
Evaluation of Fecundity, Fertilization, Hatching, and Gonadosomatic Index of Exotic Clarias gariepinus (Burchell, 1822) and Native Clarias macromystax (Gunther, 1864) under Semi-Arid Conditions of Nigeria
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Collection Area
2.3. Brood Stock Selection Inducement
2.4. Determination of Fecundity
2.5. Total Egg Number Release
2.6. Determination of Gonadosomatic Index
2.7. Estimation of Fertilization and Hatching Rate
- (i)
- Fertilization rate (%) = Number of fertilized eggs/Number of estimated eggs × 100;
- (ii)
- Hatchability (%) = × 100;
- (iii)
- Survival (%) = × 100%.
2.8. Determination of Water Quality
2.9. Statistical Analysis
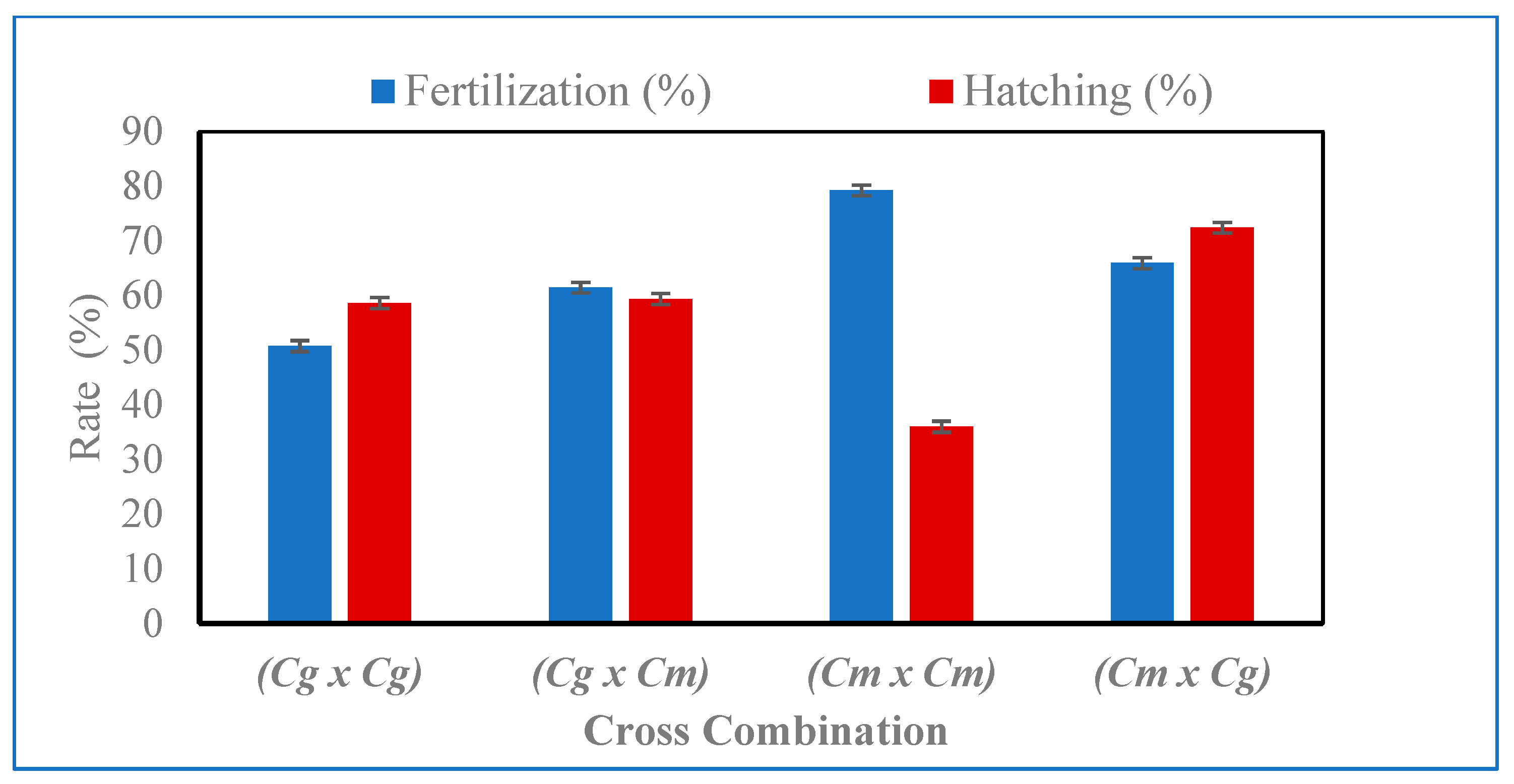
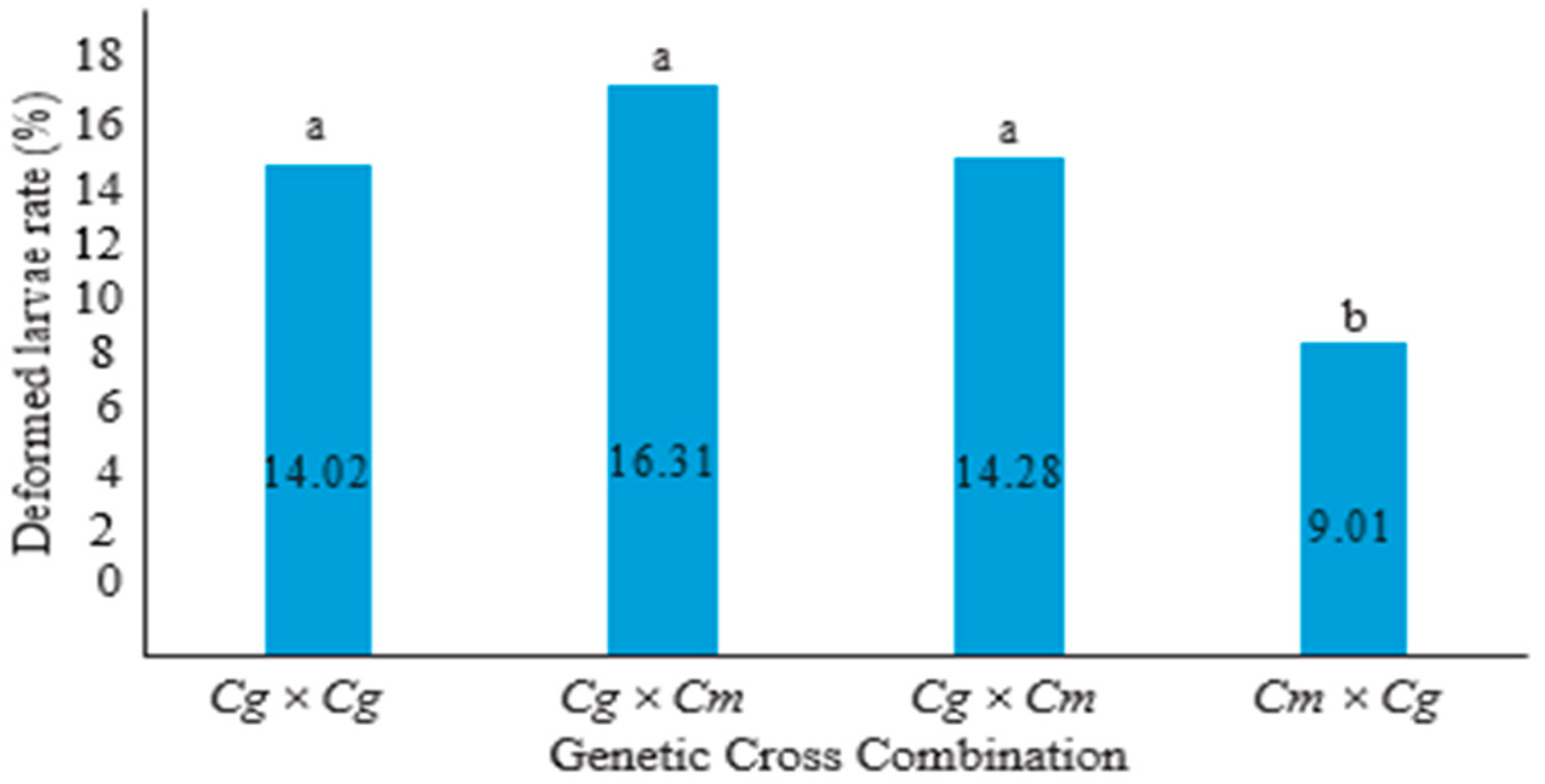
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adebayo, O.T. Reproductive performance of African Clariid Catfish Clarias gariepinus broodstock on varying maternal stress. J. Fish. Int. 2006, 1, 17–20. [Google Scholar]
- Adah, P.; Onyia, L.U.; Obande, R.A. Fish Hybridization in Some Catfishes: A Review. Biotechnology 2014, 13, 248–251. [Google Scholar] [CrossRef]
- Aminur Rah, M.; Arshad, A.B.; Marimuthu, K.; Ara, R.; Amin, S.M.N. Inter-specific Hybridization and Its Potential for Aquaculture of Fin Fishes. Asian J. Anim. Vet. Adv. 2013, 8, 139–153. [Google Scholar] [CrossRef]
- Akinwande, A.A.; Moody, F.O.; Sogbesan, O.A.; Ugwumba, A.A.A.; Ovie, S.O. A study of African mud catfish Clarias gariepinus. J. Arid. Zone Fish. 2005, 1, 21–22. [Google Scholar]
- Adewolu, M.A.; Ogunsanmi, A.O.; Yunusa, A. Studies on Growth Performance and Feed utilization of two clariid catfish and their hybrid reared under different culture systems. Eur. J. Sci. Res. 2008, 23, 252–260. [Google Scholar]
- Omeji, S.; Obande, R.A.; Oyaje, J. Intra-specific hybridization of local and exotic Clarias gariepinus. IJMBR 2013, 1, 35–41. [Google Scholar]
- Antechoir, M.A. Catfish Aquaculture Industry Assessment in Nigeria. African Union; Inter African Bureau for Animal Resources: Lagos, Nigeria, 2013; pp. 235–247. [Google Scholar]
- Aluko, P.O.; Shaba, M. Intra and Interspecific Hybridization Studies between exotic Clarias gariepinus and two indigenous Clariid species. Niger. J. Genet. 1999, 14, 56. [Google Scholar]
- Rahman, M.A.; Bhadra, A.; Begum, N.; Islam, M.S.; Hussain, M.G. Production of hybrid vigor through cross breeding between Clarias batrachus Lin. and Clarias gariepinus Bur. Aquaculture 1995, 138, 125–130. [Google Scholar] [CrossRef]
- Akinwande, A.A.; Fagbenro, O.A.; Adebayo, O.T. Growth and heterosis in reciprocal Clarias hybrids between Clarias gariepinus and Clarias angularis. J. Fish. Int. 2011, 6, 67–70. [Google Scholar]
- Owodeinde, F.G.; Fakoya, K.A.; Anetekhai, M.A. Growth performance of hybrid Catfish (Clarias gariepinus × Heterobranchus bidorsalis) in earthen ponds. Asian J. Biol. Sci. 2012, 5, 192–200. [Google Scholar] [CrossRef]
- Um-E-Kalsoom; Salim, M.; Shahzadi, T.; Barlas, A. Growth performance and feed conversion ratio (FCR) in hybrid fish (Catla catla × Labeo rohita) fed on wheat bran, rice broken and blood meal. Park. Vet. J. 2009, 29, 55–58. [Google Scholar]
- Dunham, R.A.; Smitherman, R.O.; Goodman, R.K. Comparison of mass selection, cross breeding, and hybridization for improving growth of channel catfish. Progress. Fish-Cult. 1987, 49, 293–296. [Google Scholar] [CrossRef]
- Ndimele, P.E.; Owodeinde, F.G. Comparative Reproductive and Growth Performance of Clarias gariepinus (Burchell, 1822) and Its Hybrid Induced with Synthetic Hormone and Pituitary Gland of Clarias gariepinus. Turk. J. Fish. Aquat. Sci. 2012, 12, 619–626. [Google Scholar] [CrossRef]
- Richter, C.J.J.; Edarg, E.H.; Verreth, A.J. African catfish (Clarias gariepinus). In Brood Stock Management and Egg and Larval Quality; Bromage, N.R., Roberts, R.J., Eds.; Blackwell Scientific Publication: Oxford, UK, 1995; pp. 242–276. [Google Scholar]
- Srivastava, P.P.; Raizada, S.; Dayal, R.; Chowdhary, S.; Lakra, W.S.; Akhilesh, K.Y.; Sharma, P.; Gupta, J. Breeding and Larval Rearing of Asian Catfish, Clarias batrachus (Linnaeus, 1758) on Live and Artificial Feed. J. Aquac. Res. Dev. 2012, 3, 4. [Google Scholar] [CrossRef]
- Argungu, L.A.; Christianus, M.N.; Amin, S.K.; Daud, I.S.; Siraj, S.; Rahman, M. Asian Catfish Clarias batrachus (Linnaeus, 1758) Getting Critically Endangered. Asian J. Anim. Vet. Adv. 2012, 8, 1683–1699. [Google Scholar] [CrossRef]
- Mohapatra, B.K. Conservation of the Asiatic catfish, Clarias batrachus through artificial propagation and larval rearing technique in India. Aquaculture 2004, 9, 8–9. [Google Scholar]
- de Graaf, G.J.; Galemoni, F.; Banzoussi, B. The Artificial Reproduction and Fingerling Production of the African Catfish, Clarias gariepinus (Burchell, 1822) in protected and unprotected ponds. Aquac. Res. 1995, 26, 233–242. [Google Scholar] [CrossRef]
- Yadav, A.K.; Mishra, R.K.; Singh, S.K.; Varshney, P.K.; Pandey, A.K.; Lakra, W.S. Induced spawning of Asian catfish, Clarias batrachus, with different doses of GnRH-based drugs. J. Exp. Zool. 2011, 14, 199–202. [Google Scholar]
- Ataguba, G.; Annune, A.P.; Ogbe, G.F. Growth performance of two African catfishes Clarias gariepinus and Heterobranchus longifilis and their hybrids in plastic aquaria. Livest. Res. Rural Dev. 2010, 22, 30. [Google Scholar]
- Hulata, G. Genetic manipulation in aquaculture: A review of stock improvement by classical and modern technologies. Genetica 2001, 111, 155–173. [Google Scholar] [CrossRef]
- Jothilakshmanan, N.; Karal Marx, K. Hybridization between Indian catfish, female Hetropneustes fossilis (Bloch) and Asian catfish, male Clarias batrachus (Linn). Afr. J. Biotechnol. 2013, 12, 976–981. [Google Scholar] [CrossRef]
- Ndimele, P.E.; Owodeinde, F.G.; Kumolu-Johnson, C.A.; Jimoh, A.A.; Whenu, O.O.; Onyenania, O.B. Growth Performance of the Reciprocal Hybrids of Clarias gariepinus (Burchell, 1822) and Heterobranchus bidorsalis (Valenciennes, 1840). J. Biol. Sci. 2011, 3, 137–140. [Google Scholar]
- Oguguah, N.M.; Nwadukwe, F.; Atama, C.I.; Chdobem, J.I.; Eyo, J.E. Growth performance of hybrid catfish (Hetrobranchus bidorsalis (♀) x Clarias gariepinus (♂) at various stocking densities in varied culture tanks. Anim. Res. Int. 2011, 8, 1419–1430. [Google Scholar]
- Ataguba, G.A.; Annune, P.A.; Ogbe, F.G. Induced breeding and early growth of progeny from crosses between two African clariid fishes, Clarias gariepinus (Burchell) and Heterobranchus longifilis under hatchery conditions. J. Appl. Biosci. 2009, 14, 755–760. [Google Scholar]
- Hossain, Q.M.; Altaf Hossain, A.H.; Parween, S. Artificial breeding and nursery practices of Clarias batrachus (Linnaeus, 1758). Sci. World 2006, 4, 4. [Google Scholar]
- Morni, M.M. Study on Cross Breeding between Asian and African Catfish (Clarias macrocephalus and Clarias gariepinus) and Some Aspects of The Hybrid Larvae Development and Rearing. Master’s Thesis, Kolej University of Science and Technology Malaysia, Petaling Jaya, Malaysia, 2003; p. 137. [Google Scholar]
- Madu, C.T.; Okoye, F.C.; Sado, E.K.; Omoruikoba, V.S.; Butkole, N.O.; Millita, E.O. A Preliminary Report on Induced Breeding Trials with the Mudfish Clarias angutllaris; Kl. Rl Annual Report; KLRI: Korea, Republic of Korea, 1994; pp. 144–145. [Google Scholar]
- Sahoo, S.K.; Giri, S.S.; Chandra, S.; Sahu, A.K. Spawning performance and egg quality of Asian catfish Clarias batrachus (Linn.) at various doses of Human Chorionic Gonadotropin (HCG) injection and latency periods during spawning induction. Aquaculture 2007, 266, 289–292. [Google Scholar] [CrossRef]
- Food Agricultural Organisation. Cultured Aquatic Species Information Programme. Clarias gariepinus; Pouomogne, V., Ed.; FAO Fisheries and Aquaculture: Rome, Italy, 2007. [Google Scholar]
- Ariola, C.N.; Okpokwasili, G.C. The effects of indigenous probiotics on egg hatchability and larval viability of Clarias gariepinus. Interdiscip. J. Appl. Sci. 2012, 7, 1. [Google Scholar] [CrossRef]
- Oyeleye, O.; Omitogun, O.G. Evaluation of motility of the short term cryopreserved sperm of African giant catfish-Clarias gariepinus. Ife J. Agric. 2007, 22, 11–16. [Google Scholar]



{kind=link}
{kind=link}
| Parameter | C. gariepinus | C. macromystax |
|---|---|---|
| Female weight (kg) | 1.13 a | 0.85 b |
| Number of eggs (1/g−1) | 700 a | 845 b |
| Fecundity (eggs/kg−1 fish) | 145,715.17 ± 1283.51 b | 93,672.50 ± 477.92 a |
| Strains | Weight (g) | Total Length (cm) | Weight of Ovary (g) | Number of Eggs | GSI (%) | PGSI |
|---|---|---|---|---|---|---|
| C. gariepinus | 1310.0 a | 51.0 a | 192.8 a | 198,105.0 a | 12.92 b | 17.41 b |
| C. macromystax | 651.0 b | 43.0 a | 132.1 b | 163,054.0 b | 16.89 a | 25.51 a |
| Standard Error of the Mean | 0.57 * | 0.57 ns | 0.58 * | 0.57 * | 0.06 * | 0.06 * |
| Strains | Weight (g) | Length (cm) | Testes Weight (g) | Length of Right Testes (cm) | Length of Left Testes (cm) | Milt Volume (mL) | Motility Duration (s) |
|---|---|---|---|---|---|---|---|
| C. gariepinus | 1500.0 a | 56.40 a | 8.90 a | 4.74 a | 6.55 a | 9.50 a | 63.00 a |
| C. macromystax | 550.0 b | 49.75 b | 6.70 b | 3.80 b | 4.55 b | 7.50 a | 55.00 b |
| Standard Error of the Mean | 28.86 * | 0.05 * | 0.58 * | 0.01 * | 0.58 * | 0.57 ns | 0.57 * |
| Parameter | C. macromystax | C. gariepinus | p-Value |
|---|---|---|---|
| Weight of the right whole testis (g) | 13.21 ± 4.61 | 2.64 ± 1.33 * | 0.04 |
| Weight right testis | 4.81 ± 1.73 | 1.52 ± 0.53 | 0.08 |
| Volume right testis semen (mL) | 8.41 ± 3.08 | 1.12 ± 0.80 * | 0.03 |
| Weight of the left whole testis (g) | 13.40 ± 2.55 | 1.45 ± 0.32 * | 0.00 |
| Weight of the left testis (g) | 4.82 ± 1.30 | 1.09 ± 0.17 * | 0.01 |
| Volume left testis semen (mL) | 8.58 ± 1.92 | 0.37 ± 0.18 * | 0.00 |
| Total semen volume (mL) | 16.69 ± 5.58 | 2.49 ± 0.98 * | 0.01 |
| Total testis weight (g) | 10.25 ± 3.36 | 2.60 ± 0.67 * | 0.03 |
| Liveweight (kg) | 2.54 ± 0.12 | 2.11 ± 0.15 * | 0.04 |
| Total gonadal weight (g) | 27.94 ± 8.17 | 4.09 ± 1.63 * | 0.01 |
| Gonadosomatic Index | 1.15 ± 0.38 | 0.19 ± 0.06 * | 0.02 |
| Condition factor | 0.84 ± 0.07 | 0.66 ± 0.06 | 0.01 |
| Parameter | C. macromystax (n = 20) | C. gariepinus (n = 20) | p-Value |
|---|---|---|---|
| Live weight (kg) | 2.07 ± 0.11 | 3.66 ± 1.18 | 0.08 |
| Weight of eggs (g) | 166.67 ± 31.69 | 233.33 ± 55.48 | 0.30 |
| Egg population (×103 eggs/female) | 77.28 ± 22.52 | 94.08 ± 44.63 | 0.72 |
| Gonadosomatic Index | 8.24 ± 1.62 | 7.04 ± 1.64 | 0.66 |
| Condition factor | 0.85 ± 0.13 | 0.85 ± 0.08 | 0.32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esa, Y.B.; Dadile, A.M.; Syukri, F.; Christianus, A.; Diyaware, M.Y. Evaluation of Fecundity, Fertilization, Hatching, and Gonadosomatic Index of Exotic Clarias gariepinus (Burchell, 1822) and Native Clarias macromystax (Gunther, 1864) under Semi-Arid Conditions of Nigeria. Animals 2023, 13, 1723. https://doi.org/10.3390/ani13111723
Esa YB, Dadile AM, Syukri F, Christianus A, Diyaware MY. Evaluation of Fecundity, Fertilization, Hatching, and Gonadosomatic Index of Exotic Clarias gariepinus (Burchell, 1822) and Native Clarias macromystax (Gunther, 1864) under Semi-Arid Conditions of Nigeria. Animals. 2023; 13(11):1723. https://doi.org/10.3390/ani13111723
Chicago/Turabian StyleEsa, Yuzine B., Abdulrahman Muhammad Dadile, Fadhil Syukri, Annie Christianus, and Mohammad Y. Diyaware. 2023. "Evaluation of Fecundity, Fertilization, Hatching, and Gonadosomatic Index of Exotic Clarias gariepinus (Burchell, 1822) and Native Clarias macromystax (Gunther, 1864) under Semi-Arid Conditions of Nigeria" Animals 13, no. 11: 1723. https://doi.org/10.3390/ani13111723