
The Linear-Logistic Model: A Novel Paradigm for Estimating Dietary Amino Acid Requirements


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
- Only studies written in English were considered;
- Data from abstracts and oral presentations were not considered;
- Studies that provided AA in a wide-enough range to detect the linear-logistic pattern in protein retention and growth measurements were considered;
- Growth was considered an indirect measure of protein retention in stages other than gestation, as the products of conception are less protein-dense than lean tissue [22]; therefore, increases in body weight may not necessarily indicate increased protein retention during gestation;
- Only studies that measured the effect of AA intake on physiological responses related to reproduction, health, and survivability, in addition to growth or protein retention variables, were considered;
- Studies that measured plasma AA concentrations, in addition to growth or protein retention variables, were considered;
- Studies that reported methodological issues, such as unexplained responses caused by factors other than dietary AA intake, were not considered.
- Studies in which physiological responses did not follow a clear trend were considered to have random fluctuations (i.e., unexplained variance) and thus not considered.
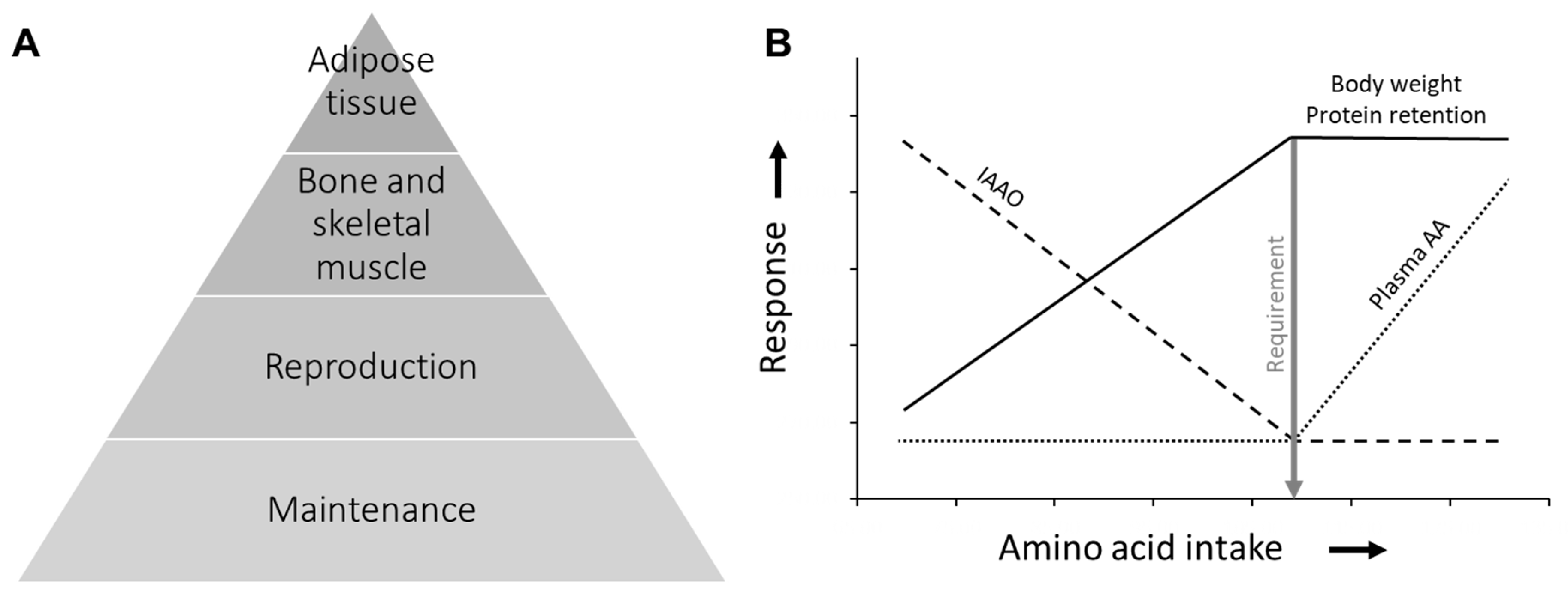
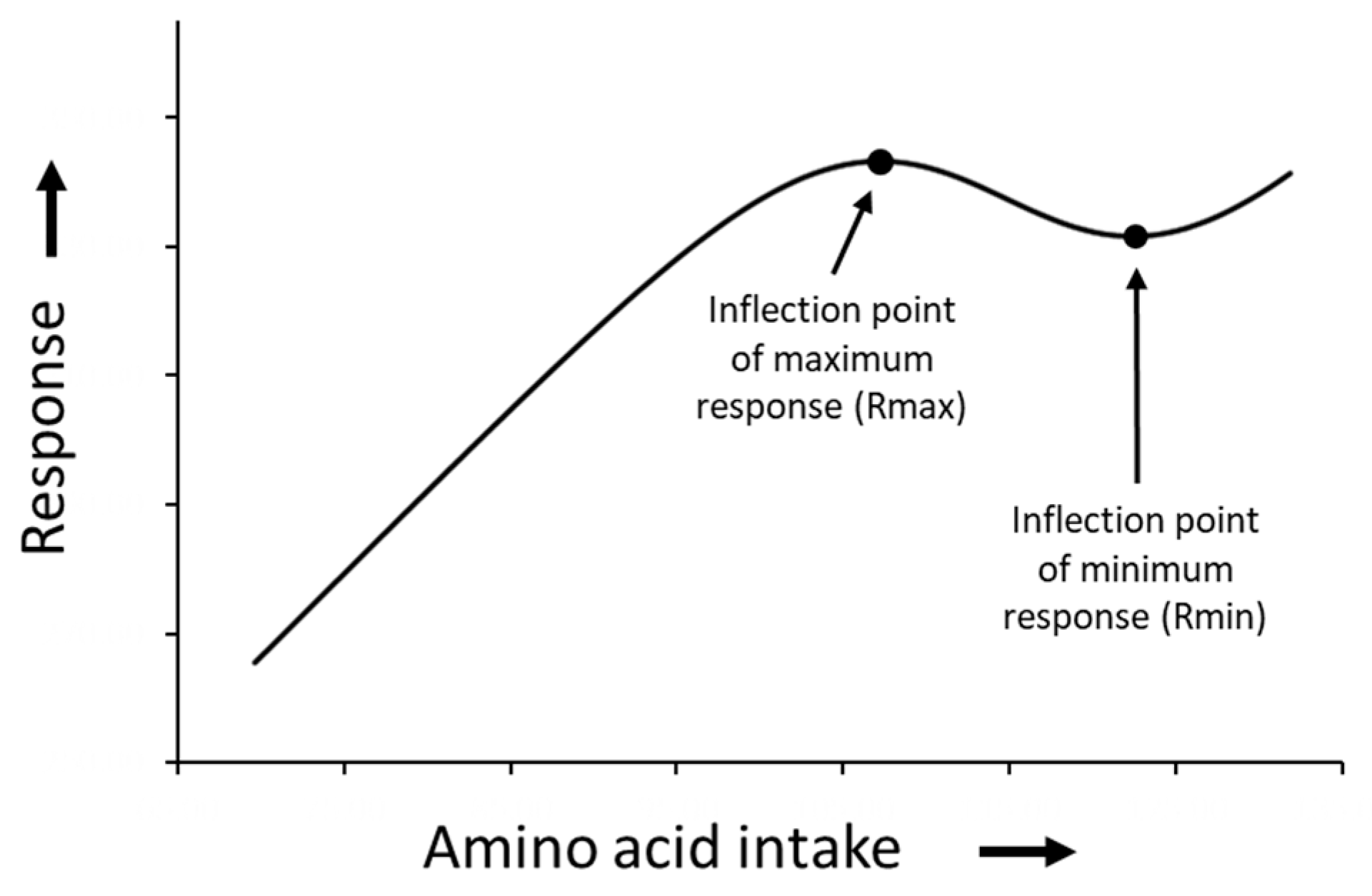
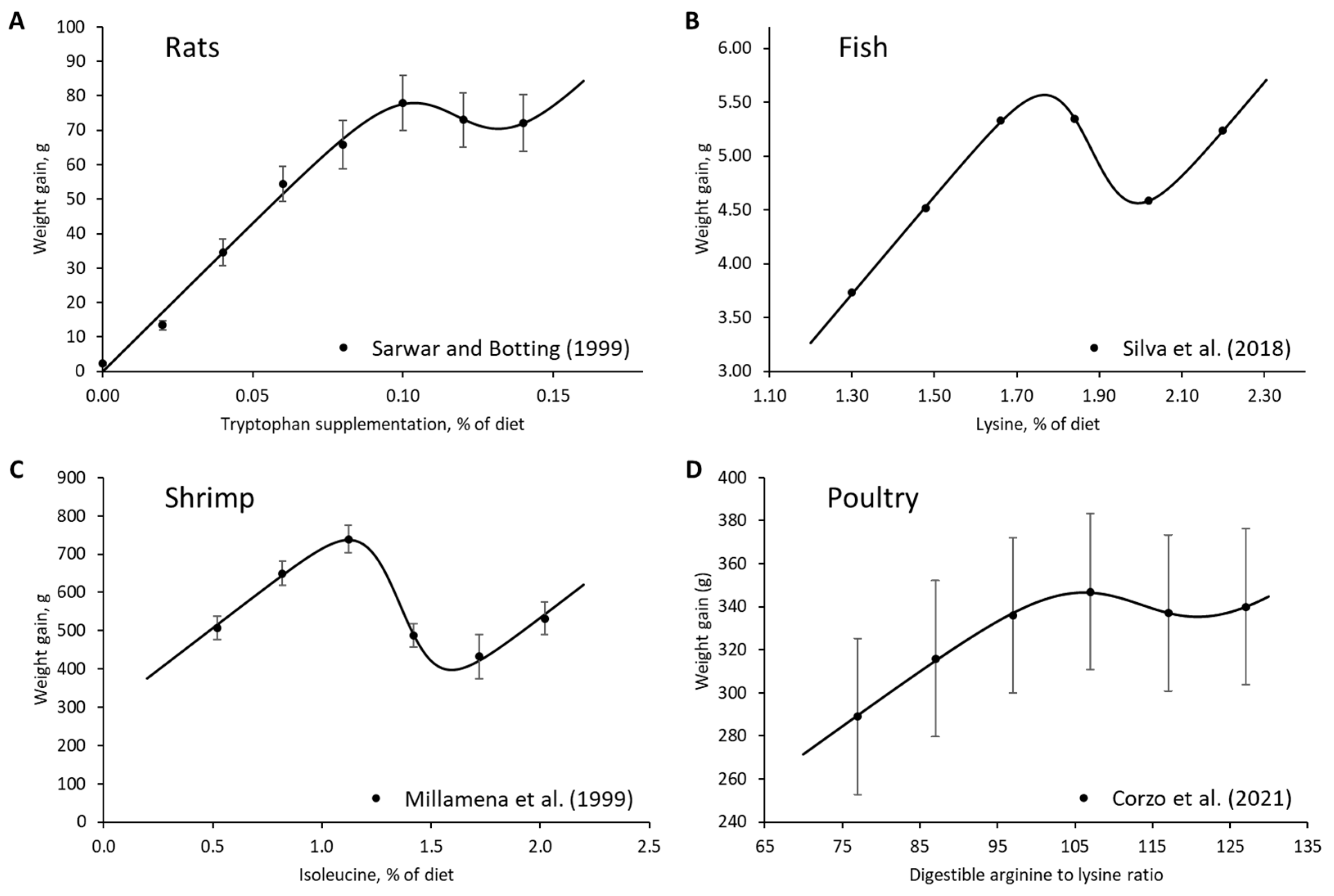
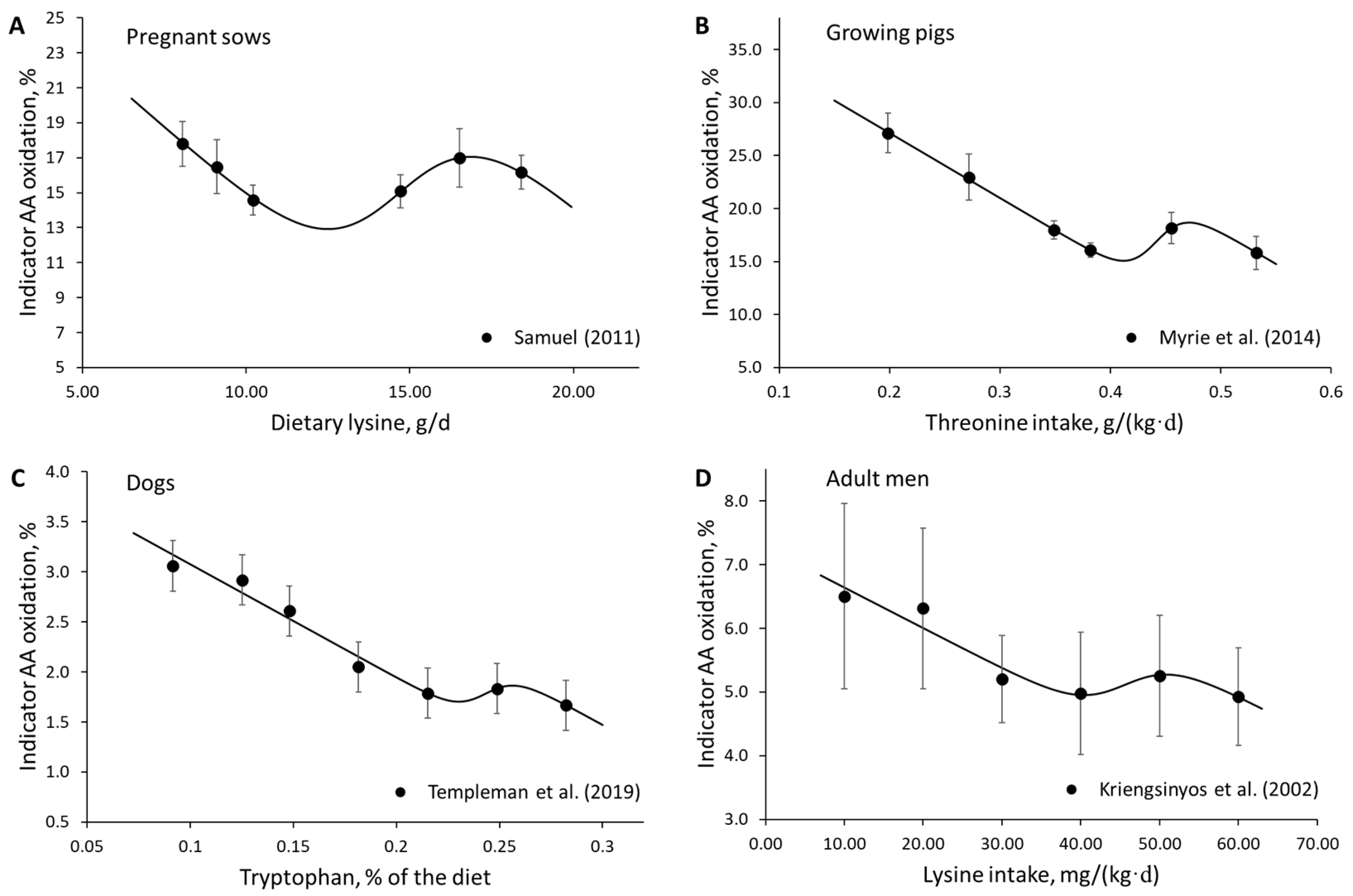
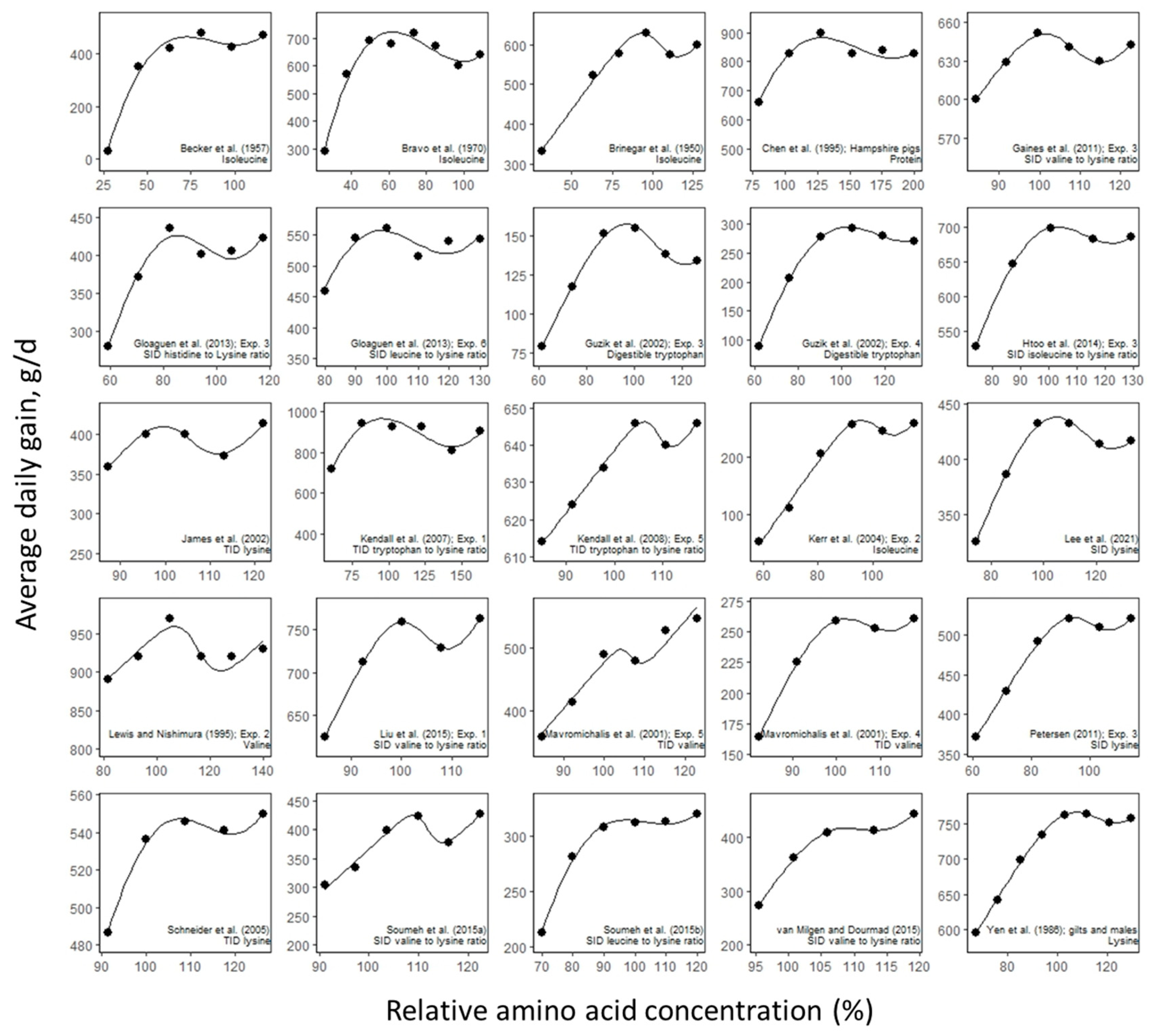
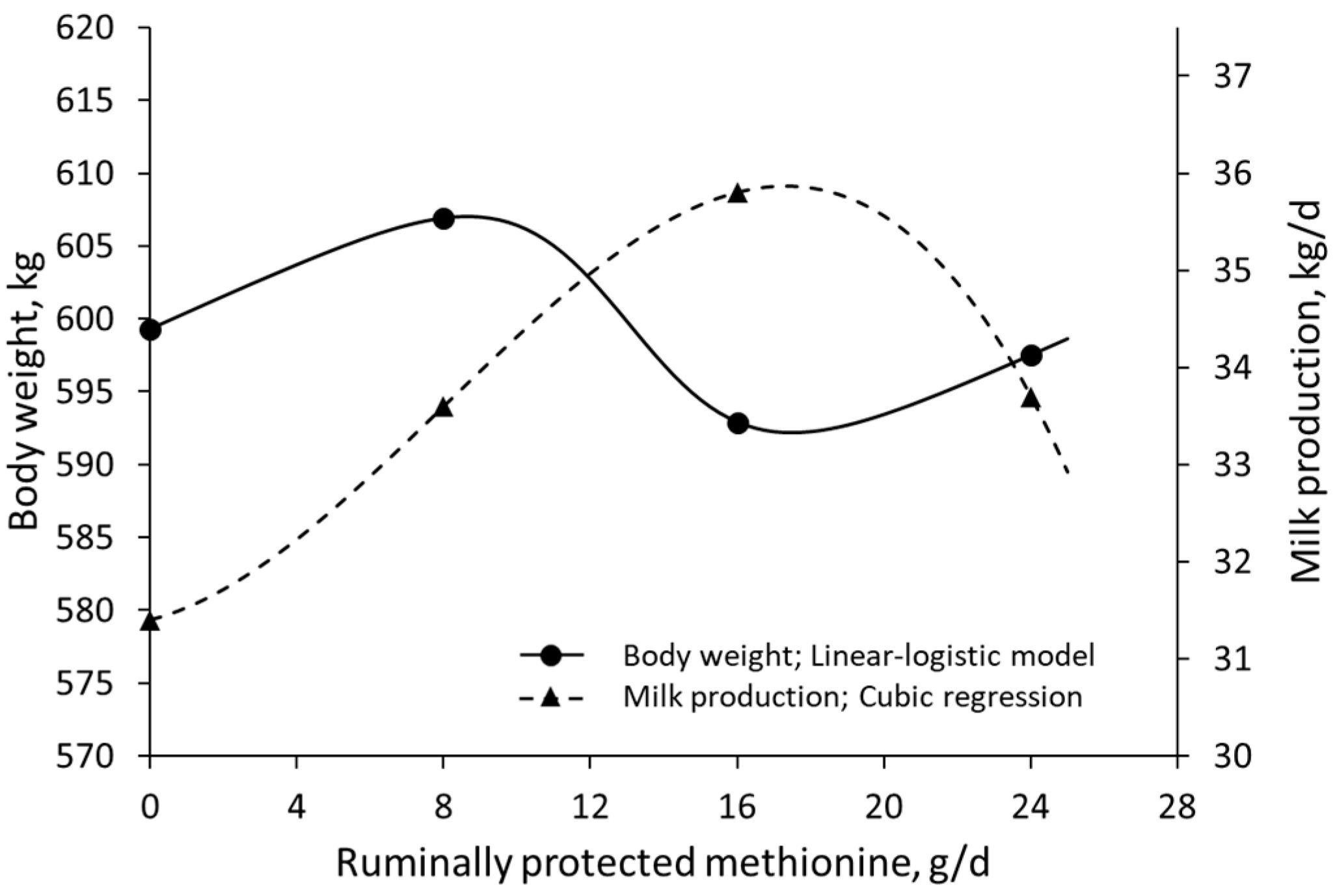
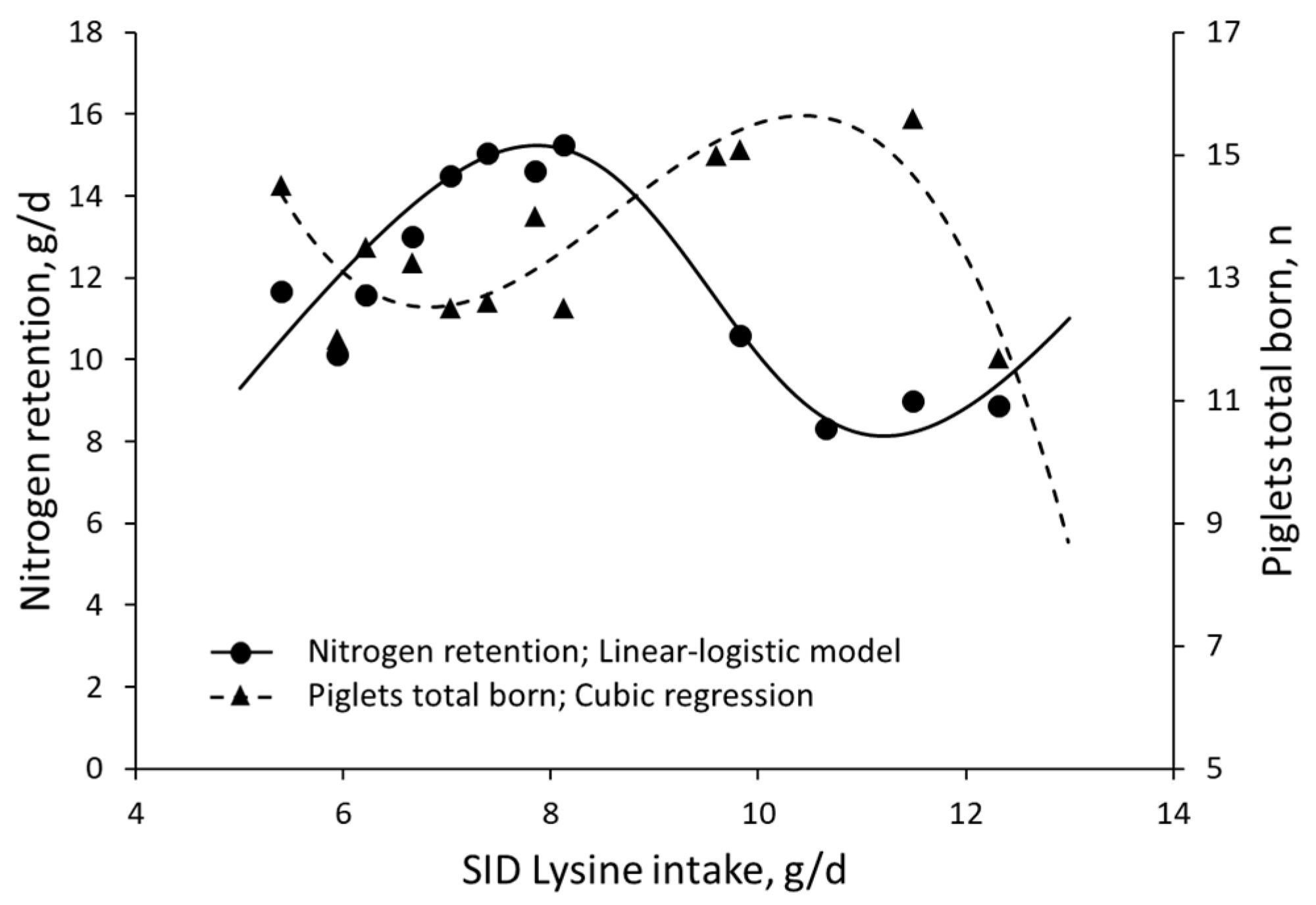
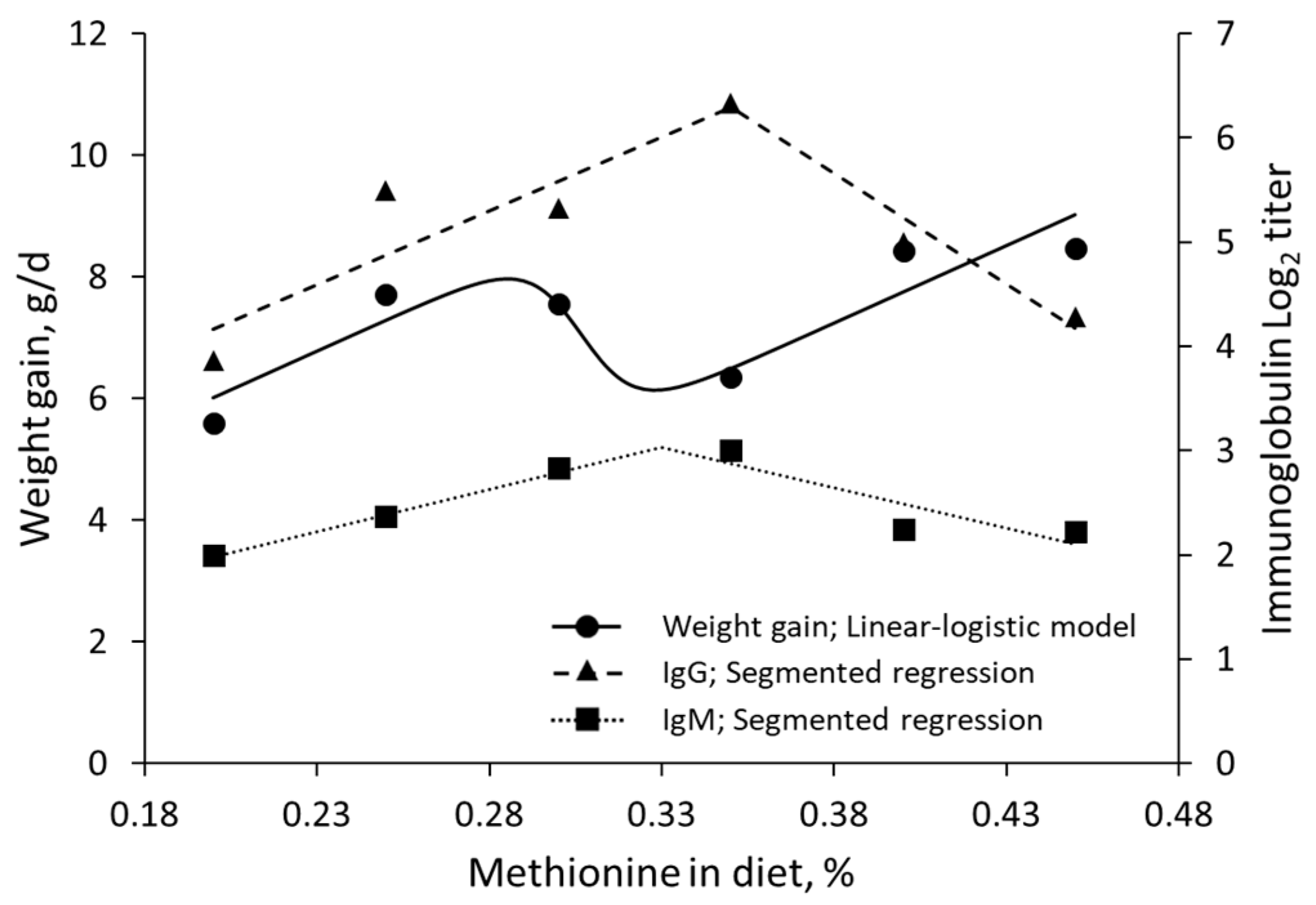
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Monteiro, M.S.; Matias, D.N.; Poor, A.P.; Dutra, M.C.; Moreno, L.Z.; Parra, B.M.; Silva, A.P.S.; Matajira, C.E.C.; de Moura Gomes, V.T.; Barbosa, M.R.F. Causes of Sow Mortality and Risks to Post-Mortem Findings in a Brazilian Intensive Swine Production System. Animals 2022, 12, 1804. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, J.T.; Thomsen, R. Identification of Risk Factors and Strategies for Reducing Sow Mortality; DCA-Report. Available online: https://dcapub.au.dk/djfpdf/DCArapport097.pdf (accessed on 4 March 2023).
- Kirk, R.; Svensmark, B.; Ellegaard, L.; Jensen, H. Locomotive disorders associated with sow mortality in Danish pig herds. J. Vet. Med. 2005, 52, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Kikuti, M.; Preis, G.M.; Deen, J.; Pinilla, J.C.; Corzo, C.A. Sow mortality in a pig production system in the midwestern USA: Reasons for removal and factors associated with increased mortality. Vet. Rec. 2022, 192, e2539. [Google Scholar] [CrossRef] [PubMed]
- Baxter, E.M.; Edwards, S.A. Chapter 3: Piglet mortality and morbidity: Inevitable or unacceptable. In Advances in Pig Welfare; Špinka, M., Ed.; Elsevier: Cambridge, MA, USA, 2018; pp. 73–100. [Google Scholar] [CrossRef]
- Muns, R.; Nuntapaitoon, M.; Tummaruk, P. Non-infectious causes of pre-weaning mortality in piglets. Livest. Sci. 2016, 184, 46–57. [Google Scholar] [CrossRef]
- Gebhardt, J.T.; Tokach, M.D.; Dritz, S.S.; DeRouchey, J.M.; Woodworth, J.C.; Goodband, R.D.; Henry, S.C. Postweaning mortality in commercial swine production. I: Review of non-infectious contributing factors. Transl. Anim. Sci. 2020, 4, 462–484. [Google Scholar] [CrossRef]
- Stalder, K.J. Pork Industry Productivity Analysis; National Pork Board Report. Available online: https://jygatech.com/wp-content/uploads/2021/05/PigmortalityStalderIowa.pdf (accessed on 22 August 2022).
- Iannetti, L.; Romagnoli, S.; Cotturone, G.; Podaliri Vulpiani, M. welfare assessment in antibiotic-free and conventional broiler chicken. Animals 2021, 11, 2822. [Google Scholar] [CrossRef]
- Putman, B.; Thoma, G.; Burek, J.; Matlock, M. A retrospective analysis of the United States poultry industry: 1965 compared with 2010. Agric. Syst. 2017, 157, 107–117. [Google Scholar] [CrossRef]
- Junghans, A.; Deseniß, L.; Louton, H. Data evaluation of broiler chicken rearing and slaughter—An exploratory study. Front. Vet. Sci. 2022, 9, 957786. [Google Scholar] [CrossRef]
- Edwards, S.A.; Baxter, E. Piglet mortality: Causes and prevention. In The Gestating and Lactating Sow; Farmer, C., Ed.; Academic Publishers: Wageningen, The Netherlands, 2015; pp. 193–230. [Google Scholar] [CrossRef]
- Chalvon-Demersay, T.; Luise, D.; Le Floc’H, N.; Tesseraud, S.; Lambert, W.; Bosi, P.; Trevisi, P.; Beaumont, M.; Corrent, E. Functional amino acids in pigs and chickens: Implication for gut health. Front. Vet. Sci. 2021, 8, 663727. [Google Scholar] [CrossRef]
- Robinson, J.L.; Bartlett, R.K.; Harding, S.V.; Randell, E.W.; Brunton, J.A.; Bertolo, R.F. Dietary methyl donors affect in vivo methionine partitioning between transmethylation and protein synthesis in the neonatal piglet. Amino Acids 2016, 48, 2821–2830. [Google Scholar] [CrossRef]
- Marchini, J.S.; Cortiella, J.; Hiramatsu, T.; Chapman, T.E.; Young, V.R. Requirements for indispensable amino acids in adult humans: Longer-term amino acid kinetic study with support for the adequacy of the Massachusetts Institute of Technology amino acid requirement pattern. Am. J. Clin. Nutr. 1993, 58, 670–683. [Google Scholar] [CrossRef] [PubMed]
- Tukey, J.W. Exploratory Data Analysis; Pearson: London, UK, 1977; Volume 2. [Google Scholar]
- Hammond, J. Physiological factors affecting birth weight. Proc. Nutr. Soc. 1944, 2, 8. [Google Scholar]
- Elango, R.; Ball, R.O.; Pencharz, P.B. Recent advances in determining protein and amino acid requirements in humans. Br. J. Nutr. 2012, 108, S22–S30. [Google Scholar] [CrossRef] [PubMed]
- France, J.; Kebreab, E. Mathematical Modelling in Animal Nutrition; CABI: Wallingford, UK, 2008. [Google Scholar]
- Young, V.R.; Marchini, J.S. Mechanisms and nutritional significance of metabolic responses to altered intakes of protein and amino acids, with reference to nutritional adaptation in humans. Am. J. Clin. Nutr. 1990, 51, 270–289. [Google Scholar] [CrossRef] [PubMed]
- Mikolajewicz, N.; Komarova, S.V. Meta-analytic methodology for basic research: A practical guide. Front. Physiol. 2019, 10, 203. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.; Ma, Y.; Payne, R.; Lindemann, M. Amino acid composition of fetus, placenta, and uterus in gilts throughout gestation. J. Anim. Sci. 2017, 95, 4448–4461. [Google Scholar] [CrossRef]
- Sarwar, G.; Botting, H.G. Liquid concentrates are lower in bioavailable tryptophan than powdered infant formulas, and tryptophan supplementation of formulas increases brain tryptophan and serotonin in rats. J. Nutr. 1999, 129, 1692–1697. [Google Scholar] [CrossRef]
- Silva, J.C.; Bomfim, M.A.D.; Lanna, E.A.T.; Ribeiro, F.B.; de Siqueira, J.C.; de Sousa, T.J.R.; Marchão, R.S.; do Nascimento, D.C.N. Lysine requirement for tambaqui juveniles. Semin. Cienc. Agrar. 2018, 39, 2157–2168. [Google Scholar] [CrossRef]
- Millamena, O.M.; Teruel, M.; Kanazawa, A.; Teshima, S. Quantitative dietary requirements of postlarval tiger shrimp, Penaeus monodon, for histidine, isoleucine, leucine, phenylalanine and tryptophan. Aquaculture 1999, 179, 169–179. [Google Scholar] [CrossRef]
- Corzo, A.; Lee, J.; Vargas, J.; Silva, M.; Pacheco, W. Determination of the optimal digestible arginine to lysine ratio in Ross 708 male broilers. J. Appl. Poult. Res. 2021, 30, 100136. [Google Scholar] [CrossRef]
- Samuel, R.S. Simultaneous Measurement of Protein and Energy Metabolism and Application to Determine Lysine Requirements in Sows. Ph.D. Dissertation, University of Alberta, Edmonton, AB, Canada, 2011. [Google Scholar] [CrossRef]
- Myrie, S.B.; Bertolo, R.F.; Moehn, S.; Ball, R.O. Barley does not change threonine requirement in growing pigs fed a barley–casein-based diet compared to a casein-based diet, as determined by the indicator amino acid oxidation method. Livest. Sci. 2014, 170, 108–115. [Google Scholar] [CrossRef]
- Templeman, J.R.; Mansilla, W.D.; Fortener, L.; Shoveller, A.K. Tryptophan requirements in small, medium, and large breed adult dogs using the indicator amino acid oxidation technique. J. Anim. Sci. 2019, 97, 3274–3285. [Google Scholar] [CrossRef] [PubMed]
- Kriengsinyos, W.; Wykes, L.J.; Ball, R.O.; Pencharz, P.B. Oral and intravenous tracer protocols of the indicator amino acid oxidation method provide the same estimate of the lysine requirement in healthy men. J. Nutr. 2002, 132, 2251–2257. [Google Scholar] [CrossRef]
- Becker, D.; Jensen, A.; Terrill, S.; Smith, I.; Norton, H. The isoleucine requirement of weanling swine fed two protein levels. J. Anim. Sci. 1957, 16, 26–34. [Google Scholar] [CrossRef]
- Bravo, F.; Meade, R.; Stockland, W.; Nordstrom, J. Reevaluation of the Insoleucine Requirement of the Growing Pig—Plasma Free Isoleucine as a Response Criterion. J. Anim. Sci. 1970, 31, 1137–1141. [Google Scholar] [CrossRef]
- Brinegar, M.; Loosli, J.; Maynard, L.; Williams, H. The isoleucine requirement for the growth of swine. J. Nutr. 1950, 42, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Miller, P.; Lewis, A.; Wolverton, C.; Stroup, W. Changes in plasma urea concentration can be used to determine protein requirements of two populations of pigs with different protein accretion rates. J. Anim. Sci. 1995, 73, 2631–2639. [Google Scholar] [CrossRef]
- Gaines, A.; Kendall, D.; Allee, G.; Usry, J.; Kerr, B. Estimation of the standardized ileal digestible valine-to-lysine ratio in 13-to 32-kilogram pigs. J. Anim. Sci. 2011, 89, 736–742. [Google Scholar] [CrossRef] [PubMed]
- Gloaguen, M.; Le Floc’H, N.; Primot, Y.; Corrent, E.; van Milgen, J. Response of piglets to the standardized ileal digestible isoleucine, histidine and leucine supply in cereal–soybean meal-based diets. Animal 2013, 7, 901–908. [Google Scholar] [CrossRef]
- Guzik, A.; Southern, L.; Bidner, T.; Kerr, B. The tryptophan requirement of nursery pigs. J. Anim. Sci. 2002, 80, 2646–2655. [Google Scholar] [CrossRef]
- Htoo, J.; Zhu, C.; Huber, L.; de Lange, C.; Quant, A.; Kerr, B.; Cromwell, G.; Lindemann, M. Determining the optimal isoleucine: Lysine ratio for 10–22 kg and 24–39 kg pigs fed diets containing non-excess levels of leucine. J. Anim. Sci. 2014, 92, 3482–3490. [Google Scholar] [CrossRef] [PubMed]
- James, B.; Hastad, C.; Lawrence, K.; Usry, J.; Tokach, M.D.; Goodband, R.D.; Nelssen, J.L.; Dritz, S.S. The optimal true ileal digestible lysine requirement for nursery pigs between 27 to 44 lb. Kansas Agric. Exp. Stn. Res. Rep. 2002, 63–65. [Google Scholar] [CrossRef]
- Kendall, D.; Gaines, A.; Kerr, B.; Allee, G. True ileal digestible tryptophan to lysine ratios in ninety-to one hundred twenty-five-kilogram barrows. J. Anim. Sci. 2007, 85, 3004–3012. [Google Scholar] [CrossRef]
- Kendall, D.; Gaines, A.; Allee, G.; Usry, J. Commercial validation of the true ileal digestible lysine requirement for eleven-to twenty-seven-kilogram pigs. J. Anim. Sci. 2008, 86, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Kerr, B.; Kidd, M.; Cuaron, J.; Bryant, K.; Parr, T.; Maxwell, C.; Campbell, J. Isoleucine requirements and ratios in starting (7 to 11 kg) pigs. J. Anim. Sci. 2004, 82, 2333–2342. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; González-Vega, J.; Htoo, J.; Nyachoti, C. Evaluating the standardized ileal digestible lysine requirement of 7-to 15-kg weanling pigs fed corn-soybean meal-based diets. Animal 2021, 15, 100127. [Google Scholar] [CrossRef]
- Lewis, A.; Nishimura, N. Valine requirement of the finishing pig. J. Anim. Sci. 1995, 73, 2315–2318. [Google Scholar] [CrossRef]
- Liu, X.; Ma, W.; Zeng, X.; Xie, C.; Thacker, P.; Htoo, J.; Qiao, S. Estimation of the standardized ileal digestible valine to lysine ratio required for 25-to 120-kilogram pigs fed low crude protein diets supplemented with crystalline amino acids. J. Anim. Sci. 2015, 93, 4761–4773. [Google Scholar] [CrossRef]
- Mavromichalis, I.; Kerr, B.; Parr, T.; Albin, D.; Gabert, V.; Baker, D. Valine requirement of nursery pigs. J. Anim. Sci. 2001, 79, 1223–1229. [Google Scholar] [CrossRef]
- Petersen, G.I. Estimation of the Ideal Standardized Ileal Digestible Tryptophan: Lysine Ratio in 10 to 20 kg Pigs. Ph.D. Dissertation, University of Illinois at Urbana-Champaign, Urbana, IL, USA, 2011. [Google Scholar]
- Schneider, J.; Tokach, M.D.; Goodband, R.D.; Nelssen, J.L.; DeRouchey, J.M.; Dritz, S.S. The optimal true ileal digestible lysine and total sulfur amino acid requirement for nursery pigs between 20 and 50 lb. In Proceedings of the Swine Day Conference, Manhattan, KS, USA, November 2005; Kansas State University: Manhattan, KS, USA, 2005. [Google Scholar]
- Soumeh, E.A.; Van Milgen, J.; Sloth, N.M.; Corrent, E.; Poulsen, H.D.; Nørgaard, J.V. Requirement of standardized ileal digestible valine to lysine ratio for 8-to 14-kg pigs. Animal 2015, 9, 1312–1318. [Google Scholar] [CrossRef]
- Soumeh, E.A.; Van Milgen, J.; Sloth, N.M.; Corrent, E.; Poulsen, H.D.; Nørgaard, J.V. The optimum ratio of standardized ileal digestible leucine to lysine for 8 to 12 kg female pigs. J. Anim. Sci. 2015, 93, 2218–2224. [Google Scholar] [CrossRef] [PubMed]
- van Milgen, J.; Dourmad, J.-Y. Concept and application of ideal protein for pigs. J. Anim. Sci. Biotechnol. 2015, 6, 15. [Google Scholar] [CrossRef] [PubMed]
- Yen, H.; Cole, D.; Lewis, D. Amino acid requirements of growing pigs. 7. The response of pigs from 25 to 55 kg live weight to dietary ideal protein. Anim. Prod. 1986, 43, 141–154. [Google Scholar] [CrossRef]
- Lara, A.; Mendoza, G.; Landois, L.; Barcena, R.; Sánchez-Torres, M.; Rojo, R.; Ayala, J.; Vega, S. Milk production in Holstein cows supplemented with different levels of ruminally protected methionine. Livest. Sci. 2006, 105, 105–108. [Google Scholar] [CrossRef]
- Ramirez-Camba, C.D.; Dunn, J.L.; Htoo, J.K.; González-Vega, J.C.; Touchette, K.; Samuel, R.S.; Levesque, C.L. Efficiency of standardized ileal digestible lysine utilization for whole body protein deposition in pregnant gilts and sows during early-, mid-, and late-gestation. J. Anim. Sci. 2020, 98, skaa340. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A.; Peo, E., Jr.; Moser, B.; Crenshaw, T. Lysine requirement of pigs weighing 5 to 15 kg fed practical diets with and without added fat. J. Anim. Sci. 1980, 51, 361–366. [Google Scholar] [CrossRef]
- Hosseini, S.A.; Zaghari, M.; Lotfollahian, H.; Shivazad, M.; Moravaj, H. Reevaluation of methionine requirement based on performance and immune responses in broiler breeder hens. J. Poult. Sci. 2011, 49, 26–33. [Google Scholar] [CrossRef]
- Jayaraman, B.; Htoo, J.; Nyachoti, C.M. Effects of dietary threonine: Lysine ratioes and sanitary conditions on performance, plasma urea nitrogen, plasma-free threonine and lysine of weaned pigs. Anim. Nutr. 2015, 1, 283–288. [Google Scholar] [CrossRef]
- Xiao, W.; Li, D.; Zhu, J.; Zou, Z.; Yue, Y.; Yang, H. Dietary valine requirement of juvenile Nile tilapia, Oreochromis niloticus. Aquac. Nutr. 2018, 24, 315–323. [Google Scholar] [CrossRef]
- Remus, A.; Hauschild, L.; Corrent, E.; Létourneau-Montminy, M.-P.; Pomar, C. Pigs receiving daily tailored diets using precision-feeding techniques have different threonine requirements than pigs fed in conventional phase-feeding systems. J. Anim. Sci. Biotechnol. 2019, 10, 16. [Google Scholar] [CrossRef]
- Wellington, M.O.; Thiessen, R.B.; Van Kessel, A.G.; Columbus, D.A. Intestinal health and threonine requirement of growing pigs fed diets containing high dietary fibre and fermentable protein. Animals 2020, 10, 2055. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Guo, Y. Supplemental zinc reduced intestinal permeability by enhancing occludin and zonula occludens protein-1 (ZO-1) expression in weaning piglets. Br. J. Nutr. 2009, 102, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Reigh, R.C.; Williams, M.B. Growth effects of lysine in compounded diets for American alligator, Alligator mississippiensis. J. World Aquac. Soc. 2022, 53, 995–1005. [Google Scholar] [CrossRef]
- Bhargava, K.; Hanson, R.; Sunde, M. Effects of methionine and valine on antibody production in chicks infected with Newcastle disease virus. J. Nutr. 1970, 100, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Riedijk, M.A.; Stoll, B.; Chacko, S.; Schierbeek, H.; Sunehag, A.L.; van Goudoever, J.B.; Burrin, D.G. Methionine transmethylation and transsulfuration in the piglet gastrointestinal tract. Proc. Natl. Acad. Sci. USA 2007, 104, 3408–3413. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, D.; Dai, Z.; Piao, X.; Wu, Z.; Wang, B.; Zhu, Y.; Zeng, Z. L-methionine supplementation maintains the integrity and barrier function of the small-intestinal mucosa in post-weaning piglets. Amino Acids 2014, 46, 1131–1142. [Google Scholar] [CrossRef]
- Holm, M.B.; Kristiansen, O.; Holme, A.M.; Bastani, N.E.; Horne, H.; Blomhoff, R.; Haugen, G.; Henriksen, T.; Michelsen, T.M. Placental release of taurine to both the maternal and fetal circulations in human term pregnancies. Amino Acids 2018, 50, 1205–1214. [Google Scholar] [CrossRef]
- Ripps, H.; Shen, W. Review: Taurine: A “very essential” amino acid. Mol. Vis. 2012, 18, 2673. [Google Scholar]
- Wen, C.; Li, F.; Zhang, L.; Duan, Y.; Guo, Q.; Wang, W.; He, S.; Li, J.; Yin, Y. Taurine is involved in energy metabolism in muscles, adipose tissue, and the liver. Mol. Nutr. Food Res. 2019, 63, 1800536. [Google Scholar] [CrossRef]
- Tang, Z.; Liu, J.; Sun, Z.; Li, J.; Sun, W.; Mao, J.; Wang, Y. Protective effects of taurine on growth performance and intestinal epithelial barrier function in weaned piglets challenged without or with lipopolysaccharide. Anim. Prod. Sci. 2018, 58, 2011–2020. [Google Scholar] [CrossRef]
- Mu, T.; Feng, Y.; Che, Y.; Lv, Q.; Hu, J.; Yang, Q.; Yang, J. Taurine Promotes In-vitro Follicle Development, Oocyte Maturation, Fertilization and Cleavage of rats. In Taurine 11; Hu, J., Piao, F., Schaffer, S., El Idrissi, A., Wu, J.Y., Eds.; Springer: Singapore, 2019; Volume 1155, pp. 197–203. [Google Scholar] [CrossRef]
- Norberg, S.; Powell, T.L.; Jansson, T. Intrauterine growth restriction is associated with a reduced activity of placental taurine transporters. Pediatr. Res. 1998, 44, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, Y.; Obayashi, M.; Murakami, T.; Harris, R.A. Regulation of branched-chain amino acid catabolism: Nutritional and hormonal regulation of activity and expression of the branched-chain α-keto acid dehydrogenase kinase. Curr. Opin. Clin. Nutr. Metab. Care 2001, 4, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.T.; Sweatt, A.J.; Hutson, S.M. Expression of mitochondrial branched-chain aminotransferase and α-keto-acid dehydrogenase in rat brain: Implications for neurotransmitter metabolism. Front. Neuroanat. 2012, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Holm, M.B.; Bastani, N.E.; Holme, A.M.; Zucknick, M.; Jansson, T.; Refsum, H.; Mørkrid, L.; Blomhoff, R.; Henriksen, T.; Michelsen, T.M. Uptake and release of amino acids in the fetal-placental unit in human pregnancies. PLoS ONE 2017, 12, e0185760. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Feng, D.; Zhang, Y.; Dahanayaka, S.; Li, X.; Yao, K.; Wang, J.; Wu, Z.; Dai, Z.; Wu, G. Regulation of leucine catabolism by metabolic fuels in mammary epithelial cells. Amino Acids 2012, 43, 2179–2189. [Google Scholar] [CrossRef]
- Sun, Y.; Wu, Z.; Li, W.; Zhang, C.; Sun, K.; Ji, Y.; Wang, B.; Jiao, N.; He, B.; Wang, W. Dietary L-leucine supplementation enhances intestinal development in suckling piglets. Amino Acids 2015, 47, 1517–1525. [Google Scholar] [CrossRef]
- Chen, L.; Li, P.; Wang, J.; Li, X.; Gao, H.; Yin, Y.; Hou, Y.; Wu, G. Catabolism of nutritionally essential amino acids in developing porcine enterocytes. Amino Acids 2009, 37, 143–152. [Google Scholar] [CrossRef]
- Waterlow, J. Protein turnover with special reference to man. Q. J. Exp. Physiol. 1984, 69, 409–438. [Google Scholar] [CrossRef]
- Moberg, G.P.; Mench, J.A. The Biology of Animal Stress: Basic Principles and Implications for Animal Welfare, 1st ed.; CABI Pub: Wallingford, CT, USA, 2000; pp. 309–377. [Google Scholar] [CrossRef]
- Morelle, K.; Podgórski, T.; Prévot, C.; Keuling, O.; Lehaire, F.; Lejeune, P. Towards understanding wild boar Sus scrofa movement: A synthetic movement ecology approach. Mamm. Rev. 2015, 45, 15–29. [Google Scholar] [CrossRef]
- Tolon, V.; Dray, S.; Loison, A.; Zeileis, A.; Fischer, C.; Baubet, E. Responding to spatial and temporal variations in predation risk: Space use of a game species in a changing landscape of fear. Can. J. Zool. 2009, 87, 1129–1137. [Google Scholar] [CrossRef]
- Le Floc’h, N.; Wessels, A.; Corrent, E.; Wu, G.; Bosi, P. The relevance of functional amino acids to support the health of growing pigs. Anim. Feed Sci. Technol. 2018, 245, 104–116. [Google Scholar] [CrossRef]
- Bouyeh, M. Effect of excess lysine and methionine on immune system and performance of broilers. Ann. Biol. Res 2012, 3, 3218–3224. [Google Scholar]
- Yao, H.; Yuzuriha, T.; Koga, H.; Fukuda, K.; Endo, K.; Matsumoto, T.; Kato, A.; Uchino, A.; Ezaki, T.; Ibayashi, S. Decreased plasma tryptophan associated with deep white matter lesions in elderly subjects. J. Neurol. Neurosurg. Psychiatry 1999, 66, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Capozzalo, M.; Kim, J.; Htoo, J.; de Lange, C.; Mullan, B.; Hansen, C.; Resink, J.; Stumbles, P.; Hampson, D.; Pluske, J. An increased ratio of dietary tryptophan to lysine improves feed efficiency and elevates plasma tryptophan and kynurenine in the absence of antimicrobials and regardless of infection with enterotoxigenic Escherichia coli in weaned pigs. J. Anim. Sci. 2012, 90, 191–193. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yin, Y.-L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef]
- Mateo, R.D.; Wu, G.; Bazer, F.W.; Park, J.C.; Shinzato, I.; Kim, S.W. Dietary L-arginine supplementation enhances the reproductive performance of gilts. J. Nutr. 2007, 137, 652–656. [Google Scholar] [CrossRef]
- Xia, M.; Pan, Y.; Guo, L.; Wei, X.X.; Xiong, J.; Wang, L.; Peng, J.; Wang, C.; Peng, J.; Wei, H. Effect of gestation dietary methionine/lysine ratio on placental angiogenesis and reproductive performance of sows. J. Anim. Sci. 2019, 97, 3487–3497. [Google Scholar] [CrossRef]
- Li, H.; Wan, H.; Mercier, Y.; Zhang, X.; Wu, C.; Wu, X.; Tang, L.; Che, L.; Lin, Y.; Xu, S. Changes in plasma amino acid profiles, growth performance and intestinal antioxidant capacity of piglets following increased consumption of methionine as its hydroxy analogue. Br. J. Nutr. 2014, 112, 855–867. [Google Scholar] [CrossRef]
- Che, L.; Hu, L.; Wu, C.; Xu, Q.; Zhou, Q.; Peng, X.; Fang, Z.; Lin, Y.; Xu, S.; Feng, B. Effects of increased energy and amino acid intake in late gestation on reproductive performance, milk composition, metabolic, and redox status of sows. J. Anim. Sci. 2019, 97, 2914–2926. [Google Scholar] [CrossRef]
- Wang, C.X.; Chen, F.; Zhang, W.F.; Zhang, S.H.; Shi, K.; Song, H.Q.; Wang, Y.J.; Kim, S.W.; Guan, W.T. Leucine promotes the growth of fetal pigs by increasing protein synthesis through the mTOR signaling pathway in longissimus dorsi muscle at late gestation. J. Agric. Food Chem. 2018, 66, 3840–3849. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Publication | Animal | Amino Acid | Dose Range | Protein Accretion Variable | Physiological Response Variable |
|---|---|---|---|---|---|
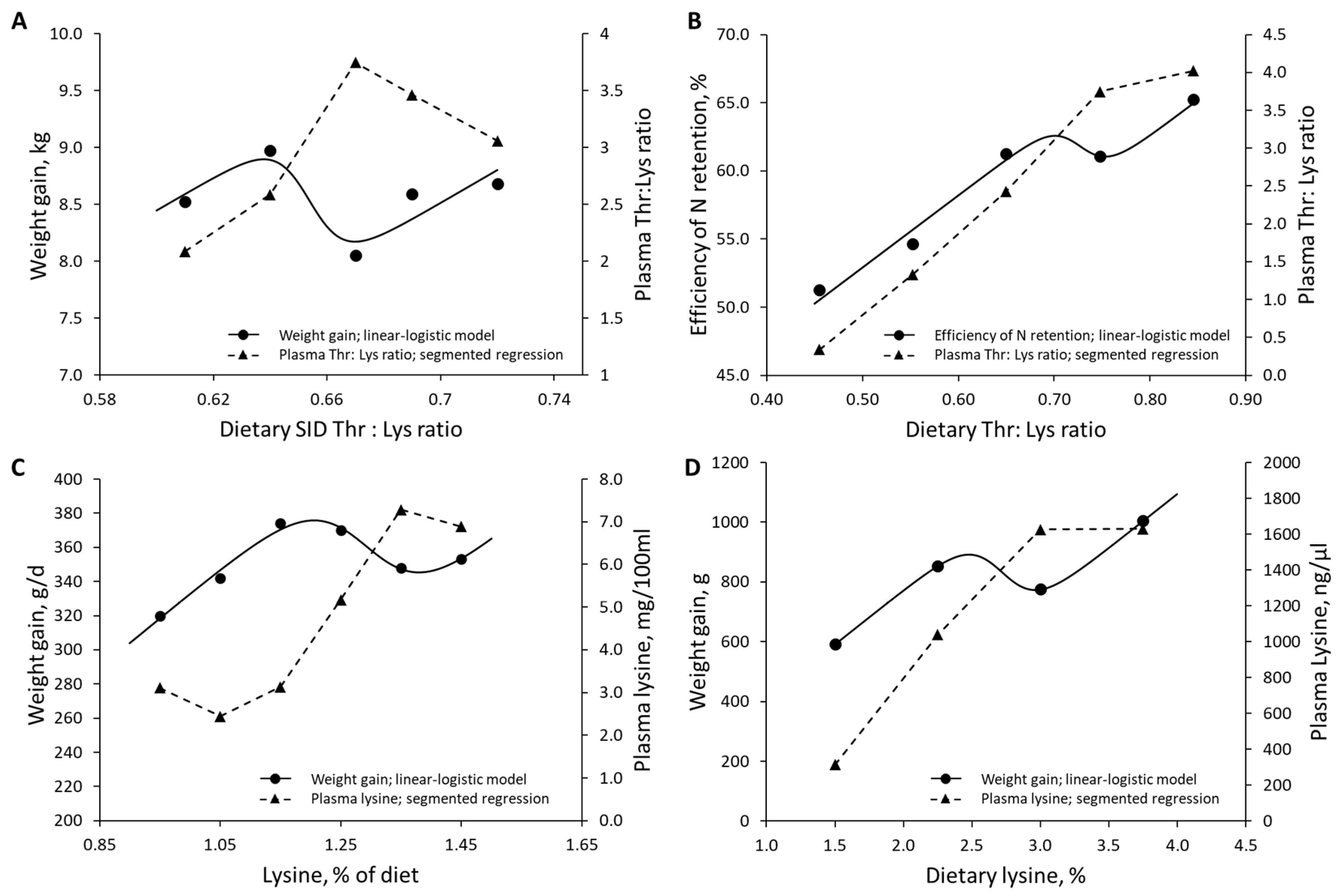
| Lewis et al. (1980) [55] | Growing pigs | Lys | 0.95–1.45% of the diet | Weight gain | Plasma AA |
| Lara et al. (2006) [53] | Holstein cows | RP Met | 0–24 g/d | Body weight | Milk production |
| Hosseini et al. (2011) [56] | Broiler hens | Met | 0.2%–0.45% of the diet | Weight gain | IgG & IgM |
| Jayaraman et al. (2015) [57] | Growing pigs | Lys | 61–72 SID Thr: Lys ratio | Weight gain | Plasma AA |
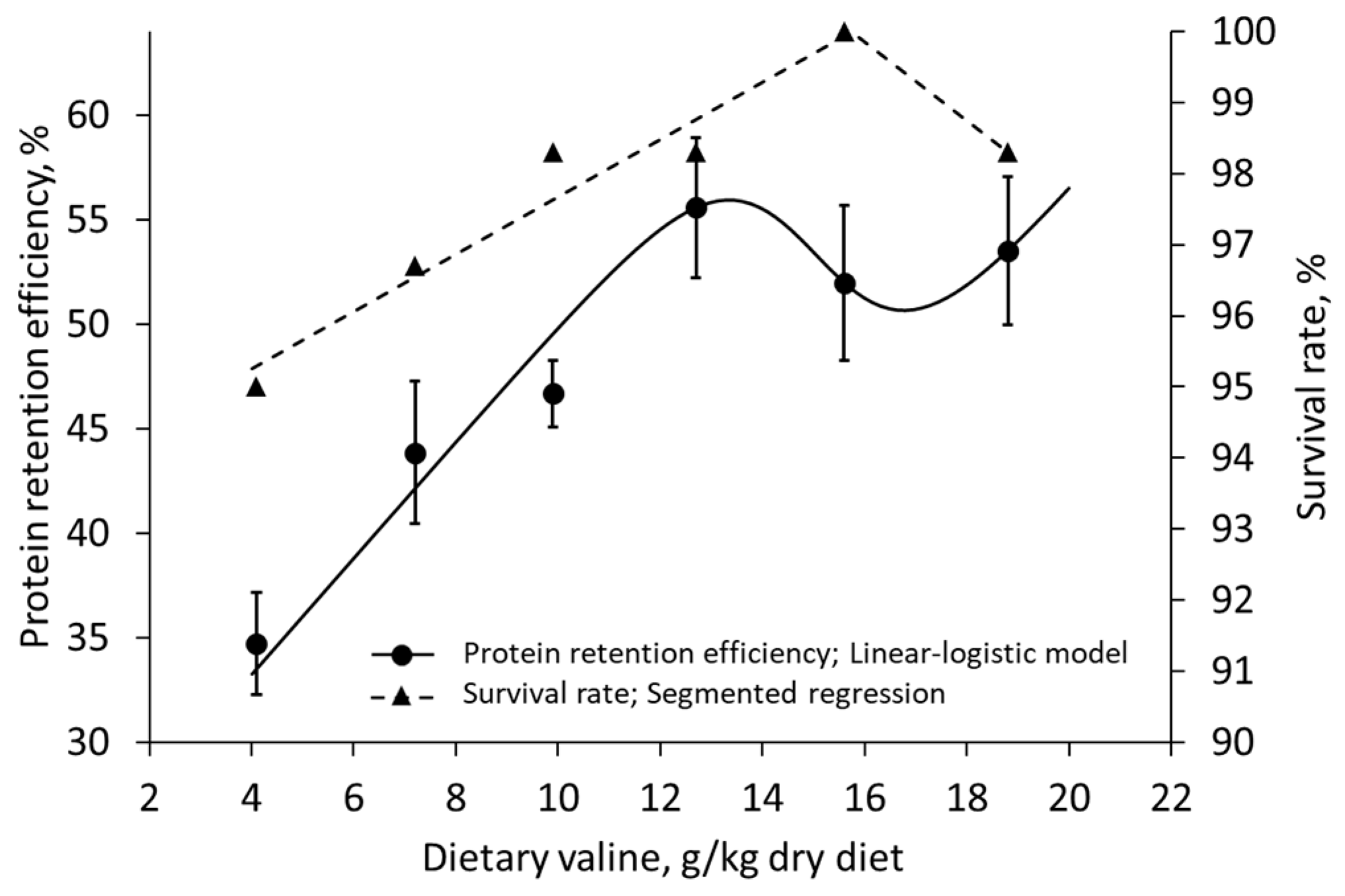
| Xiao et al. (2018) [58] | Nile tilapia | Val | 4.1–18.8 g/kg dried diet | Protein retention | Survival rate |
| Remus et al. (2019) [59] | Growing pigs | Thr | 46–85 Thr to Lys ratio | N retention | Plasma AA |
| Ramirez-Camba et al. (2020) [54] | Pregnant sows | SID Lys | 5.4–12.3 g/d | N retention | Total piglets born |
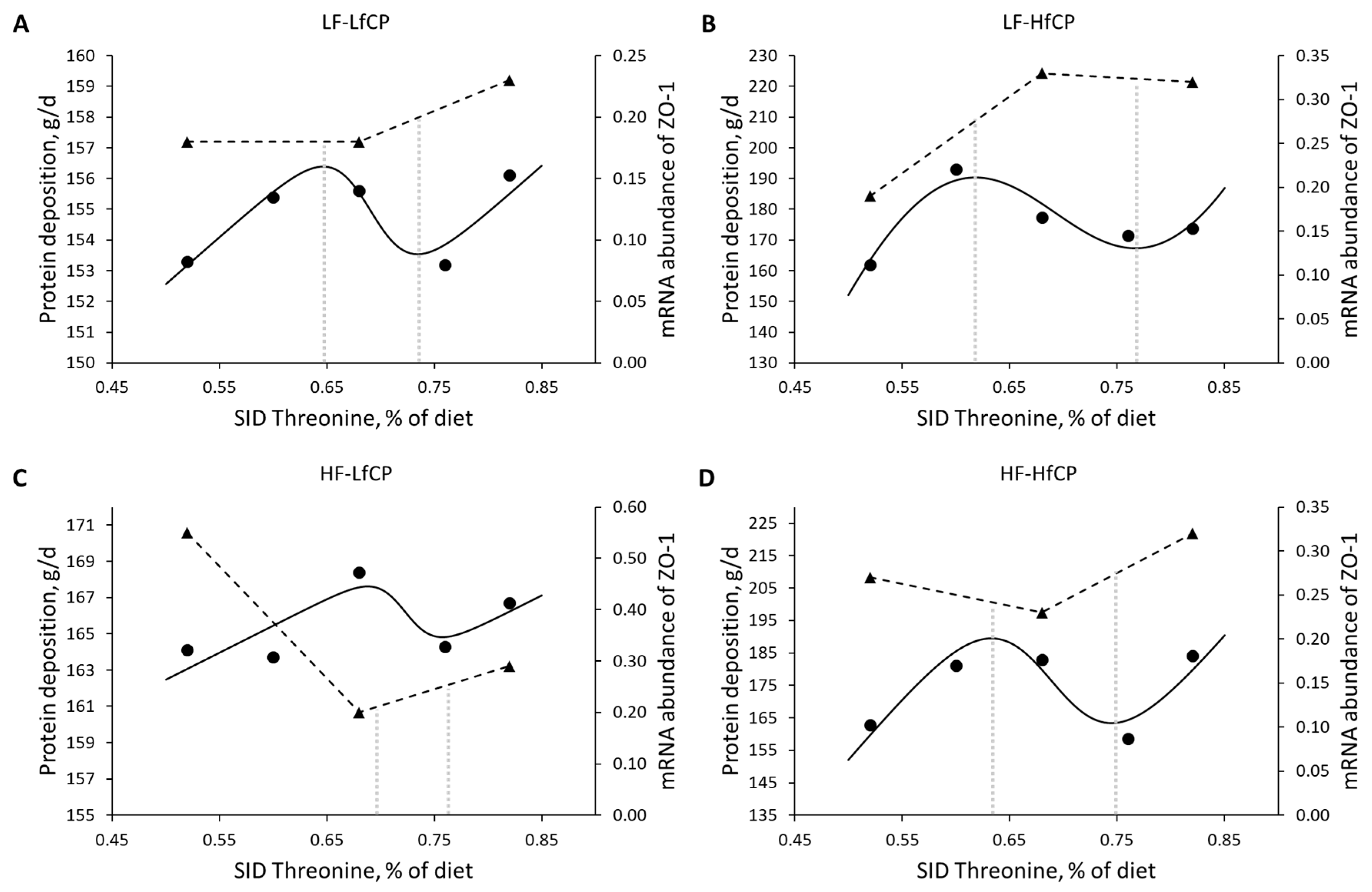
| Wellington et al. (2020) [60] | Growing pigs | SID Thr | 0.52–0.82% of the diet | Protein retention | ZO-1 1 |
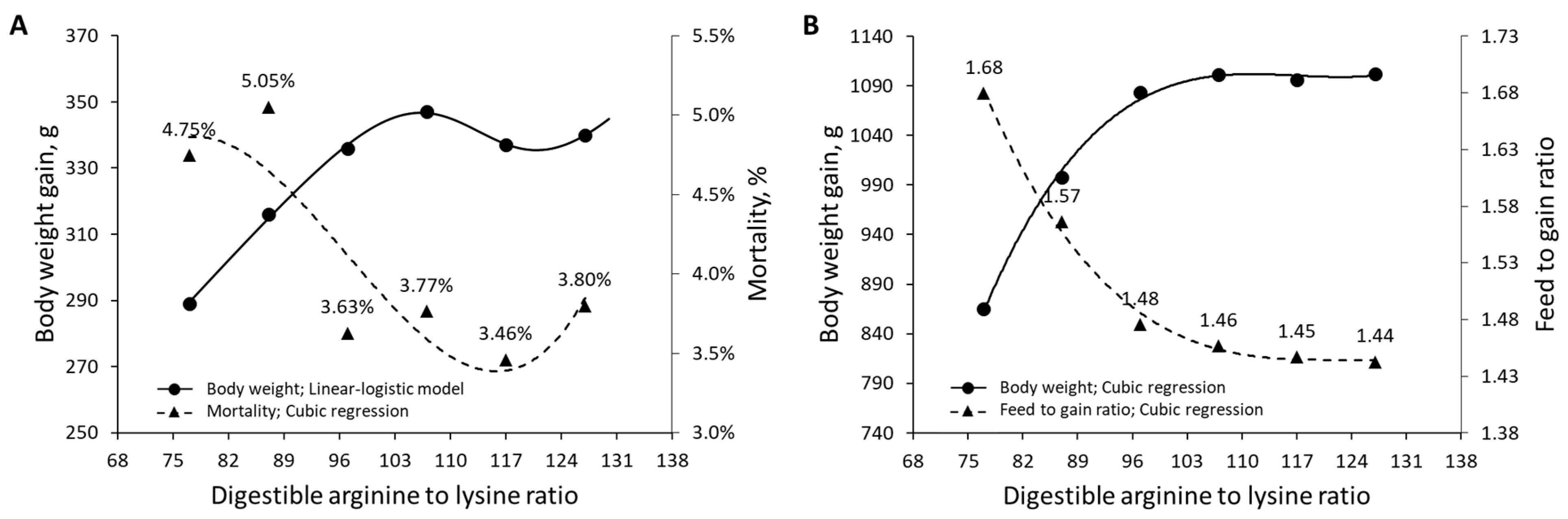
| Corzo et al. (2021) [26] | Male broilers | Arg | 77–127 dArg: Lys ratio | Body weight | Survival rate |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramirez-Camba, C.D.; Levesque, C.L. The Linear-Logistic Model: A Novel Paradigm for Estimating Dietary Amino Acid Requirements. Animals 2023, 13, 1708. https://doi.org/10.3390/ani13101708
Ramirez-Camba CD, Levesque CL. The Linear-Logistic Model: A Novel Paradigm for Estimating Dietary Amino Acid Requirements. Animals. 2023; 13(10):1708. https://doi.org/10.3390/ani13101708
Chicago/Turabian StyleRamirez-Camba, Christian D., and Crystal L. Levesque. 2023. "The Linear-Logistic Model: A Novel Paradigm for Estimating Dietary Amino Acid Requirements" Animals 13, no. 10: 1708. https://doi.org/10.3390/ani13101708