
Foraging and Food Selection in a Desert Rodent: Diet Shifts of the Sandy Inland Mouse between Population Booms and Busts


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Direct Observations of Foraging
2.3. Collection of Stomach Material and Diet Analysis
2.4. Statistical Analyses
3. Results
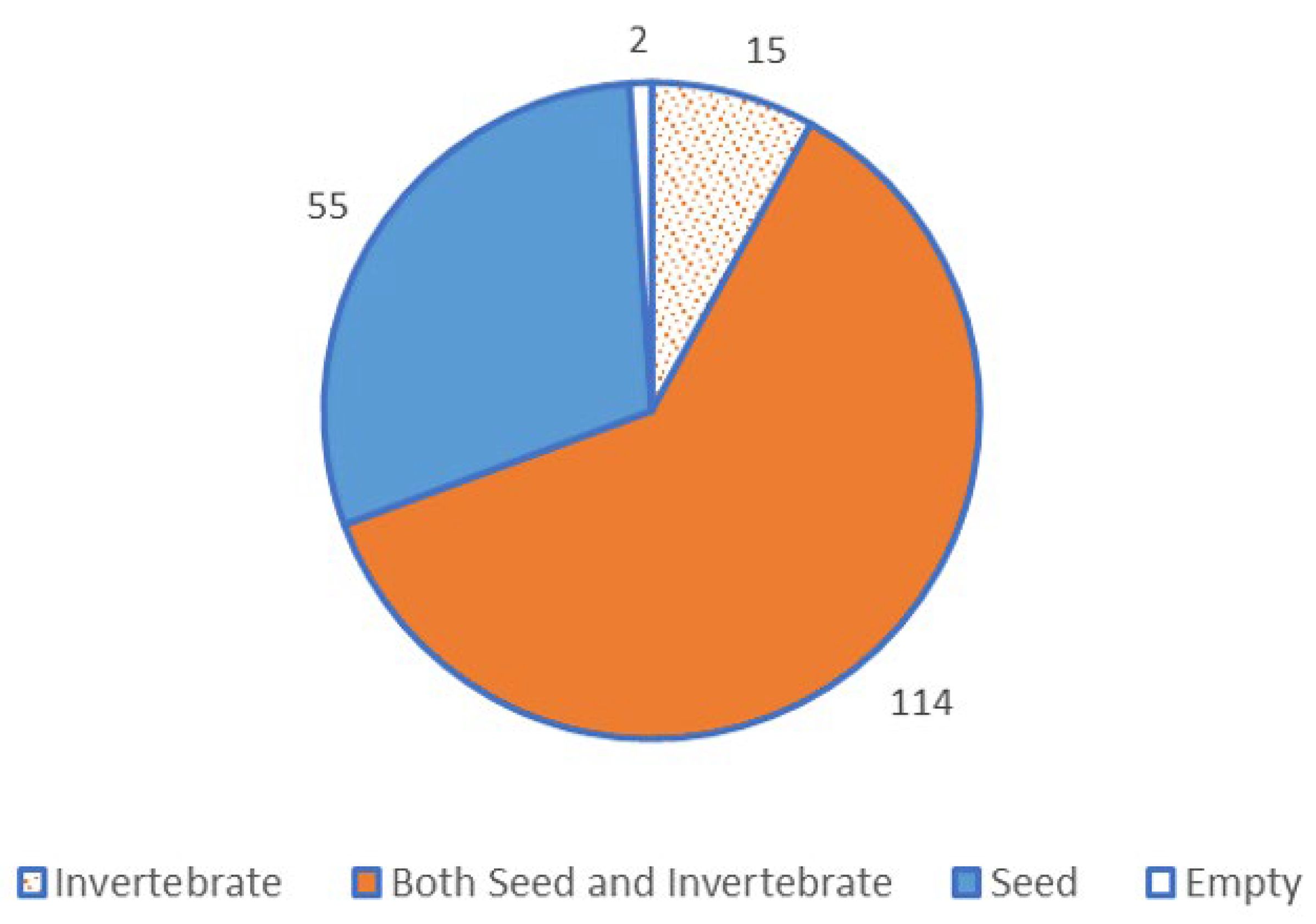
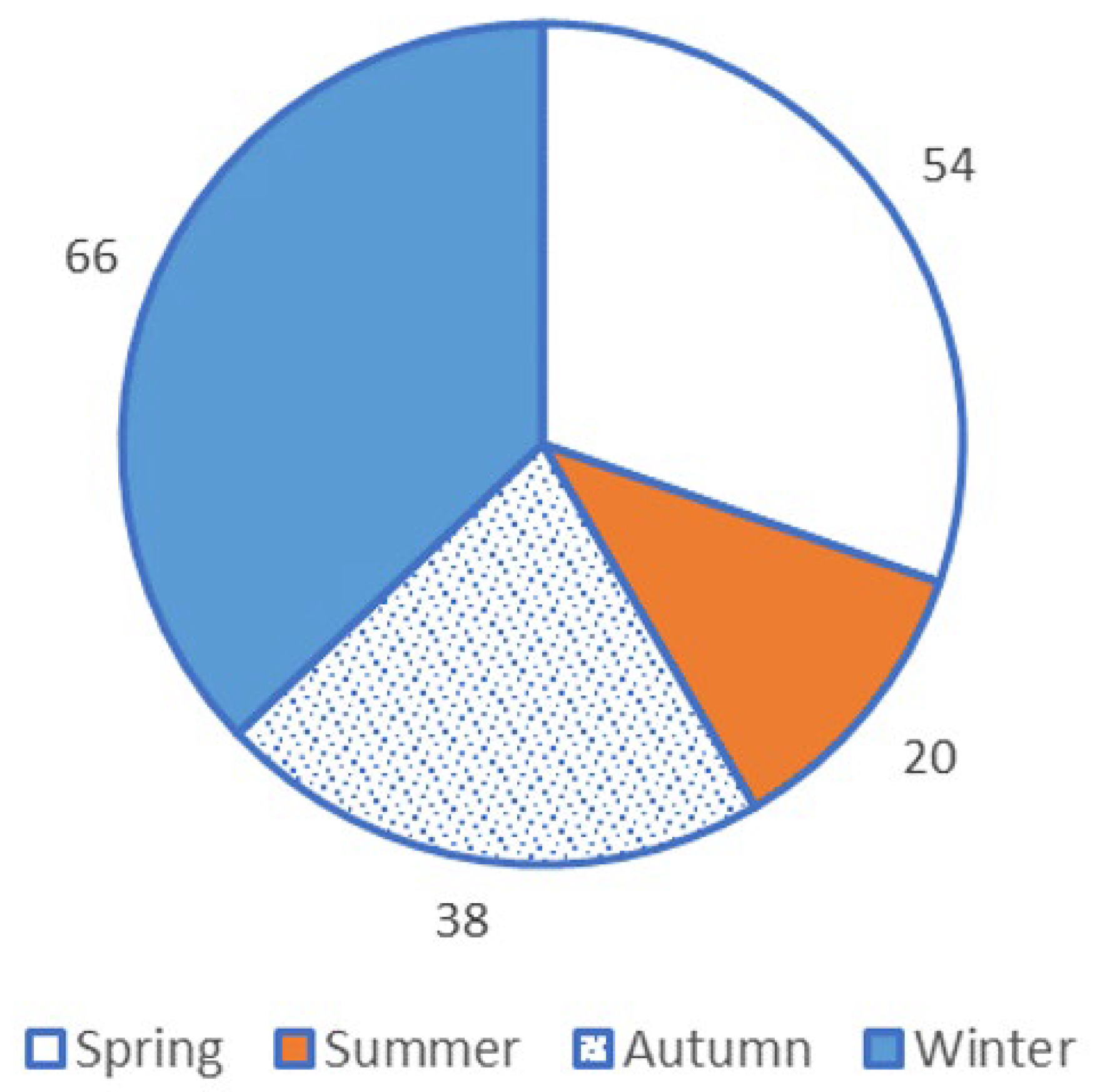
3.1. Direct Observations
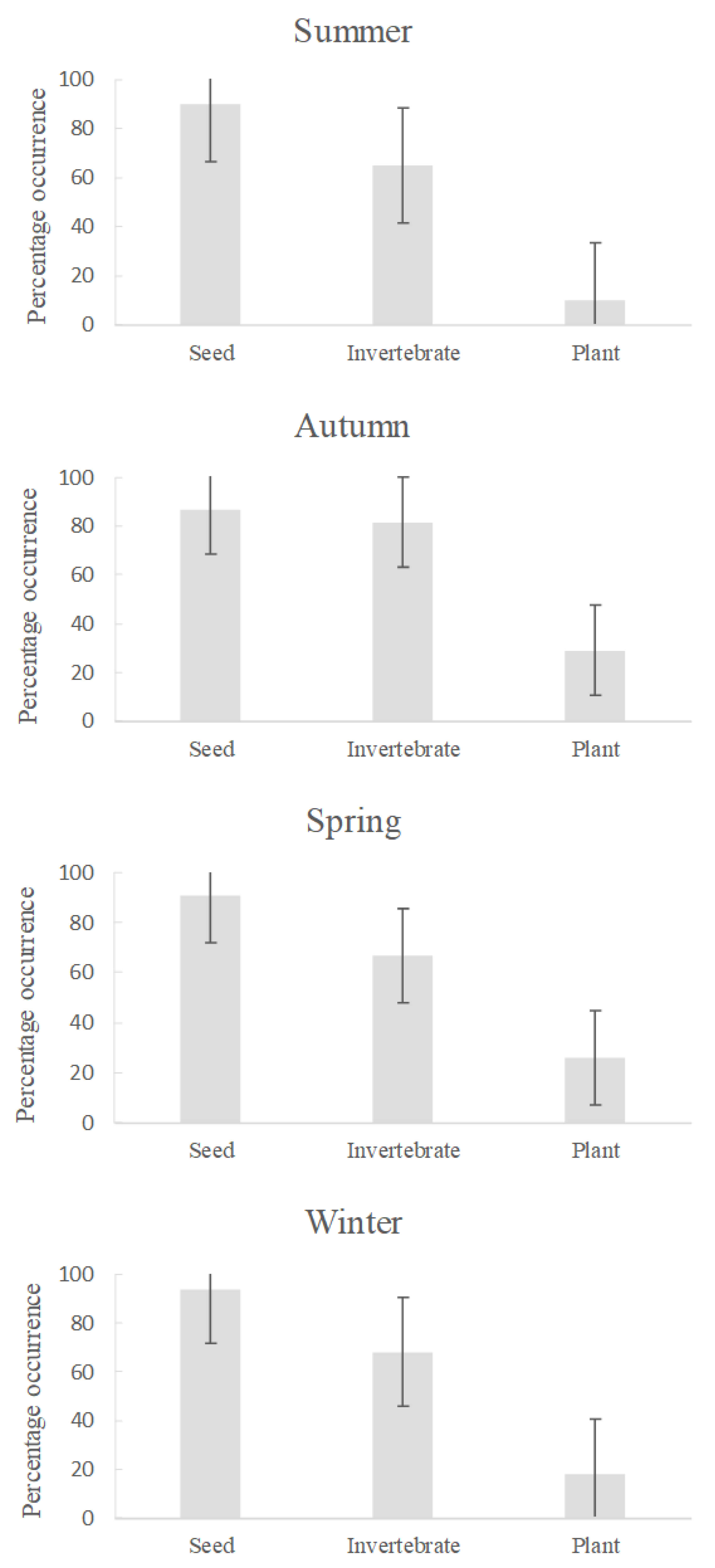
3.2. Stomach Content Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arregoitia, L.D.V.; D’Elía, G. Classifying rodent diets for comparative research. Mamm. Rev. 2021, 51, 51–65. [Google Scholar] [CrossRef]
- Brown, J.H.; Reichman, O.J.; Davidson, D.W. Granivory in desert ecosystems. Ann. Rev. Ecol. Syst. 1979, 10, 201–227. [Google Scholar] [CrossRef]
- Guo, Q.; Thompson, D.B.; Valone, T.J.; Brown, J.H. The effects of vertebrate granivores and folivores on plant community structure in the Chihuahuan Desert. Oikos 1995, 73, 251–259. [Google Scholar] [CrossRef]
- Chen, T.C.; Valone, T.J. Rodent granivory strengthens relationships between seed size and plant abundance in a desert annual community. J. Veg. Sci. 2017, 28, 808–814. [Google Scholar] [CrossRef]
- Mares, M.A. South American mammal zoogeography: Evidence from convergent evolution in desert rodents. Proc. Nat. Acad. Sci. USA 1975, 72, 1702–1706. [Google Scholar] [CrossRef] [PubMed]
- Mares, M.A. Convergent evolution among desert rodents: A global perspective. Bull. Carnegie Mus. Nat. Hist. 1980, 16, 1–51. [Google Scholar] [CrossRef]
- Mares, M.A. Desert rodents, seed consumption, and convergence. BioScience 1993, 43, 372–379. [Google Scholar] [CrossRef]
- Campos, C.M.; Ojeda, R.; Monge, S.; Dacar, M. Utilization of food resources by small and medium-sized mammals in the Monte Desert biome, Argentina. Austral Ecol. 2001, 26, 435–438. [Google Scholar] [CrossRef]
- Prevedello, J.A.; Vieira, M.V.; Vieira, E.M.; Dickman, C.R. The importance of food supply in high-productivity ecosystems: Short-term experimental tests with small rodents. Austral Ecol. 2017, 42, 176–186. [Google Scholar] [CrossRef]
- Fiedler, L.A. Rodent pest problems in Africa. In Rodent Pest Management; Prakash, I., Ed.; CRC Press: Boca Raton, FL, USA, 1988; pp. 35–65. [Google Scholar] [CrossRef]
- Granjon, L.; Cosson, J.-F.; Quesseveur, E.; Sicard, B. Population dynamics of the multimammate rat Mastomys huberti in an annually flooded agricultural region of central Mali. J. Mammal. 2005, 86, 997–1008. [Google Scholar] [CrossRef]
- Kerley, G.I.H.; Whitford, W.G. Desert-dwelling small mammals as granivores: Intercontinental variations. Aust. J. Zool. 1994, 42, 543–555. [Google Scholar] [CrossRef]
- Cockburn, A. Diet and habitat preference of the silky desert mouse, Pseudomys apodemoides (Rodentia). Aust. Wildl. Res. 1981, 8, 475–497. [Google Scholar] [CrossRef]
- Murray, B.R.; Dickman, C.R.; Watts, C.H.S.; Morton, S.R. The dietary ecology of Australian desert rodents. Wildl. Res. 1999, 26, 421–437. [Google Scholar] [CrossRef]
- Murray, B.R.; Dickman, C.R. Granivory and microhabitat use in Australian desert rodents: Are seeds important? Oecologia 1994, 99, 216–225. [Google Scholar] [CrossRef]
- Murray, B.R.; Dickman, C.R. Food preferences and seed selection in two species of Australian desert rodents. Wildl. Res. 1994, 21, 647–655. [Google Scholar] [CrossRef]
- Predavec, M. Population dynamics and environmental changes during natural irruptions of Australian desert rodents. Wildl. Res. 1994, 21, 569–581. [Google Scholar] [CrossRef]
- Predavec, M. Variable energy demands in Pseudomys hermannsburgensis: Possible ecological consequences. Aust. J. Zool. 1997, 45, 85–94. [Google Scholar] [CrossRef]
- Watts, C.H.S.; Aslin, H.J. The Rodents of Australia; Angus & Robertson: Sydney, Australia, 1981. [Google Scholar]
- Breed, B.; Ford, F. Native Mice and Rats; CSIRO Publishing: Melbourne, Australia, 2007. [Google Scholar]
- Watts, C.H.S. The foods eaten by some Australian desert rodents. South Aust. Nat. 1970, 44, 71–74. [Google Scholar]
- Watts, C.H.S.; Morton, S.R. Notes on the diets of Mus musculus and Pseudomys hermannsburgensis (Rodentia: Muridae) in western Queensland. Aust. Mammal. 1983, 6, 81–82. [Google Scholar] [CrossRef]
- Morton, S.R. Diversity of desert-dwelling mammals: A comparison of Australia and North America. J. Mammal. 1979, 60, 253–264. [Google Scholar] [CrossRef]
- Ricci, S. Population Dynamics and Trophic Ecology of Two Species of Australian Desert Rodents. Ph.D. Thesis, University of Sydney, Sydney, Australia, 2003. [Google Scholar]
- Morton, S.R.; Stafford Smith, D.M.; Dickman, C.R.; Dunkerley, D.L.; Friedel, M.H.; McAllister, R.R.J.; Reid, J.R.W.; Roshier, D.A.; Smith, M.A.; Walsh, F.J.; et al. A fresh framework for the ecology of arid Australia. J. Arid Environ. 2011, 75, 313–329. [Google Scholar] [CrossRef]
- Ward, A.L. Stomach content and fecal analysis: Methods of forage identification. In Range and Wildlife Habitat Evaluation: A Research Symposium; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1970; pp. 146–158. [Google Scholar]
- Brand, M.R. A method to correct for differential digestibility in fecal analysis. Am. Midl. Nat. 1978, 100, 228–232. [Google Scholar] [CrossRef]
- Wardle, G.M.; Greenville, A.C.; Frank, A.S.K.; Tischler, M.; Emery, N.J.; Dickman, C.R. Ecosystem risk assessment of Georgina gidgee woodlands in central Australia. Austral Ecol. 2015, 40, 444–459. [Google Scholar] [CrossRef]
- Dickman, C.R.; Greenville, A.C.; Beh, C.-L.; Tamayo, B.; Wardle, G.M. Social organization and movements of desert rodents during population “booms” and “busts” in central Australia. J. Mammal. 2010, 91, 798–810. [Google Scholar] [CrossRef]
- Greenville, A.C.; Dickman, C.R. The ecology of Lerista labialis (Scincidae) in the Simpson Desert: Reproduction and diet. J. Arid Environ. 2005, 60, 611–625. [Google Scholar] [CrossRef]
- Bureau of Meteorology. Climate Statistics for Australian Locations: Bedourie. Available online: http://www.bom.gov.au/climate/averages/tables/cw_038000.shtml (accessed on 5 July 2022).
- Dickman, C.R.; Mahon, P.S.; Masters, P.; Gibson, D.F. Long-term dynamics of rodent populations in arid Australia: The influence of rainfall. Wildl. Res. 1999, 26, 389–403. [Google Scholar] [CrossRef]
- Dickman, C.R.; Wardle, G.M.; Foulkes, J.; de Preu, N. Desert complex environments. In Biodiversity and Environmental Change: Monitoring, Challenges and Direction; Lindenmayer, D., Burns, E., Thurgate, N., Lowe, A., Eds.; CSIRO Publishing: Melbourne, Australia, 2014; pp. 379–438. [Google Scholar]
- Greenville, A.C.; Wardle, G.M.; Dickman, C.R. Extreme climatic events drive mammal irruptions: Regression analysis of 100-year trends in desert rainfall and temperature. Ecol. Evol. 2012, 2, 2645–2658. [Google Scholar] [CrossRef]
- Greenville, A.C.; Wardle, G.M.; Dickman, C.R. Extreme rainfall events predict irruptions of rat plagues in central Australia. Austral Ecol. 2013, 38, 754–764. [Google Scholar] [CrossRef]
- Greenville, A.C.; Wardle, G.M.; Nguyen, V.; Dickman, C.R. Population dynamics of desert mammals: Similarities and contrasts within a multispecies assemblage. Ecosphere 2016, 7, 1343. [Google Scholar] [CrossRef]
- Dickman, C.R.; Greenville, A.C.; Tamayo, B.; Wardle, G.M. Spatial dynamics of small mammals in central Australian desert habitats: The role of drought refugia. J. Mammal. 2011, 92, 1193–1209. [Google Scholar] [CrossRef]
- Lemen, C.; Freeman, P.W. Tracking animals with fluorescent pigments: A new technique. J. Mammal. 1985, 66, 134–136. [Google Scholar] [CrossRef]
- Boonstra, R.; Craine, I.T.M. Natal nest location and small mammal tracking with a spool and line technique. Can. J. Zool. 1986, 64, 1034–1036. [Google Scholar] [CrossRef]
- Nikbakht, N.; Diamond, M.E. Conserved visual capacity of rats under red light. eLife 2021, 10, e66429. [Google Scholar] [CrossRef] [PubMed]
- Beh, C.-L. Population Dynamics of Australian Desert Rodents: The Effects of Food and Water. Ph.D. Thesis, University of Sydney, Sydney, Australia, 2011. [Google Scholar]
- Gregory, D.G. The Role of Woodlands as Micro-refugia in a Desert Sand Dune Environment. Honours Thesis, University of Sydney, Sydney, Australia, 2008. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 12 November 2022).
- Cloudsley-Thompson, J.L. Ecophysiology of Desert Arthropods and Reptiles; Springer: Berlin, Germany, 1991. [Google Scholar]
- Whitford, W.G. Ecology of Desert Systems; Academic Press: London, UK, 2002. [Google Scholar]
- Brown, J.H. Species diversity of seed-eating desert rodents in sand dune habitats. Ecology 1973, 54, 775–787. [Google Scholar] [CrossRef]
- Reichman, O.J. Relation of desert rodent diets to available resources. J. Mammal. 1975, 56, 731–751. [Google Scholar] [CrossRef]
- Predavec, M. Food limitation in Australian desert rodents: Experiments using supplementary feeding. Oikos 2000, 91, 512–522. [Google Scholar] [CrossRef]
- Gordon, C.E.; Letnic, M. Functional extinction of a desert rodent: Implications for seed fate and vegetation dynamics. Ecography 2016, 39, 815–824. [Google Scholar] [CrossRef]
- Gibb, H.; Wardle, G.M.; Greenville, A.C.; Grossman, B.F.; Dickman, C.R. Top-down response to spatial variation in productivity and bottom-up response to temporal variation in productivity in a long-term study of desert ants. Biol. Lett. 2022, 18, 20220314. [Google Scholar] [CrossRef] [PubMed]
- Kwok, A.B.; Wardle, G.M.; Greenville, A.C.; Dickman, C.R. Long-term patterns of invertebrate abundance and relationships to environmental factors in arid Australia. Austral Ecol. 2016, 41, 480–491. [Google Scholar] [CrossRef]
- Thompson, S.N.; Simpson, S.J. Nutrition. In Encyclopedia of Insects, 2nd ed.; Resh, V.H., Cardé, R.T., Eds.; Elsevier: Amsterdam, The Netherlands, 2009; pp. 715–720. [Google Scholar]
- Bleicher, S.S.; Marko, H.; Morin, D.J.; Teemu, K.; Ylönen, H. Balancing food, activity and the dangers of sunlit nights. Behav. Ecol. Sociobiol. 2019, 73, 95. [Google Scholar] [CrossRef]
- Kotler, B.P.; Dickman, C.R.; Brown, J.S. The effects of water on patch use by two Simpson Desert granivores (Corvus coronoides and Pseudomys hermannsburgensis). Aust. J. Ecol. 1998, 23, 574–578. [Google Scholar] [CrossRef]
- Yip, S.J.S. Dietary Shifts in Vertebrates: Factors Affecting Seed Selection in Desert Rodents. Ph.D. Thesis, University of Sydney, Sydney, Australia, 2023. [Google Scholar]
- Jackson, S.M.; Dickman, C.R.; Old, J.; Hulst, F.; Gleen, W. Rodents. In Australian Mammals: Biology and Captive Management, 2nd ed.; Jackson, S.M., Ed.; CSIRO Publishing: Melbourne, Australia, 2023; in press. [Google Scholar]
- Vander Wall, S.B. How plants manipulate the scatter-hoarding behaviour of seed-dispersing animals. Proc. Roy. Soc. B 2010, 365, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Morton, S.R. Determinants of diversity in animal communities of arid Australia. In Species Diversity in Ecological Communities: Historical and Geographical Perspectives; Ricklefs, R.E., Schluter, D., Eds.; University of Chicago Press: Chicago, IL, USA, 1993; pp. 159–169. [Google Scholar]
- Kelt, D.A.; Brown, J.H.; Heske, E.J.; Marquet, P.A.; Morton, S.R.; Reid, J.R.; Rogovin, K.A.; Shenbrot, G. Community structure of desert small mammals: Comparisons across four continents. Ecology 1996, 77, 746–761. [Google Scholar] [CrossRef]
- Vernes, K.; Cooper, T.; Green, S. Seasonal fungal diets of small mammals in an Australian temperate forest ecosystem. Fungal Ecol. 2015, 18, 107–114. [Google Scholar] [CrossRef]
- Elliott, T.F.; Truong, C.; Jackson, S.M.; Zúñiga, C.L.; Trappe, J.M.; Vernes, K. Mammalian mycophagy: A global review of ecosystem interactions between mammals and fungi. Fungal Syst. Evol. 2022, 9, 99–159. [Google Scholar] [CrossRef]
- Rodrigues, N.T.; Saranholi, B.H.; Angeloni, T.A.; Pasqualotto, N.; Chiarello, A.G.; Galetti, P.M., Jr. DNA mini-barcoding of leporids using noninvasive fecal DNA samples and its significance for monitoring an invasive species. Ecol. Evol. 2020, 10, 5219–5225. [Google Scholar] [CrossRef]
- Murray, B.R.; Dickman, C.R. Factors affecting selection of native seeds in two species of Australian desert rodents. J. Arid Env. 1997, 35, 517–525. [Google Scholar] [CrossRef]
- Dickman, C.R. The Biology and Management of Native Rodents of the Arid Zone in New South Wales; NSW National Parks and Wildlife Service: Parramatta, Australia, 1993; Volume 12, pp. 1–149.
- Morton, S.R.; Brown, J.H.; Kelt, D.A.; Reid, J.R.W. Comparisons of community structure among small mammals of North American and Australian deserts. Aust. J. Zool. 1994, 42, 501–525. [Google Scholar] [CrossRef]
- Weaver, H.J.; Smales, L.R. Parasite assemblages of Australian species of Pseudomys (Rodentia: Muridae: Murinae). J. Parasitol. 2012, 98, 30–35. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Food Type | Spring | Summer | Autumn | Winter | Boom | Bust |
|---|---|---|---|---|---|---|
| Seed | 14 (74) | 12 (71) | 21 (81) | 18 (90) | 27 (79) | 38 (79) |
| Invertebrate | 3 (16) | 5 (29) | 2 (8) | 1 (5) | 5 (15) | 6 (13) |
| Green plant | 2 (10) | 0 (0) | 3 (11) | 1 (5) | 2 (6) | 4 (8) |
| Food Type | Spring | Summer | Autumn | Winter | Boom | Bust |
|---|---|---|---|---|---|---|
| Seed | 49 (49) | 18 (55) | 33 (44) | 62 (52) | 65 (58) | 97 (45) |
| Invertebrate | 36 (36) | 13 (39) | 31 (41) | 45 (38) | 33 (29) | 92 (43) |
| Green plant | 14 (14) | 2 (6) | 11 (15) | 12 (10) | 14 (13) | 25 (12) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yip, S.J.S.; Dickman, C.R. Foraging and Food Selection in a Desert Rodent: Diet Shifts of the Sandy Inland Mouse between Population Booms and Busts. Animals 2023, 13, 1702. https://doi.org/10.3390/ani13101702
Yip SJS, Dickman CR. Foraging and Food Selection in a Desert Rodent: Diet Shifts of the Sandy Inland Mouse between Population Booms and Busts. Animals. 2023; 13(10):1702. https://doi.org/10.3390/ani13101702
Chicago/Turabian StyleYip, Stephanie J. S., and Christopher R. Dickman. 2023. "Foraging and Food Selection in a Desert Rodent: Diet Shifts of the Sandy Inland Mouse between Population Booms and Busts" Animals 13, no. 10: 1702. https://doi.org/10.3390/ani13101702