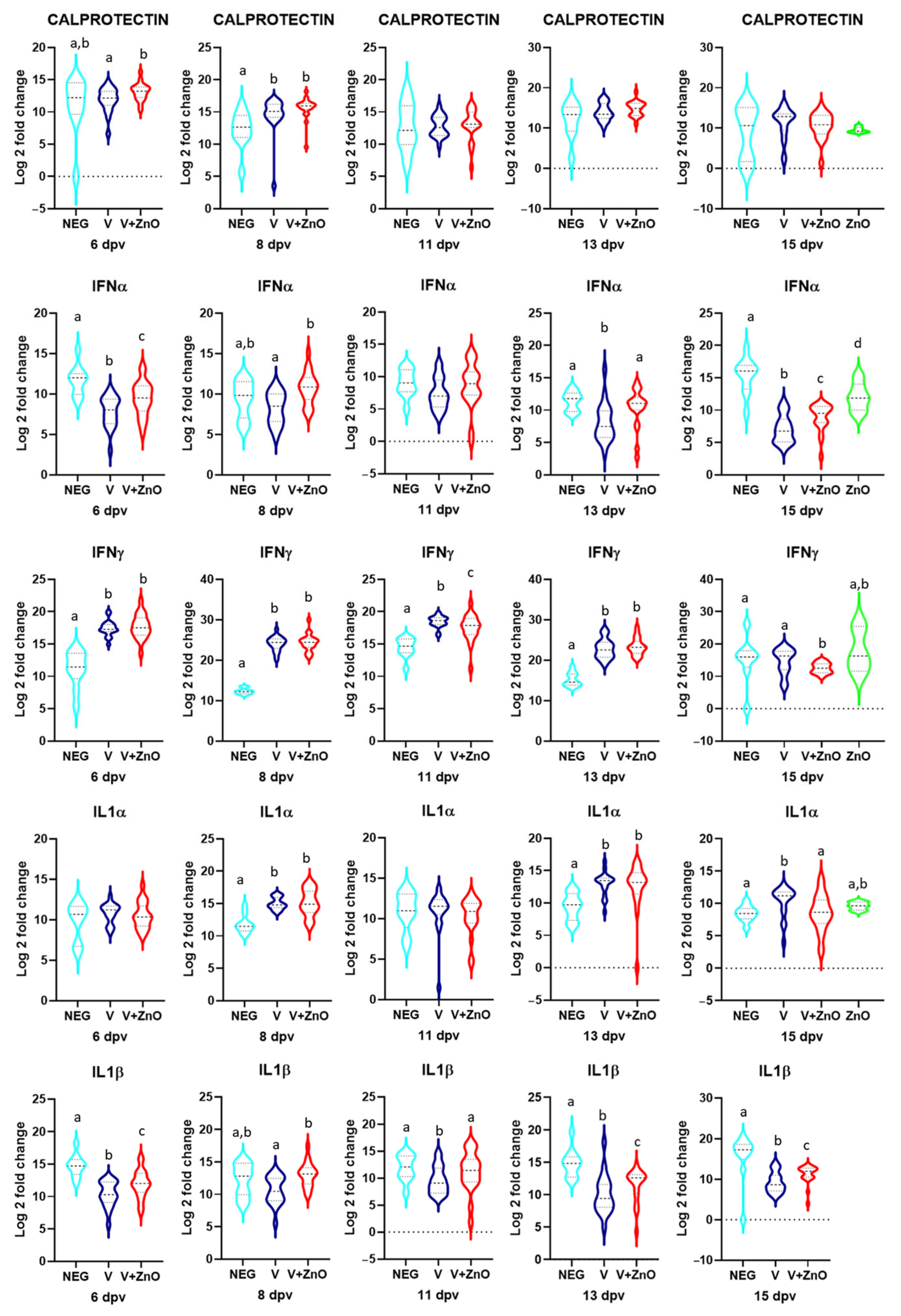
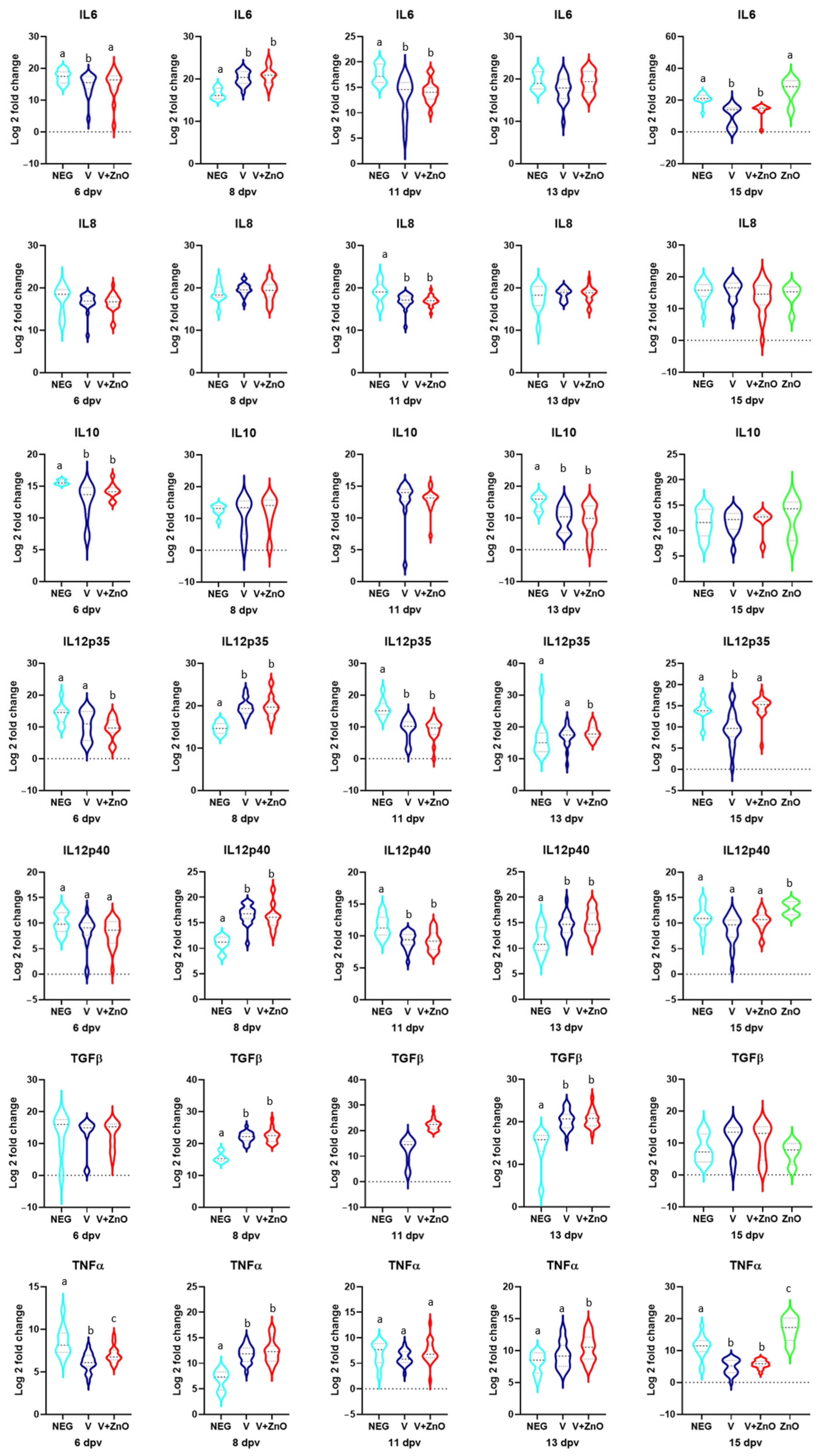
3.4. Immune Stimulation
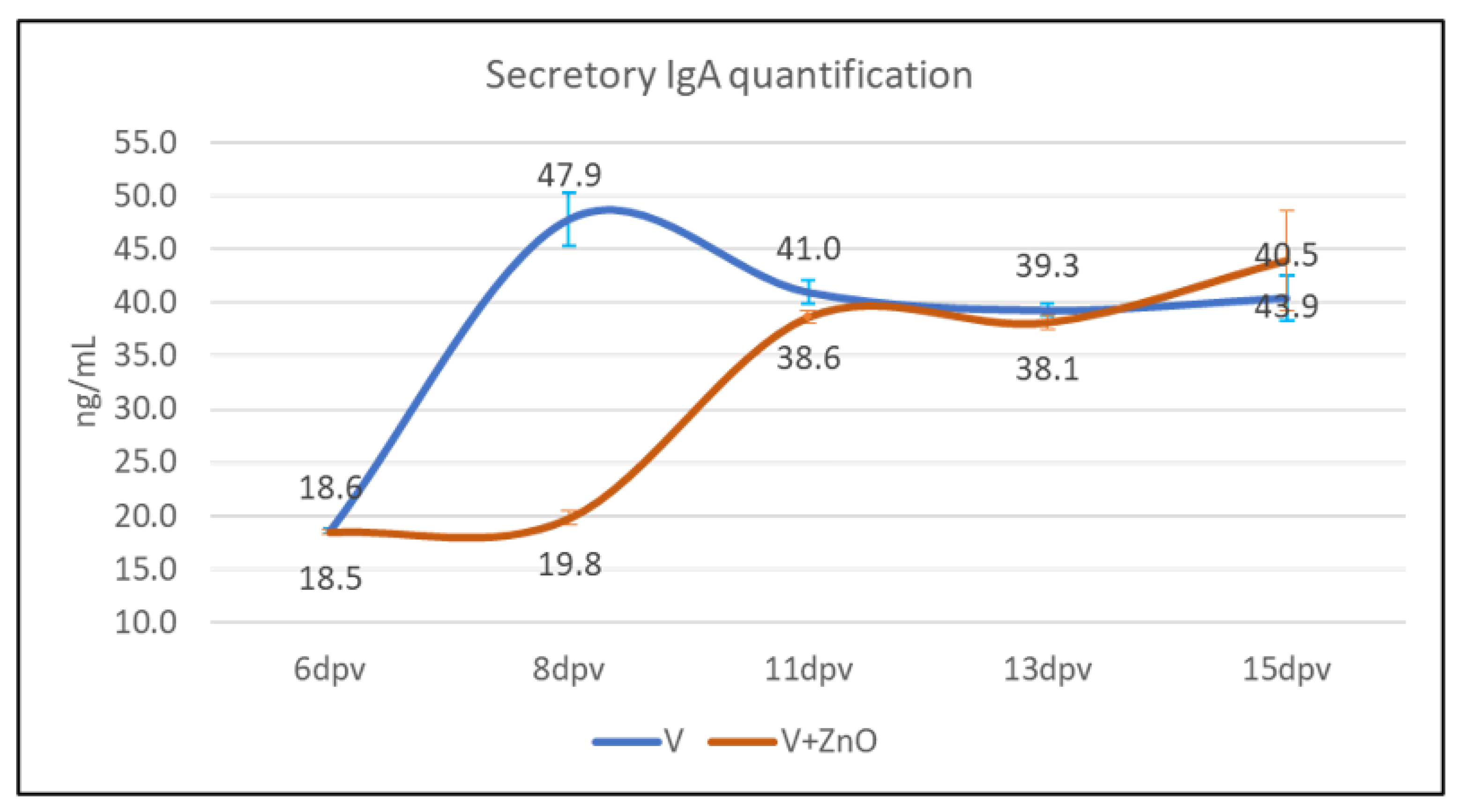
3.5. Secretory IgA Quantification
3.6. Correlations among Parameters
3.6.1. Correlations among E. coli Gene Quantification and Performances
3.6.2. Correlations among E. coli Gene Quantification and Immune Stimulation Gene Expression
3.6.3. Correlations among E. coli Genes and Intestinal Integrity
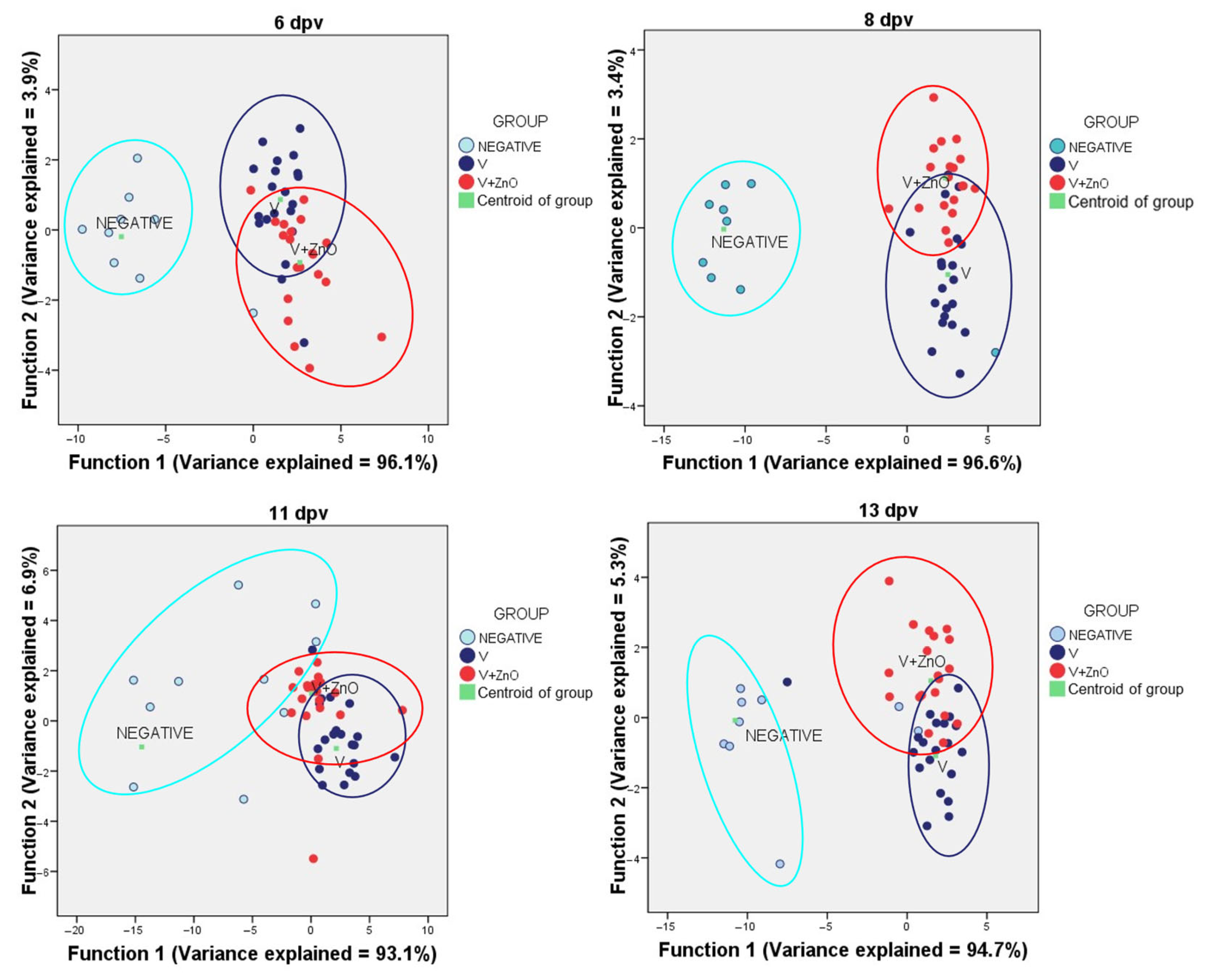
3.7. Data Reduction including Immune System and Intestinal Integrity Biomarkers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Groot, N.; Fariñas, F.; Cabrera-Gómez, C.G.; Pallares, F.J.; Ramis, G. Weaning Causes a Prolonged but Transient Change in Immune Gene Expression in the Intestine of Piglets. J. Anim. Sci. 2021, 99, skab065. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Duarte, M.E. Understanding Intestinal Health in Nursery Pigs and the Relevant Nutritional Strategies. Anim. Biosci. 2021, 34, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Rhouma, M.; Fairbrother, J.M.; Beaudry, F.; Letellier, A. Post Weaning Diarrhea in Pigs: Risk Factors and Non-Colistin-Based Control Strategies. Acta Vet. Scand. 2017, 59, 31. [Google Scholar] [CrossRef] [PubMed]
- Fairbrother, J.M.; Nadeau, É.; Gyles, C.L. Escherichia coli in Postweaning Diarrhea in Pigs: An Update on Bacterial Types, Pathogenesis, and Prevention Strategies. Anim. Health Res. Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef]
- Ramis, G.; Carrasco, L.; Pallarés, F.J.; Astorga, R.; Muñoz, A.; Gómez-Laguna, J. Capítulo 4. In Patologías Digestivas Porcinas en Imágenes; Servet: Zaragoza, Spain, 2011; pp. 110–125. [Google Scholar]
- Kloubert, V.; Blaabjerg, K.; Dalgaard, T.S.; Poulsen, H.D.; Rink, L.; Wessels, I. Influence of Zinc Supplementation on Immune Parameters in Weaned Pigs. J. Trace Elem. Med. Biol. 2018, 49, 231–240. [Google Scholar] [CrossRef]
- European Medicines Agency. Committee for Medicinal Products for Veterinary Use (CVMP) Meeting of 13–15 June 2017 CVMP Opinions on Veterinary Medicinal Products; European Medicines Agency: London, UK, 2017; Volume 44, pp. 14–16. [Google Scholar]
- European Parliament and the Council of the European Union Regulation (EC) No 1831/2003. Off. J. Eur. Union 2003, 4, 29–43.
- Nadeau, E.; Fairbrother, J.M.; Zentek, J.; Bélanger, L.; Tremblay, D.; Tremblay, C.L.; Röhe, I.; Vahjen, W.; Brunelle, M.; Hellmann, K.; et al. Efficacy of a Single Oral Dose of a Live Bivalent E. coli Vaccine against Post-Weaning Diarrhea Due to F4 and F18-Positive Enterotoxigenic E. coli. Vet. J. 2017, 226, 32–39. [Google Scholar] [CrossRef]
- Vangroenweghe, F. Improved Piglet Performance and Reduced Mortality and Antimicrobial Use Following Oral Vaccination with a Live Non-Pathogenic E. coli F4/F18 Vaccine against PWD. Austin J. Infect. Dis. 2022, 8, 2–7. [Google Scholar] [CrossRef]
- Luise, D.; Spinelli, E.; Correa, F.; Salvarani, C.; Bosi, P.; Trevisi, P. Effects of E. coli Bivalent Vaccine and of Host Genetic Susceptibility to E. coli on the Growth Performance and Faecal Microbial Profile of Weaned Pigs. Livest. Sci. 2020, 241, 104247. [Google Scholar] [CrossRef]
- Ramis, G.; Pérez-Esteruelas, P.; Gómez-Cabrera, C.; De Pascual-Monreal, C.; Gonzalez-Guijarro, B.; Ros-Párraga, E.; Sánchez-Uribe, P.; Claver-mateos, M.; Mendonça-Pascoal, L.; Martínez-Alarcón, L. Oral and Parenteral Vaccination against Escherichia coli in Piglets Results in Different Responses. Animals 2022, 12, 2758. [Google Scholar] [CrossRef]
- Johanns, V.C.; Ghazisaeedi, F.; Epping, L.; Semmler, T.; Lübke-Becker, A.; Pfeifer, Y.; Bethe, A.; Eichhorn, I.; Merle, R.; Walther, B.; et al. Effects of a Four-Week High-Dosage Zinc Oxide Supplemented Diet on Commensal Escherichia coli of Weaned Pigs. Front. Microbiol. 2019, 10, 2734. [Google Scholar] [CrossRef] [PubMed]
- Wątły, J.; Potocki, S.; Rowińska-Żyrek, M. Zinc Homeostasis at the Bacteria/Host Interface—From Coordination Chemistry to Nutritional Immunity. Chem. Eur. J. 2016, 22, 15992–16010. [Google Scholar] [CrossRef] [PubMed]
- Stocks, C.J.; Phan, M.D.; Achard, M.E.S.; Nhu, N.T.K.; Condon, N.D.; Gawthorne, J.A.; Lo, A.W.; Peters, K.M.; McEwan, A.G.; Kapetanovic, R.; et al. Uropathogenic Escherichia coli Employs Both Evasion and Resistance to Subvert Innate Immune-Mediated Zinc Toxicity for Dissemination. Proc. Natl. Acad. Sci. USA 2019, 116, 6341–6350. [Google Scholar] [CrossRef] [PubMed]
- Bolick, D.T.; Medeiros, P.H.Q.S.; Ledwaba, S.E.; Lima, A.A.M.; Nataro, J.P.; Barry, E.M.; Guerrant, R.L. Critical Role of Zinc in a New Murine Model of Enterotoxigenic Escherichia coli Diarrhea. Infect. Immun. 2018, 86, e00183-18. [Google Scholar] [CrossRef]
- Medeiros, P.; Bolick, D.T.; Roche, J.K.; Noronha, F.; Pinheiro, C.; Kolling, G.L.; Lima, A.; Guerrant, R.L. The Micronutrient Zinc Inhibits EAEC Strain 042 Adherence, Biofilm Formation, Virulence Gene Expression, and Epithelial Cytokine Responses Benefiting the Infected Host. Virulence 2013, 4, 624–633. [Google Scholar] [CrossRef]
- Bolick, D.T.; Kolling, G.L.; Moore, J.H.; de Oliveira, L.A.; Tung, K.; Philipson, C.; Viladomiu, M.; Hontecillas, R.; Bassaganya-Riera, J.; Guerrant, R.L. Zinc Deficiency Alters Host Response and Pathogen Virulence in a Mouse Model of Enteroaggregative Escherichia coli-Induced Diarrhea. Gut Microbes 2015, 5, 618–627. [Google Scholar] [CrossRef]
- Botella, H.; Peyron, P.; Levillain, F.; Poincloux, R.; Poquet, Y.; Brandli, I.; Wang, C.; Tailleux, L.; Tilleul, S.; Charrire, G.M.; et al. Mycobacterial P 1-Type ATPases Mediate Resistance to Zinc Poisoning in Human Macrophages. Cell Host Microbe 2011, 10, 248–259. [Google Scholar] [CrossRef]
- Ishida, T. Antibacterial Mechanism of Bacteriolyses of Bacterial Cell Walls by Zinc (II) Ion Induced Activations of PGN Autolysins and DNA Damages. J. Genes Proteins 2017, 1, 1–7. [Google Scholar]
- Sánchez-Uribe, P.; Romera-Recio, E.; Cabrera-Gómez, C.G.; Hernández-Rodríguez, E.V.; Lamrani, Á.; González-Guijarro, B.; de Pascual-Monreal, C.; Mendonça-Pascoal, L.; Martínez-Alarcón, L.; Ramis, G. Effect of β-Mannanase Addition during Whole Pigs Fattening on Production Yields and Intestinal Health. Animals 2022, 12, 3012. [Google Scholar] [CrossRef]
- Bravo de Laguna, F.; Cabrera, C.; González, A.B.; de Pascual, C.; Pallarés, F.J.; Chevaux, E.; Castex, M.; Saornil, D.; Lebreton, P.; Ramis, G. Effect of Feeding Saccharomyces Cerevisiae Boulardii CNCM I-1079 to Sows and Piglets on Piglets’ Immune Response after Vaccination against Actinobacillus Pleuropneumoniae. Animals 2022, 12, 2513. [Google Scholar] [CrossRef] [PubMed]
- Moue, M.; Tohno, M.; Shimazu, T.; Kido, T.; Aso, H.; Saito, T.; Kitazawa, H. Toll-like Receptor 4 and Cytokine Expression Involved in Functional Immune Response in an Originally Established Porcine Intestinal Epitheliocyte Cell Line. Biochim. Biophys. Acta Gen. Subj. 2008, 1780, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Royaee, A.R.; Husmann, R.J.; Dawson, H.D.; Calzada-Nova, G.; Schnitzlein, W.M.; Zuckermann, F.A.; Lunney, J.K. Deciphering the Involvement of Innate Immune Factors in the Development of the Host Response to PRRSV Vaccination. Vet. Immunol. Immunopathol. 2004, 102, 199–216. [Google Scholar] [CrossRef] [PubMed]
- Gabler, N.K.; Spurlock, M.E. Integrating the Immune System with the Regulation of Growth and Efficiency. J. Anim. Sci. 2008, 86, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.J.; Kovacs-Nolan, J.A.; Yang, C.; Archbold, T.; Fan, M.Z.; Mine, Y. L-Tryptophan Exhibits Therapeutic Function in a Porcine Model of Dextran Sodium Sulfate (DSS)-Induced Colitis. J. Nutr. Biochem. 2010, 21, 468–475. [Google Scholar] [CrossRef]
- Bracarense, A.P.F.L.; Lucioli, J.; Grenier, B.; Drociunas Pacheco, G.; Moll, W.D.; Schatzmayr, G.; Oswald, I.P. Chronic Ingestion of Deoxynivalenol and Fumonisin, Alone or in Interaction, Induces Morphological and Immunological Changes in the Intestine of Piglets. Br. J. Nutr. 2012, 107, 1776–1786. [Google Scholar] [CrossRef]
- Verpoest, S.; Cay, B.; Favoreel, H.; De Regge, N. Age-Dependent Differences in Pseudorabies Virus Neuropathogenesis and Associated Cytokine Expression. J. Virol. 2017, 91, e02058-16. [Google Scholar] [CrossRef]
- Borca, M.V.; Gudmundsdottir, I.; Fernández-Sainz, I.J.; Holinka, L.G.; Risatti, G.R. Patterns of Cellular Gene Expression in Swine Macrophages Infected with Highly Virulent Classical Swine Fever Virus Strain Brescia. Virus Res. 2008, 138, 89–96. [Google Scholar] [CrossRef]
- Skovgaard, K.; Mortensen, S.; Poulsen, K.T.; Angen, Ø.; Heegaard, P.M.H. Validation of Putative Reference Genes for QRT-PCR Normalization in Tissues and Blood from Pigs Infected with Actinobacillus Pleuropneumoniae. Vet. Immunol. Immunopathol. 2007, 118, 140–146. [Google Scholar] [CrossRef]
- Hermann, J.R.; Muñoz-Zanzi, C.A.; Roof, M.B.; Burkhart, K.; Zimmerman, J.J. Probability of Porcine Reproductive and Respiratory Syndrome (PRRS) Virus Infection as a Function of Exposure Route and Dose. Vet. Microbiol. 2005, 110, 7–16. [Google Scholar] [CrossRef]
- Zhu, C.; Lv, H.; Chen, Z.; Wang, L.; Wu, X.; Chen, Z.; Zhang, W.; Liang, R.; Jiang, Z. Dietary Zinc Oxide Modulates Antioxidant Capacity, Small Intestine Development, and Jejunal Gene Expression in Weaned Piglets. Biol. Trace Elem. Res. 2017, 175, 331–338. [Google Scholar] [CrossRef]
- Oropeza-Moe, M.; Grøntvedt, C.A.; Phythian, C.J.; Sørum, H.; Fauske, A.K.; Framstad, T. Zinc Oxide Enriched Peat Influence Escherichia coli Infection Related Diarrhea, Growth Rates, Serum and Tissue Zinc Levels in Norwegian Piglets around Weaning: Five Case Herd Trials. Porc. Health Manag. 2017, 3, 14. [Google Scholar] [CrossRef] [PubMed]
- Grilli, E.; Tugnoli, B.; Vitari, F.; Domeneghini, C.; Morlacchini, M.; Piva, A.; Prandini, A. Low Doses of Microencapsulated Zinc Oxide Improve Performance and Modulate the Ileum Architecture, Inflammatory Cytokines and Tight Junctions Expression of Weaned Pigs. Animal 2015, 9, 1760–1768. [Google Scholar] [CrossRef] [PubMed]
- Ramis, G.; Escuder, M.; Murillo, J.; Muñoz, A. Effect of Oral Vaccination against Escherichia coli Combined with a Nutritional Tool During. In Proceedings of the 26th International Pig Veterinary Society Congress, Rio de Janeiro, Brazil, 21–24 June 2022; Volume 1, p. 452. [Google Scholar]
- Vangroenweghe, F.A.C.J.; Boone, M. Vaccination with an Escherichia coli F4/F18 Vaccine Improves Piglet Performance Combined with a Reduction in Antimicrobial Use and Secondary Infections Due to Streptococcus Suis. Animals 2022, 12, 2231. [Google Scholar] [CrossRef] [PubMed]
- Bonetti, A.; Tugnoli, B.; Piva, A.; Grilli, E. Towards Zero Zinc Oxide: Feeding Strategies to Manage Post-Weaning Diarrhea in Piglets. Animals 2021, 11, 642. [Google Scholar] [CrossRef]
- Katouli, M.; Melin, L.; Jensen-Waern, M.; Wallgren, P.; Möllby, R. The Effect of Zinc Oxide Supplementation on the Stability of the Intestinal Flora with Special Reference to Composition of Coliforms in Weaned Pigs. J. Appl. Microbiol. 1999, 87, 564–573. [Google Scholar] [CrossRef]
- Mores, N.; Cristani, J.; Piffer, I.A.; Barioni, W., Jr.; Lima, G.M.M. Effects of Zinc Oxide on Postweaning Diarrhea Control in Pigs Experimentally Infected with E. coli. Arq. Bras. Med. Veterinária Zootec. 1998, 50, 513–523. [Google Scholar]
- Roselli, M.; Finamore, A.; Garaguso, I.; Britti, M.S.; Mengheri, E. Zinc Oxide Protects Cultured Enterocytes from the Damage Induced by Escherichia coli. J. Nutr. 2003, 133, 4077–4082. [Google Scholar] [CrossRef]
- Capaldo, C.T.; Nusrat, A. Cytokine Regulation of Tight Junctions. Biochim. Biophys. Acta Biomembr. 2010, 1788, 864–871. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, Y.; Zhang, D.; Bai, Y.; Liu, G.; Li, P.; Li, J.; Li, Y. Immunogenicity and Protective Efficacy of Enterotoxigenic Escherichia coli (ETEC) Total RNA against ETEC Challenge in a Mouse Model. Sci. Rep. 2020, 10, 20530. [Google Scholar] [CrossRef]
- Groschwitz, K.R.; Hogan, S.P. Intestinal Barrier Function: Molecular Regulation and Disease Pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef]
- Esteruelas, L.; Sánchez-Uribe, P.; Claver, M.; Benito, V.; Ramis, G. mRNA Quantification of Intestinal Integrity Biomarkers after the Administration of a Single-Dose Live Non-Pathogenic Escherichia coli Oral Vaccine. In Proceedings of the 26th International Pig Veterinary Society Congress, Rio de Janeiro, Brazil, 21–24 June 2022; Volume 1, p. 481. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rawstaff | Percentage |
|---|---|
| Corn 13% | 25.00 |
| Wheat F10 | 25.00 |
| Malting barley 9.5 | 15.72 |
| Soymeal 46% | 10.05 |
| Processed soy protein HP300 | 6.00 |
| Porcine protein hydrolysate (Palbio 50 RD) | 3.75 |
| Sheep sweet whey | 3.60 |
| Refatted whey 50 | 2.50 |
| Whey bran | 2.50 |
| Soy oil F10 | 2.36 |
| L-Lysine 50 | 0.95 |
| Calcium carbonate | 0.69 |
| Monocalcium phosphate | 0.62 |
| Vitamin-mineral corrector 1 | 0.30 |
| L-Threonine | 0.26 |
| DL-Methionine | 0.24 |
| Salt | 0.22 |
| L-Valine | 0.10 |
| L-Tryptophan | 0.08 |
| Mycotoxin absorber | 0.05 |
| Gene | Primer Forward (5′ → 3′) | Primer Reverse (5′ → 3′) | References |
|---|---|---|---|
| Occludin | 5′-TTGCTGTGAAAACTCGAAGC-3′ | 5′-CCACTCTCTCCGCATAGTCC-3′ | [12] |
| Zonulin 1 | 5′-CACAGATGCCACAGATGACAG-3′ | 5′-AGTGATAGCGAACCATGTGC-3′ | [12] |
| Claudin 1 | 5′-ACCCCAGTCAATGCCAGATA-3′ | 5′-GGCGAAGGTTTTGGATAGG-3′ | [21] |
| Calprotectin | 5′-AATTACCACGCCATCTACGC-3′ | 5′-TGATGTCCAGCTCTTTGAACC-3′ | [12] |
| IFN-α | 5′-CCCCTGTGCCTGGGAGAT-3′ | 5′-AGGTTTCTGGAGGAAGAGAAGGA-3′ | [23] |
| IFN-γ | 5-TGGTAGCTCTGGGAAACTGAATG-3′ | 5′-GGCTTTGCGCTGGATCTG-3′ | [24] |
| TNF-α | 5′-ACTCGGAACCTCATGGACAG-3′ | 5′-AGGGGTGAGTCAGTGTGACC-3′ | [25] |
| IL-12p35 | 5′-AGTTCCAGGCCATGAATGCA-3′ | 5′-TGGCACAGTCTCACTGTTGA-3′ | [23] |
| IL-12p40 | 5′-TTTCAGACCCGACGAACTCT-3′ | 5′-CATTGGGGTACCAGTCCAAC-3′ | [26] |
| IL-10 | 5′-TGAGAACAGCTGCATCCACTTC-3 | 5′-TCTGGTCCTTCGTTTGAAAGAAA-3′ | [24] |
| TGF-β | 5′-CACGTGGAGCTATACCAGAA-3′ | 5′-TCCGGTGACATCAAAGGACA-3′ | [23] |
| IL-8 | 5′-GCTCTCTGTGAGGCTGCAGTTC-3′ | 5′-AAGGTGTGGAATGCGTATTTATGC-3′ | [27] |
| IL-1α | 5′-GTGCTCAAAACGAAGACGAACC-3′ | 5′-CATATTGCCATGCTTTTCCCAGAA-3′ | [28] |
| IL-1β | 5′-AACGTGCAGTCTATGGAGT-3′ | 5′-GAACACCACTTCTCTCTTCA-3′ | [29] |
| IL-6 | 5′-CTGGCAGAAAACAACCTGAACC-3′ | 5′-TGATTCTCATCAAGCAGGTCTCC-3′ | [29] |
| β-actin | 5′-CTACGTCGCCCTGGACTTC-3′ | 5′-GATGCCGCAGGATTCCAT-3′ | [30] |
| Parameter | Neg | ZnO | V | V + ZnO | p-Value |
|---|---|---|---|---|---|
| ADG14 (Kgd−1) | 0.180 ± 0.006 a | 0.217 ± 0.013 b | 0.227 ± 0.006 b | 0.277 ± 0.007 c | <0.001 |
| ADG25 (Kgd−1) | 0.347 ± 0.054 a,b | 0.388 ± 0.040 a | 0.304 ± 0.007 b | 0.288 ± 0.014 c | 0.045 |
| Sampling dpv | Group | Neg | V | V + ZnO | p-Value |
|---|---|---|---|---|---|
| 6 | ECOTotal | 100% (10/10) | 100% (20/20) | 90% (18/20) | NS |
| F4 | 0% (0/10) AR = −3.7 | 70% (14/20) AR = 2.1 | 60% (12/20) | <0.001 | |
| H7 | 100% (10/10) | 90% (18/20) | 85% (17/20) | NS | |
| CRISPR | 0% (0/10) AR = −2.8 | 55% (11/20) AR = 2.0 | 40% (8/20) | 0.003 | |
| O8 | 70% (7/10) | 85% (17/20) | 70% (14/20) | NS | |
| 8 | ECOTotal | 100% (10/10) | 100% (20/20) | 100% (20/20) | NS |
| F4 | 0% (0/10) AR = −3.7 | 85% (17/20) AR = 2.1 | 85% (17/20) AR = 2.1 | <0.001 | |
| H7 | 100% (10/10) | 100% (20/20) | 85% (17/20) AR = −2.2 | 0.055 | |
| CRISPR | 0% (0/10) AR = −3.1 | 55% (11/20) | 75% (15/20) | 0.001 | |
| O8 | 40% (4/10) AR = −2.7 | 55% (11/20) | 90% (18/20) AR = 2.1 | 0.013 | |
| 11 | ECOTotal | 100% (10/10) | 80% (16/20) | 75% (15/20) | NS |
| F4 | 0% (0/10) | 55% (11/20) | 45% (9/20) | 0.013 | |
| H7 | 100% (10/10) | 65% (13/20) | 55% (11/20) | NS | |
| CRISPR | 0% (0/19) | 15% (3/20) | 15% (3/20) | NS | |
| O8 | 30% (4/10) | 65% (13/20) AR = 2.2 | 30% (6/20) | 0.027 | |
| 13 | ECOTotal | 100% (10/10) | 100% (20/20) | 90% (18/20) | NS |
| F4 | 0% (0/10) | 5% (1/20) | 15% (3/20) | NS | |
| H7 | 90% (9/10) | 90% (18/20) | 70% (14/20) | NS | |
| CRISPR | 0% (0/10) | 35% (7/10) AR = 2.2 | 15% (3/20) | 0.027 | |
| O8 | 100% (10/10) AR = 4.3 | 30% (6/20) | 15% (3/20) AR = −2.7 | <0.001 | |
| 15 | ECOTotal | 100% (10/10) | 100% (20/20) | 65% (17/20) | 0.055 |
| F4 | 0% (0/10) | 5% (1/20) | 5% (1/20) | NS | |
| H7 | 100% (10/10) AR = 2.9 | 60% (12/20) | 40% (8/20) AR = −2.4 | 0.007 | |
| CRISPR | 0% (0/10) | 10% (2/20) | 10% (2/20) | NS | |
| O8 | 80% (8/10) AR = 2.7 | 65% (13/20) AR = 2.7% | 0% (0/20) AR = −4.9 | <0.001 |
| F4 | H7 | CRISPR | O8 | |
|---|---|---|---|---|
| ECOTotal | 0.502 | 0.717 | 0.531 | 0.231 |
| F4 | 0.809 | 0.549 | 0.574 | |
| H7 | 0.576 | 0.515 | ||
| CRISPR | 0.325 |
| ECOTotal | F4 | H7 | CRISPR | O8 | |
|---|---|---|---|---|---|
| CALP | 0.068 | 0.210 | 0.315 | −0.081 | 0.306 |
| IFNα | −0.305 | −0.110 | −0.202 | 0.589 | −0.049 |
| IFNγ | −0.018 | 0.063 | 0.278 | −0.067 | 0.345 |
| IL1α | −0.189 | 0.007 | −0.003 | −0.101 | 0.289 |
| IL1β | −0.280 | −0.184 | −0.225 | 0.592 | −0.092 |
| IL6 | 0.237 | 0.092 | 0.332 | −0.039 | 0.092 |
| IL8 | −0.262 | 0.000 | −0.080 | −0.164 | 0.244 |
| IL10 | −0.229 | −0.195 | −0.277 | −0.433 | −0.177 |
| IL12p35 | −0.062 | 0.062 | 0.202 | −0.105 | 0.202 |
| IL12p40 | −0.039 | 0.071 | 0.229 | −0.134 | 0.208 |
| TGFβ | −0.118 | 0.008 | 0.133 | −0.115 | 0.319 |
| TNFα | 0.035 | 0.019 | 0.196 | −0.144 | 0.140 |
| ECOTotal | F4 | H7 | CRISPR | O8 | |
|---|---|---|---|---|---|
| OCLD | −0.174 | −0.115 | 0.022 | −0.158 | 0.265 |
| ZON | −0.259 | 0.069 | 0.084 | −0.142 | 0.587 |
| CLAU | −0.241 | −0.294 | −0.103 | 0.374 | −0.129 |
| Day | Original | Predicted Membership Group (%) | ||
|---|---|---|---|---|
| Neg | V | V + ZnO | ||
| 6 | Neg | 70.0 | 10.0 | 20.0 |
| V | 10.0 | 75.0 | 15.0 | |
| V + ZnO | 5.0 | 40.0 | 55.0 | |
| 8 | Neg | 90.0 | 10.0 | 0.0 |
| V | 0.0 | 80.0 | 20.0 | |
| V + ZnO | 0.0 | 10.0 | 90.0 | |
| 11 | Neg | 40.0 | 0.0 | 60.0 |
| V | 0.0 | 80.0 | 20.0 | |
| V + ZnO | 0.0 | 20.0 | 80.0 | |
| 13 | Neg | 80.0 | 10.0 | 10.0 |
| V | 5.0 | 85.0 | 10.0 | |
| V + ZnO | 0.0 | 15.8 | 84.2 | |
| 15 | Neg | 100.0 | 0.0 | 0.0 |
| V | 0.0 | 90.0 | 10.0 | |
| V + ZnO | 0.0 | 31.6 | 68.4 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramis, G.; Murciano, F.; Orengo, J.; González-Guijarro, B.; Cuellar-Flores, A.; Serrano, D.; Muñoz Luna, A.; Sánchez-Uribe, P.; Martínez-Alarcón, L. Is Oral Vaccination against Escherichia coli Influenced by Zinc Oxide? Animals 2023, 13, 1754. https://doi.org/10.3390/ani13111754
Ramis G, Murciano F, Orengo J, González-Guijarro B, Cuellar-Flores A, Serrano D, Muñoz Luna A, Sánchez-Uribe P, Martínez-Alarcón L. Is Oral Vaccination against Escherichia coli Influenced by Zinc Oxide? Animals. 2023; 13(11):1754. https://doi.org/10.3390/ani13111754
Chicago/Turabian StyleRamis, Guillermo, Francisco Murciano, Juan Orengo, Belén González-Guijarro, Amanda Cuellar-Flores, Daniel Serrano, Antonio Muñoz Luna, Pedro Sánchez-Uribe, and Laura Martínez-Alarcón. 2023. "Is Oral Vaccination against Escherichia coli Influenced by Zinc Oxide?" Animals 13, no. 11: 1754. https://doi.org/10.3390/ani13111754