
Radio Tracking Reveals the Home Range and Activity Patterns of Nutria (Myocastor coypus) in the Macdo Wetland in South Korea



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Capturing and Handling
2.3. Data Analysis
Home Range Estimation
2.4. Activity Patterns
3. Results
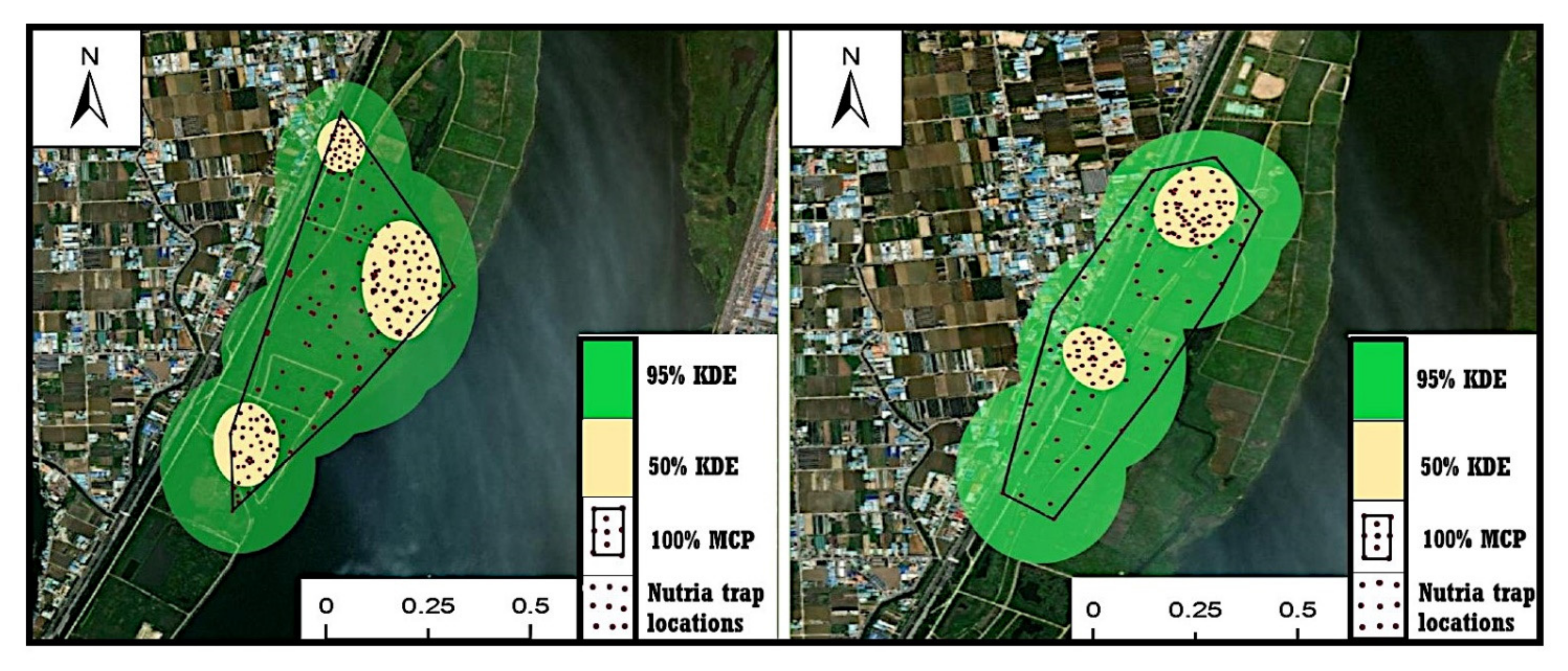
3.1. Home Range of Nutria
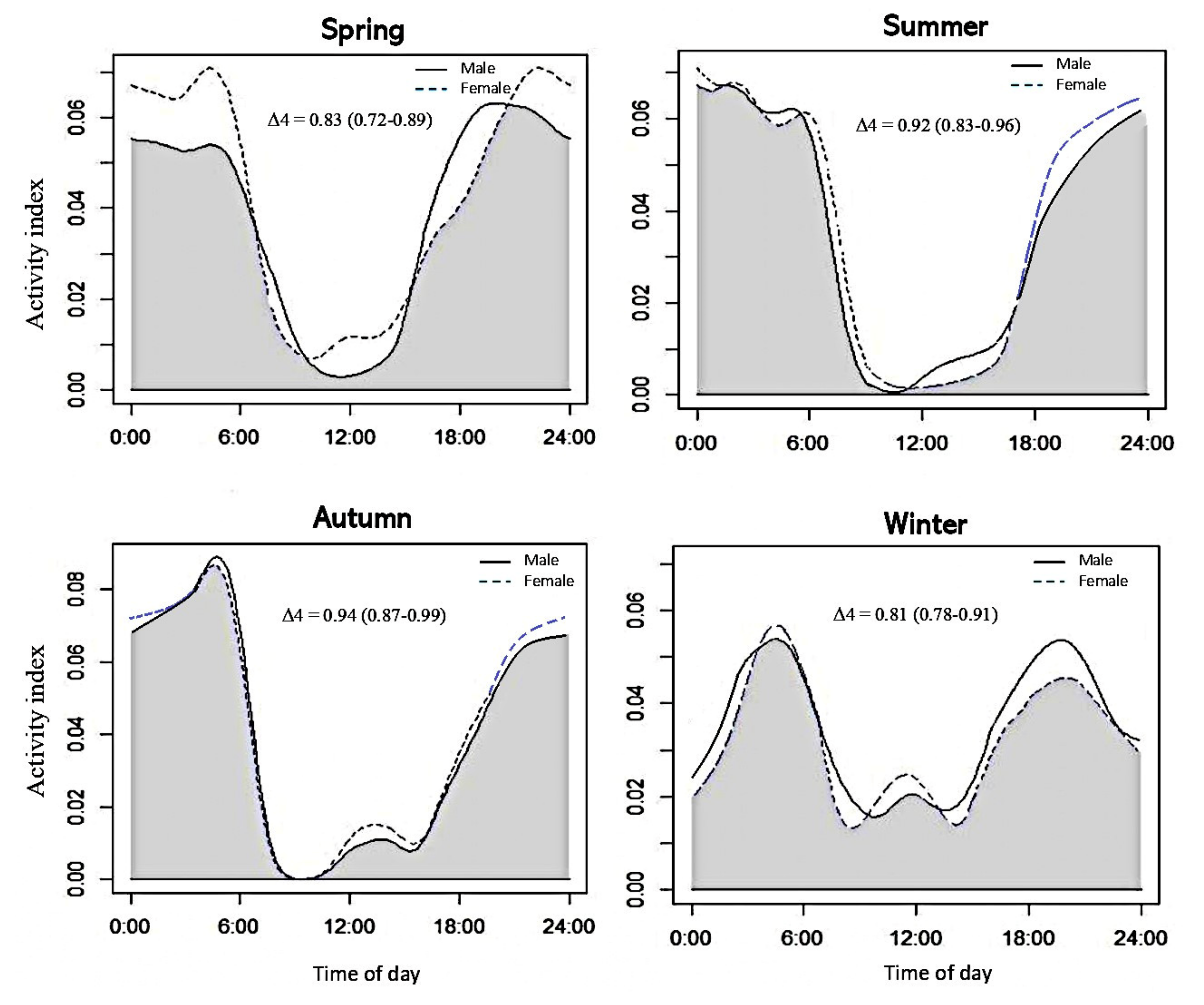
3.2. Activity Patterns of Nutria
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien species as a driver of recent extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef] [PubMed]
- Mazza, G.; Tricarico, E.; Genovesi, P.; Gherardi, F. Biological invaders are threats to human health: An overview. Ethol. Ecol. Evol. 2014, 26, 112–129. [Google Scholar] [CrossRef]
- Carter, J.; Leonard, B.P. A review of the literature on the worldwide distribution, spread of, and efforts to eradicate the coypu (Myocastor coypus). Wildl. Soc. Bull. 2002, 30, 162–175. [Google Scholar]
- Kim, I.R.; Choi, W.; Kim, A.; Lim, J.; Lee, D.H.; Lee, J.R. Genetic diversity and population structure of nutria (Myocastor coypus) in South Korea. Animals 2019, 9, 1164. [Google Scholar] [CrossRef]
- Ojeda, R.; Bindau, C.; Emmons, L. Myocastor Coypus (errata version published in 2017); The IUCN Red List of Threatened Species: Cambridge, UK, 2016; E.T14085A12173. [Google Scholar]
- Kim, Y.-C.; Kim, A.; Lim, J.; Kim, T.S.; Park, S.G.; Kim, M.; Lee, J.H.; Lee, J.R.; Lee, D.H. Distribution and management of nutria (Myocastor coypus) populations in South Korea. Sustainability 2019, 11, 4169. [Google Scholar] [CrossRef]
- Banjade, M.; Han, S.H.; Jeong, Y.H.; Oh, H.S. Diel and seasonal activity pattern of alien sika deer with sympatric mammalian species form Muljangori-oreum wetland of Hallasan National Park, South Korea. J. Ecol. Environ. 2021, 45, 10. [Google Scholar] [CrossRef]
- Hong, S.; Do, Y.; Kim, J.Y.; Kim, D.K.; Joo, G.J. Distribution, spread and habitat preferences of nutria (Myocastor coypus) invading the lower Nakdong River, South Korea. Biol. Invasions 2015, 17, 1485–1496. [Google Scholar] [CrossRef]
- Adhikari, P.; Kim, B.J.; Hong, S.H.; Lee, D.H. Climate change induced habitat expansion of nutria (Myocastor coypus) in South Korea. Sci. Rep. 2022, 12, 3300. [Google Scholar] [CrossRef]
- Fratini, F.; Turchi, B.; Ebani, V.V.; Bertelloni, F.; Galiero, A.; Cerri, D. The presence of leptospira in coypus (Myocastor coypus) and rats (Rattus norvegicus) living in a protected wetland in Tuscany (Italy). Vet. Arh. 2015, 85, 407–414. [Google Scholar]
- Park, J.H.; Novilla, M.N.; Song, J.; Kim, K.S.; Chang, S.N.; Han, J.H.; Lee, B.H.; Lee, D.H.; Kim, H.M.; Kim, Y.H.; et al. The first case of Capillaria Hepatica infection in a nutria (Myocastor coypus) in Korea. Korean J. Parasitol. 2014, 52, 527–529. [Google Scholar] [CrossRef] [PubMed]
- Guichon, M.L.; Doncaster, C.P.; Cassini, M.H. Population structure of coypus (Myocastor coypus) in their region of origin and comparision with introduced populations. J. Zool. 2003, 261, 265–272. [Google Scholar] [CrossRef]
- Bertolino, S.; Perrone, A.; Gola, L. Effectiveness of coypu control in small Italian wetland areas. Wildl. Soc. Bull. 2005, 33, 714–720. [Google Scholar] [CrossRef]
- Schertler, A.; Rabitsch, W.; Moser, D.; Wessely, J.; Essl, F. The potential current distribution of the coypu (Myocastor coypus) in Europe and climate change induced shifts in the near future. NeoBiota 2020, 58, 129–160. [Google Scholar] [CrossRef]
- Harris, S.; Cresswell, W.J.; Forde, P.G.; Trewella, W.J.; Woollard, T.; Wray, S. Home-range analysis using radio-tracking data–a review of problems and techniques particularly as applied to the study of mammals. Mammal Rev. 1990, 20, 97–123. [Google Scholar] [CrossRef]
- Girard, I.; Dussalt, C.; Ouellet, J.-P.; Courtis, R.; Caron, A. Balancing number of locations with number of individuals in telemetry studies. J. Wildl. Manag. 2006, 70, 1249–1256. [Google Scholar] [CrossRef]
- Do, Y.; Kim, J.Y.; Im, R.; Kim, S.; Choi, J.; Joo, G. Spatial distribution and social characteristics for wetlands in Gyeongsangnam-Do Province. Korean J. Limnol. 2012, 45, 252–260. [Google Scholar]
- Lee, D.-H.; Kil, J.-H.; Kim, D.-E. The study on the distribution and inhabiting status of nutria (Myocastor coypus) in Korea. Korean J. Environ. Ecol. 2013, 27, 316–326. [Google Scholar] [CrossRef]
- Gannon, W.L.; Sikes, R.S. Guidelines of the American Society of mammalogist for the use of wild animals in Research. J. Mammal. 2007, 88, 809–823. [Google Scholar] [CrossRef]
- Schoener, T.W. An Empirically based estimate of home range. Theor. Popul. Biol. 1981, 20, 281–325. [Google Scholar] [CrossRef]
- Borger, L.; Franconi, N.; De Michele, G.; Gantz, A.; Meschi, F.; Manica, A.; Lovari, S.; Coulson, T. Effects of sampling regime on the mean and variance of home range size estimates. J. Anim. Ecol. 2006, 75, 1393–1405. [Google Scholar] [CrossRef]
- Brian, J.K.; Roberta, G.; Millspaugh, J. Analysis of animal space use and movements. In Radio Tracking and Animal Populations; Millspaugh, J.J., Marzluff, J.M., Eds.; Academic Press: Cambridge, MA, USA, 2001; pp. 125–166. [Google Scholar] [CrossRef]
- Rodgers, A.; Carr, A.; Beyer, H.; Smith, L.; Kie, J. HRT: Home Range Tools for ArcGIS; Version 2.04; Ontario Ministry of Natural Resources, Centre for Northern Forest Ecosystem Research: Wawa, ON, Canada, 2007. [Google Scholar]
- Worton, A.B.J. Kernel methods for estimating the utilization distribution in home-range studies. Ecology 1989, 70, 164–168. [Google Scholar] [CrossRef]
- Seaman, D.E.; Griffith, B.; Powell, R.A. KERNELHR: A Program for estimating animal home ranges. Wildl. Soc. Bull. 1998, 26, 95–100. [Google Scholar]
- Chynoweth, M.W.; Lepczyk, C.A.; Litton, C.M.; Hess, S.C.; Kellner, J.R.; Cordell, S. Home range use and movement patterns of non-native feral goats in a tropical island montane dry landscape. PLoS ONE 2015, 10, e0119231. [Google Scholar] [CrossRef] [PubMed]
- Bennie, J.J.; Duffy, J.P.; Inger, R.; Gaston, K.J. Biogeography of time partitioning in mammals. Proc. Natl. Acad. Sci. USA 2014, 111, 13727–13732. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.J. Control and eradication of invasive mammals in Great Britain. Rev. Sci. Tech. 2010, 29, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Willner, G.R.; Chapman, J.A.; Pursley, D. Reproduction, physiological responses, food habits, and abundance of nutria on Maryland marshes. Wildl. Monogr. 1979, 65, 3–43. [Google Scholar]
- Clements, L.E.N. Nutria survivorship, movement patterns, and home ranges. Southeast. Nat. 2009, 8, 399–410. [Google Scholar] [CrossRef]
- Kim, A.; Kim, Y.-C.; Lee, D.-H. Home range and daily activity of nutria (Myocastor coypus) using radio tracking in South Korea. J. Environ. Impact Assess 2020, 29, 182–197. [Google Scholar]
- Smith, L.M.; Cherry, R.P. Movement, seasonal activity, and home range of an isolated population of Glyptemys muhlenbergii, bog turtle, in the southern appalachians. Southeast. Nat. 2016, 15, 207–219. [Google Scholar] [CrossRef]
- Denena, M.M. Home Range and Movement of Nutria (Myocastor coypus) at Spring Lake in Central Texas, with Anecdotal Comments on the American Beaver (Castor canadensis) of the Same Area. Master’s Thesis, Southwest Texas State University, San Marcos, TX, USA, 2002. [Google Scholar]
- Cutrera, A.P.; Antinuchi, C.D.; Mora, M.S.; Vassallo, A.I. Home-range and activity patterns of the South American subterranean rodent Ctenomys talarum. J. Mammal. 2006, 87, 1183–1191. [Google Scholar] [CrossRef]
- Whisson, D.A.; Quinn, J.H.; Collins, K.C. Home range and movements of roof rats (Rattus rattus) in an old-growth riparian forest, California. J. Mammal. 2007, 88, 589–594. [Google Scholar] [CrossRef]
- Nevo, E. Mosaic Evolution of Subterranean Mammals: Tinkering, Regression, Progression, and Global Convergence; Oxford University Press: New York, NY, USA, 2007. [Google Scholar]
- Glogowski, R.; Panas, M. Efficiency and proximate composition of meat in male and female nutria (Myocastor coypus) in an extensive feeding system. Meat Sci. 2009, 81, 752–754. [Google Scholar] [CrossRef] [PubMed]
- Reggiani, G.; Boitani, L.; Antoni, S.; Stefano, R. De biology and control of the coypu in the mediterranean area. Suppl. Alle Ric. Di Biol. Della Selvag. 1993, 273, 67–100. [Google Scholar]
- Fisher, D.O.; Owens, I.P.F. Female home range size and the evolution of social organization in macropod marsupials. J. Anim. Ecol. 2000, 69, 1083–1098. [Google Scholar] [CrossRef]
- Clutton-brock, T.H.; Guinness, F.E.; Albon, S.D. Red Deer : Behavior and Ecology of Two Sexes; Edinburgh University: Edinburgh, UK, 1982. [Google Scholar]
- Komers, P.E.; Brotherton, P.N.M. Female space use is the best predictor of monogamy in mammals. Proc. R. Soc. B Biol. Sci. 1997, 264, 1261–1270. [Google Scholar] [CrossRef]
- Adler, G.H. Spacing patterns and social mating systems of echimyid rodents. J. Mammal. 2011, 92, 31–38. [Google Scholar] [CrossRef]
- Waterman, J.; Wolff, J.; Sherman, P. Male mating strategy in rodents. In Rodent Societies: An Ecological and Evolutionary Perspective; University of Chicago Press: Chicago, IL, USA, 2007. [Google Scholar]
- Finn, K.T.; Janse van Vuuren, A.K.; Hart, D.W.; Süess, T.; Zöttl, M.; Bennett, N.C. Seasonal changes in locomotor activity patterns of wild social natal Mole-Rats (Cryptomys hottentotus natalensis). Front. Ecol. Evol. 2022, 10, 62. [Google Scholar] [CrossRef]
- Masaki, M.; Koshimoto, C.; Tsuchiya, K.; Nishiwaki, A.; Morita, T. Body temperature profiles of the Korean field mouse Apodemus peninsulae during winter aggregation. Mammal Study 2005, 30, 33–40. [Google Scholar] [CrossRef]
- Halle, S.; Stenseth, N. Activity Patterns in Small Mammals: An Ecological Approach; Springer: New York, NY, USA, 2000. [Google Scholar]
- Lee, E.; Rhim, S. Seasonal home ranges and activity of three rodent species in a post-fire planted stand. Folia Zool. 2016, 65, 101–106. [Google Scholar] [CrossRef]
- Salas, M.; Hernandez-Hernandez, J.C.; Iglesias, M.; Corriale, M.J. Activity patterns and behavior of Myocastor coypus in a gated community in the metropolitan area of Buenos Aires (Argentina). J. Urban Ecol. 2022, 8, juab037. [Google Scholar] [CrossRef]
- Mori, E.; Mazza, G.; Pucci, C.; Senserini, D.; Campbell-Palmer, R.; Contaldo, M.; Viviano, A. Temporal activity patterns of the Eurasian beaver and coexisting species in a mediterranean ecosystem. Animals 2022, 12, 1961. [Google Scholar] [CrossRef] [PubMed]
- Zschille, J.; Stier, N.; Roth, M. Gender differences in activity patterns of American mink Neovison Vison in Germany. Eur. J. Wildl. Res. 2010, 56, 187–194. [Google Scholar] [CrossRef]
- Davis, R.A.; Jenson, A.G. A Note on the distribution of the coypu (Myocastor coypus) in Great Britain. J. Anim. Ecol. 1960, 29, 397. [Google Scholar] [CrossRef]
- Gosling, L.M.; Guyon, G.E.; Wright, K.M.H. Diurnal activity of feral coypus (Myocastor coypus) during the cold winter of 1978–9. J. Zool. 1980, 192, 143–146. [Google Scholar] [CrossRef]
- Gosling, L.M. The twenty-four-hour activity cycle of captive coypus Myocastor coypus. J. Zool. 1979, 187, 341–367. [Google Scholar] [CrossRef]
- Prigioni, C.; Balestrieri, A.; Remonti, L. Food habits of the coypu, Myocastor coypus, and its impact on aquatic vegetation in a freshwater habitat of NW Italy. Folia Zool. 2005, 54, 269–277. [Google Scholar]
- Marini, F.; Gabrielli, E.; Montaudo, L.; Vecchi, M.; Santoro, R.; Battisti, C.; Carpaneto, G.M. Diet of coypu (Myocastor coypus) in a mediterranean coastal wetland: A possible impact on threatened Rushbeds? Vie Milieu 2013, 63, 97–103. [Google Scholar]
- Lomnicki, A. The daily rhythm of activity in the nutria (Myocastor coypus Molina). Foila Biol. 1957, 5, 293–306. [Google Scholar]
- Meyer, J.; Klemann, N.; Halle, S. Diurnal activity patterns of coypu in an urban habitat. Acta Theriol. 2005, 50, 207–211. [Google Scholar] [CrossRef]
- Darimont, C.T.; Carlson, S.M.; Kinnison, M.T.; Paquet, P.C.; Reimchen, T.E.; Wilmers, C.C. Human predators outpace other agents of trait change in the wild. Proc. Natl. Acad. Sci. USA 2009, 106, 952–954. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Kim, G.W.; Park, Y. Habitat suitability and connectivity modeling predict genetic population structure and priority control areas for invasive nutria (Myocastor coypus) in a temperate river basin. PLoS ONE 2022, 17, e0279082. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; An, J. Assessing future climate change impact on hydrologic and water quality components in Nakdong River Basin. J. Korea Water Resour. Assoc. 2012, 45, 121–130. [Google Scholar] [CrossRef]
- Allendorf, F.W.; Luikart, G.H.; Aitken, S.N. Conservation and the Genetics of Populations; Wiley-Blackwell Publication: Oxford, UK, 2013. [Google Scholar]
- Panzacchi, M.; Bertolino, S.; Cocchi, R.; Genovesi, P. Population control of coypu (Myocastor coypus) in Italy compared to eradication in UK: A cost-benefit analysis. Wildl. Biol. 2007, 13, 159–171. [Google Scholar] [CrossRef]
- Bertolino, S.; Viterbi, R. Long-Term cost-effectiveness of coypu (Myocastor coypus) control in Piedmont (Italy). Biol. Invasions 2010, 12, 2549–2558. [Google Scholar] [CrossRef]
- Tunez, J.I.; Guichón, M.L.; Centrón, D.; Henderson, A.P.; Callahan, C.; Cassini, M.H. Relatedness and social organization of coypus in the Argentinean pampas. Mol. Ecol. 2009, 18, 147–155. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Method | Male (n = 11) | Female (n = 12) | Sex Difference | |
|---|---|---|---|---|---|
| t | p | ||||
| Spring (Mar–May) | MCP | 0.37 ± 1.3 | 0.25 ± 2.4 | 1.81 | 0.043 |
| KDE95 | 0.51 ± 1.2 | 0.35 ± 3.2 | 3.22 | 0.004 | |
| KDE50 | 0.09 ± 2.3 | 0.05 ± 0.7 | 2.32 | 0.021 | |
| Summer (Jun–Aug) | MCP | 0.32 ± 2.1 | 0.24 ± 1.0 | 2.26 | 0.010 |
| KDE95 | 0.44 ± 2.2 | 0.37 ± 2.1 | 3.61 | 0.012 | |
| KDE50 | 0.06 ± 3.1 | 0.02 ± 1.5 | 2.06 | 0.022 | |
| Autumn (Sep–Nov) | MCP | 0.47 ± 3.2 | 0.29 ± 0.5 | 3.38 | 0.004 |
| KDE95 | 0.62 ± 1.4 | 0.56 ± 1.2 | 1.10 | 0.034 | |
| KDE50 | 0.13 ± 0.3 | 0.06 ± 0.3 | 2.81 | 0.015 | |
| Winter (Dec–Feb) | MCP | 0.28 ± 1.3 | 0.17 ± 3.0 | 2.32 | 0.001 |
| KDE95 | 0.31 ± 2.1 | 0.31 ± 0.6 | 0.49 | 0.064 * | |
| KDE50 | 0.05 ± 3.2 | 0.01 ± 1.0 | 1.46 | 0.010 | |
| Average | MCP | 0.36 ± 0.6 | 0.23 ± 0.5 | 2.26 | 0.030 |
| KDE95 | 0.47 ± 1.5 | 0.39 ± 0.2 | 2.25 | 0.006 | |
| KDE50 | 0.08 ± 2.1 | 0.03 ± 0.1 | 1.80 | 0.042 | |
| Seasons | Trap Locations ± S.E. | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| Twilight | Day | Night | M | F | ||||
| M | F | M | F | M | F | |||
| Spring | 28.2 ± 1.20 | 25.1 ± 10 | 15.2 ± 0.3 | 13.1 ± 0.43 | 20.1 ± 0.21 | 16.2 ± 0.60 | 0.12 | 0.11 |
| Summer | 31.1 ± 0.18 | 27.2 ± 0.27 | 19.1 ± 0.84 | 21.3 ± 0.16 | 32.6 ± 0.01 | 26.2 ± 0.11 | 0.61 | 0.32 |
| Autumn | 21.1 ± 0.16 | 17.6 ± 0.15 | 14.1 ± 0.17 | 13.1 ± 0.11 | 22.1 ± 11 | 17.6 ± 0.07 | 0.07 | 0.13 |
| Winter | 12.3 ± 0.31 | 12.6 ± 1.2 | 10.4 ± 0.27 | 9.1 ± 0.12 | 7.3 ± 0.21 | 8.4 ± 0.16 | 0.04 * | 0.02 * |
| p-value | 0.03 * | 0.01 * | 0.06 | 0.10 | 0.31 | 0.51 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banjade, M.; Adhikari, P.; Hong, S.-H.; Lee, D.-H. Radio Tracking Reveals the Home Range and Activity Patterns of Nutria (Myocastor coypus) in the Macdo Wetland in South Korea. Animals 2023, 13, 1716. https://doi.org/10.3390/ani13101716
Banjade M, Adhikari P, Hong S-H, Lee D-H. Radio Tracking Reveals the Home Range and Activity Patterns of Nutria (Myocastor coypus) in the Macdo Wetland in South Korea. Animals. 2023; 13(10):1716. https://doi.org/10.3390/ani13101716
Chicago/Turabian StyleBanjade, Maniram, Pradeep Adhikari, Sun-Hee Hong, and Do-Hun Lee. 2023. "Radio Tracking Reveals the Home Range and Activity Patterns of Nutria (Myocastor coypus) in the Macdo Wetland in South Korea" Animals 13, no. 10: 1716. https://doi.org/10.3390/ani13101716