
Abundance and Distribution of Microplastics in Invertebrate and Fish Species and Sediment Samples along the German Wadden Sea Coastline


Abstract
:Simple Summary
Abstract
1. Introduction
- Analyzing the nature and extent of the occurrence of microplastics in biota in the North Sea coastal waters of Lower Saxony.
- Evaluation of species–specific differences and possible correlations with species- and individual-specific parameters.
- Evaluation of spatial differences and influencing factors of the occurring microplastic concentrations in biota between selected study stations along the coast of Lower Saxony.
- Provision of recommendations for a future monitoring strategy on microplastics in biota for the Lower Saxony coastal waters, with regard to the selection of indicator organisms, station selection, monitoring frequency, and analysis methodology.
2. Materials and Methods
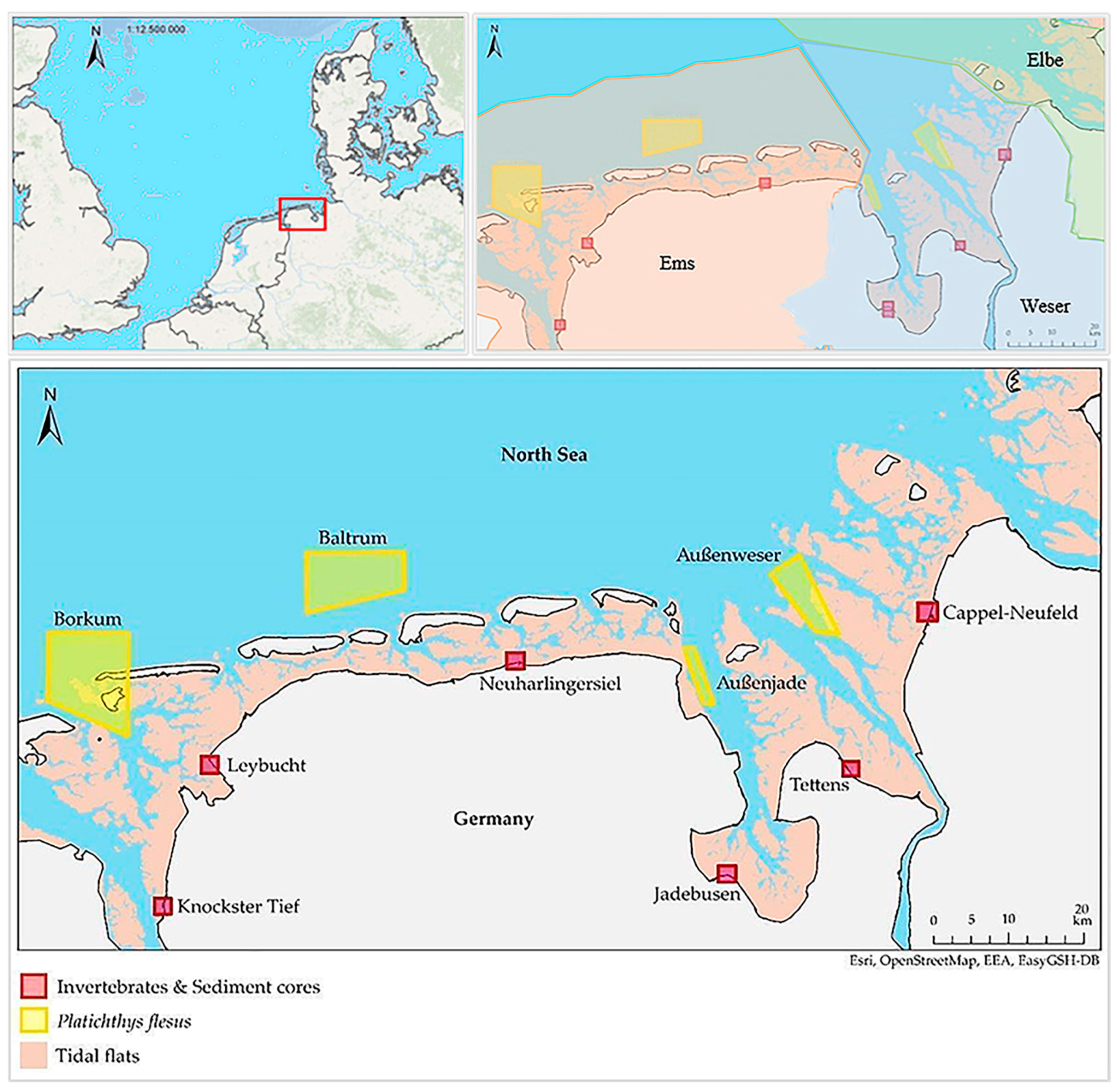
2.1. Study Area and Sampling
2.2. Laboratory Analysis—Sample Treatment
2.3. Identification of Particle Characteristics and Polymer Composition
2.4. Statistical Analysis
2.5. Background Contamination
3. Results
3.1. Procedural Blanks
3.2. Invertebrates and Fish
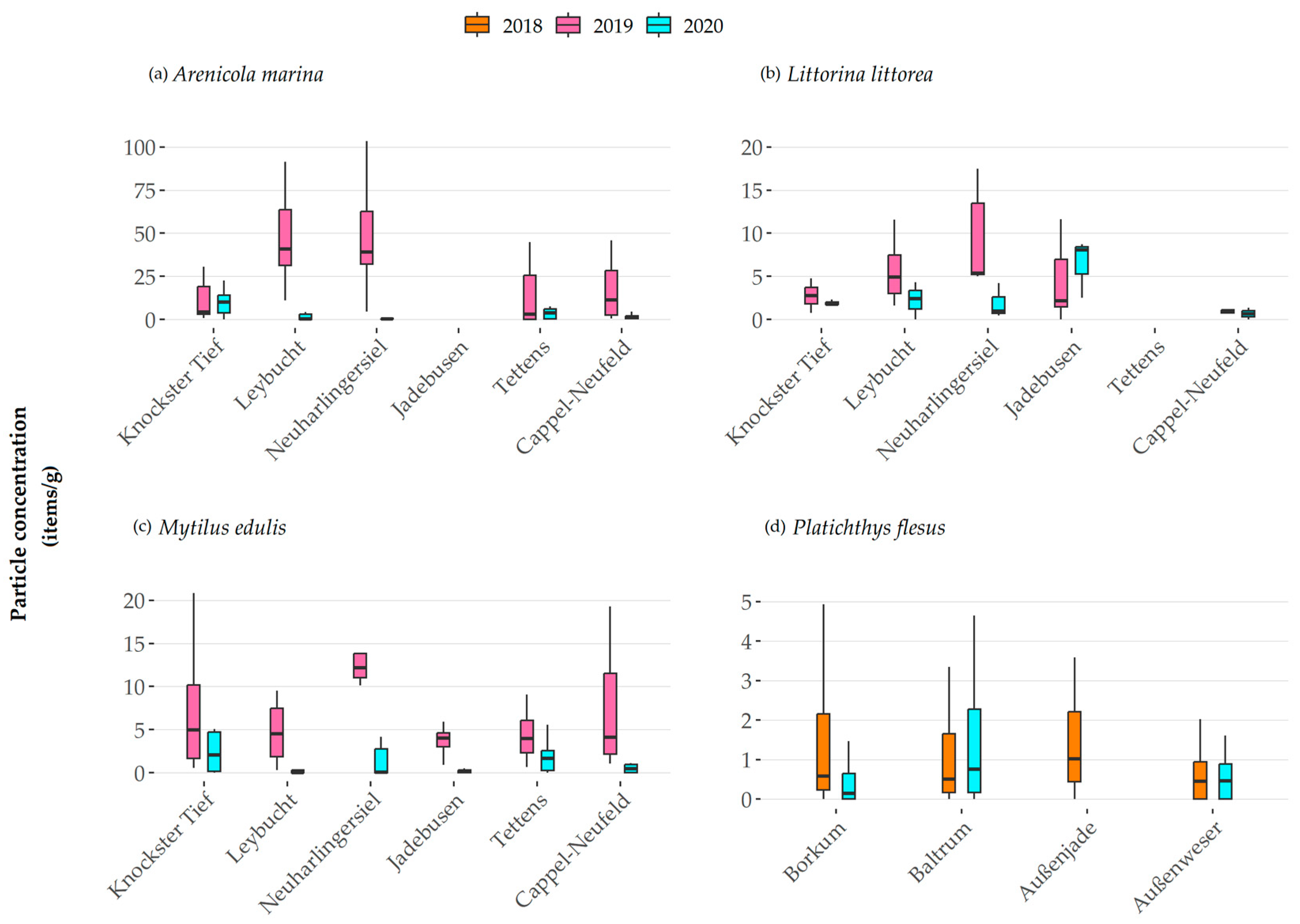
3.2.1. Microplastic Distribution and Concentrations
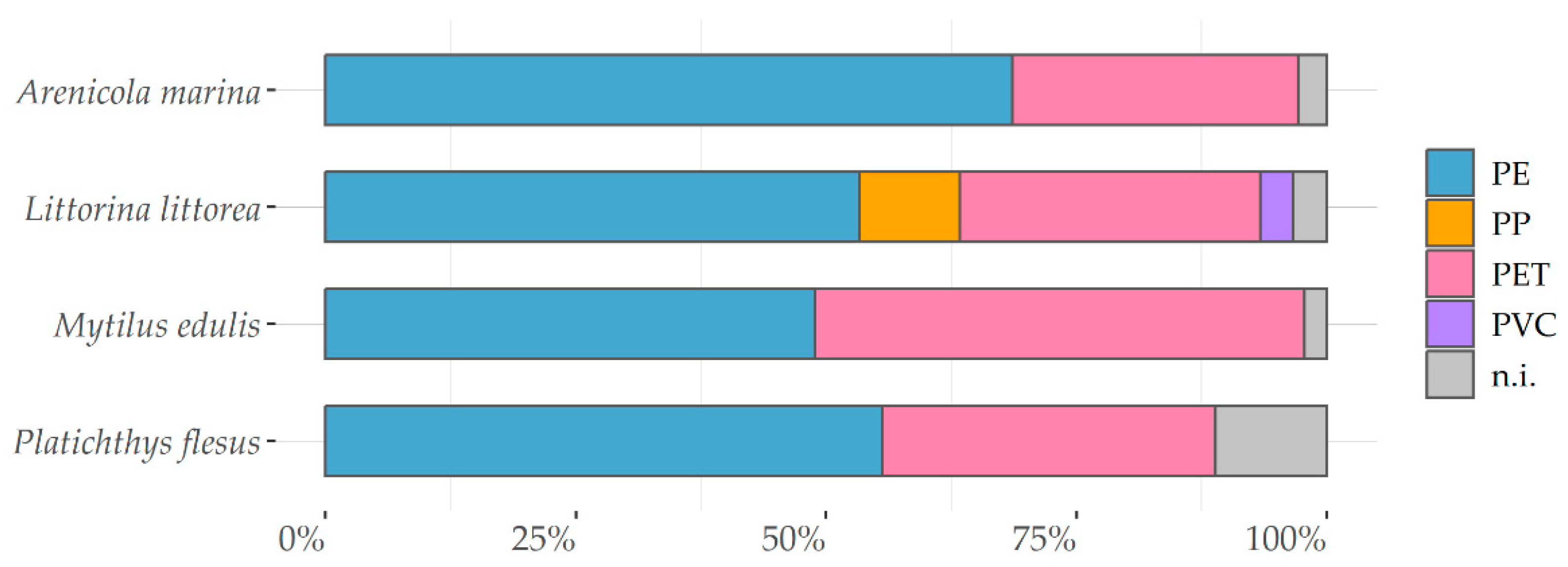
3.2.2. Particle Characteristics
3.3. Sediment Cores
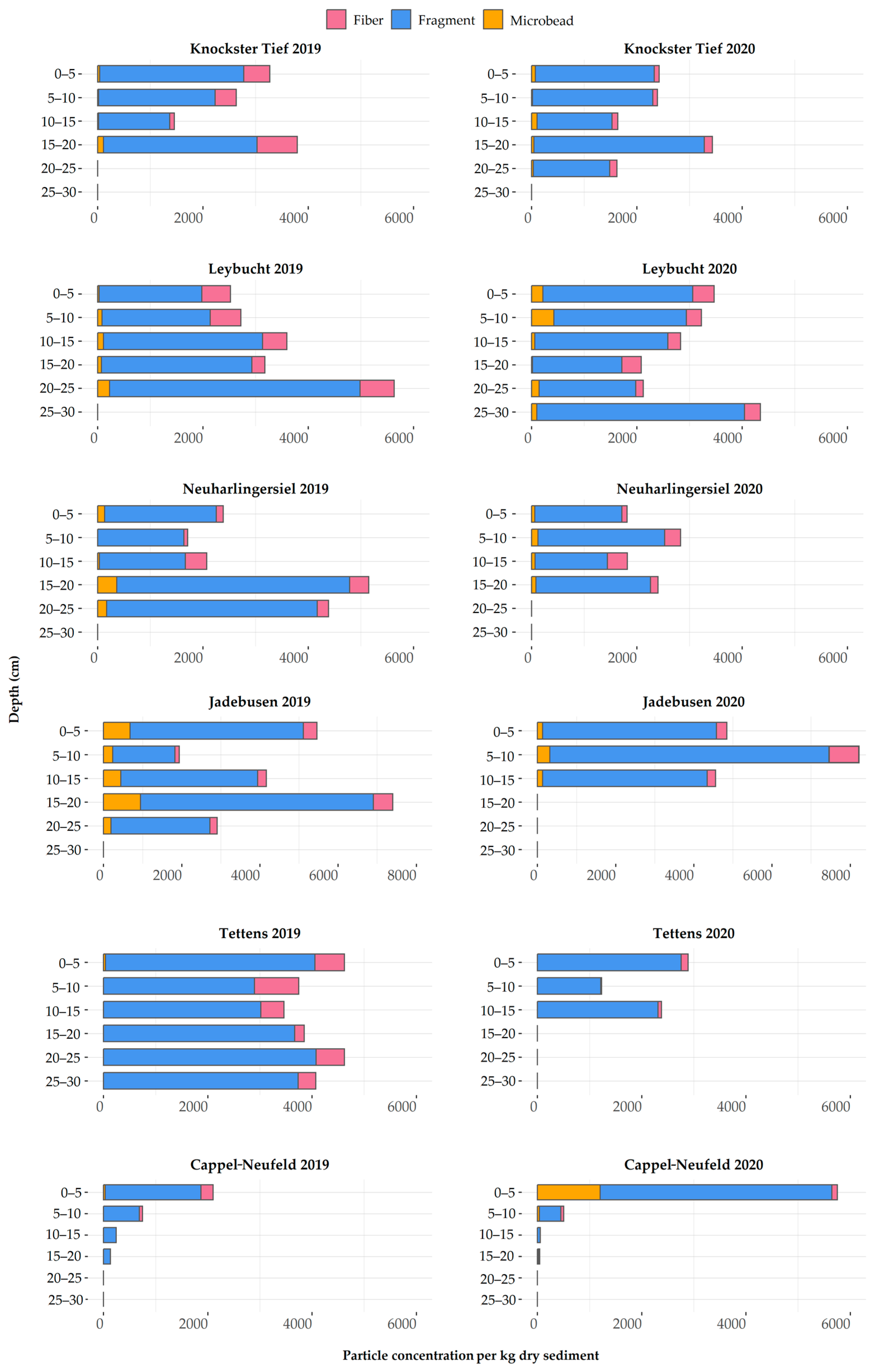
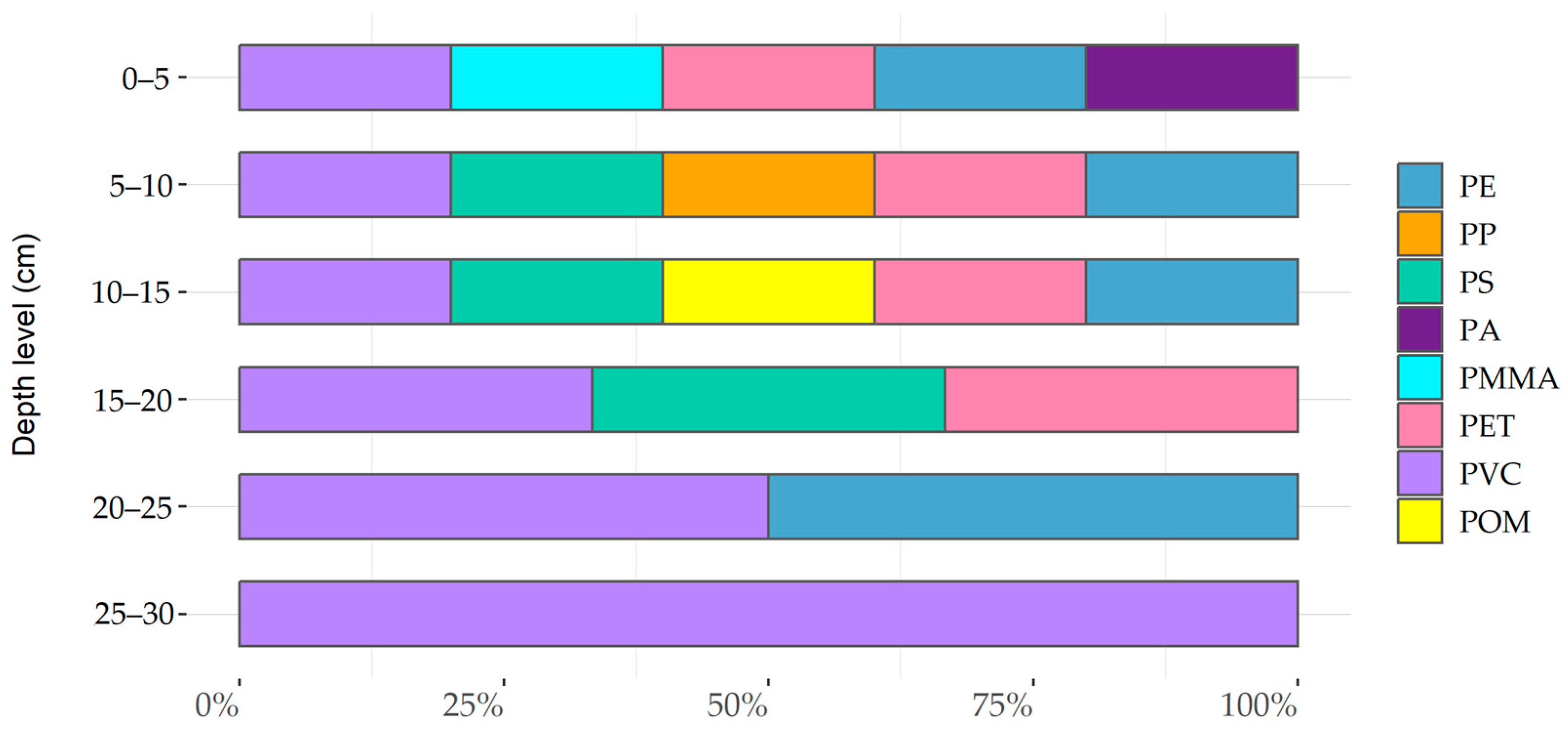
3.3.1. Microplastic Distribution and Concentrations
3.3.2. Particle Characteristics
4. Discussion
4.1. MP Concentrations in Invertebrates and Fish
4.2. MP Concentrations in Sediment Cores
4.3. Suitability of Investigated Species for MP-Biomonitoring
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kershaw, P.J.; Turra, A.; Galgani, F. (Eds.) GESAMP Guidelines or the Monitoring and Assessment of Plastic Litter and Microplastics in the Ocean; (IMO/FAO/UNESCOIOC/UNIDO/WMO/IAEA/UN/UNEP/UNDP/ISA Joint Group of Experts on the Scientific Aspects of Marine Environmental Protection); Reports and Studies. GESAMP; IMO: London, UK, 2019; Volume 99. [Google Scholar]
- HELCOM Monitoring Manual. 2021. Available online: https://helcom.fi/action-areas/monitoring-andassessment/monitoring-manual/ (accessed on 8 April 2023).
- MSFD Technical Group on Marine Litter TG Litter. 2021. Available online: https://mcc.jrc.ec.europa.eu/main/dev.py?N=41&O=434&titre_chap=TG (accessed on 8 April 2023).
- OSPAR Commission Monitoring & Assessing Marine Litter. 2021. Available online: https://www.ospar.org/work-areas/eiha/marine-litter/assessment-of-marine-litter (accessed on 8 April 2023).
- Arthur, C.; Baker, J.; Bamford, H. NOAA Technical Memorandum NOS-OR&R-30. In Proceedings of the International Research Workshop on the Occurrence, Effects, and Fate of Microplastic Marine Debris, Tacoma, WA, USA, 9–11 September 2008; pp. 9–11. [Google Scholar]
- SAPEA Science Advice for Policy by European Academies. A Scientific Perspective on Microplastics in Nature and Society. 2019. Available online: https://sapea.info/topic/microplastics/ (accessed on 8 April 2023).
- GESAMP. Sources, Fate and Effects of Microplastics in the Marine Environment. Part Two of a Global Assessment (IMO/FAO/UNESCO-IOC/UNIDO/WMO/IAEA/UN/UNEP/UNDP, 2016); Reports and Studies. GESAMP; IMO: London, UK, 2016. [Google Scholar]
- Hartmann, N.B.; Hüffer, T.; Thompson, R.C.; Hassellöv, M.; Verschoor, A.; Daugaard, A.E.; Rist, S.; Karlsson, T.; Brennholt, N.; Cole, M.; et al. Are We Speaking the Same Language? Recommendations for a Definition and Categorization Framework for Plastic Debris. Environ. Sci. Technol. 2019, 53, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Koelmans, A.A.; Redondo-Hasselerharm, P.E.; Nor, N.H.M.; De Ruijter, V.N.; Mintenig, S.M.; Kooi, M. Risk Assessment of Microplastic Particles. Nat. Rev. Mater. 2022, 7, 138–152. [Google Scholar] [CrossRef]
- Rochman, C.M. Microplastics Research—From Sink to Source. Science 2018, 360, 28–29. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.C.; Olsen, Y.; Mitchell, R.P.; Davis, A.; Rowland, S.J.; John, A.W.G.; McGonigle, D.; Russell, A.E. Lost at Sea: Where Is All the Plastic? Science 2004, 304, 838. [Google Scholar] [CrossRef] [PubMed]
- Lebreton, L.C.M.; Van Der Zwet, J.; Damsteeg, J.-W.; Slat, B.; Andrady, A.; Reisser, J. River Plastic Emissions to the World’s Oceans. Nat. Commun. 2017, 8, 15611. [Google Scholar] [CrossRef] [PubMed]
- Blettler, M.C.M.; Garello, N.; Ginon, L.; Abrial, E.; Espinola, L.A.; Wantzen, K.M. Massive Plastic Pollution in a Mega-River of a Developing Country: Sediment Deposition and Ingestion by Fish (Prochilodus lineatus). Environ. Pollut. 2019, 255, 113348. [Google Scholar] [CrossRef] [PubMed]
- Cole, M.; Lindeque, P.; Halsband, C.; Galloway, T.S. Microplastics as Contaminants in the Marine Environment: A Review. Mar. Pollut. Bull. 2011, 62, 2588–2597. [Google Scholar] [CrossRef]
- Lusher, A.L.; Welden, N.A.; Sobral, P.; Cole, M. Sampling, Isolating and Identifying Microplastics Ingested by Fish and Invertebrates. Anal. Methods 2017, 9, 1346–1360. [Google Scholar] [CrossRef]
- Hermsen, E.; Mintenig, S.M.; Besseling, E.; Koelmans, A.A. Quality Criteria for the Analysis of Microplastic in Biota Samples: A Critical Review. Environ. Sci. Technol. 2018, 52, 10230–10240. [Google Scholar] [CrossRef]
- Wesch, C.; Bredimus, K.; Paulus, M.; Klein, R. Towards the Suitable Monitoring of Ingestion of Microplastics by Marine Biota: A Review. Environ. Pollut. 2016, 218, 1200–1208. [Google Scholar] [CrossRef]
- Bessa, F.; Frias, J.; Knögel, T.; Lusher, A.; Andrade, J.; Antunes, J.C.; Sobral, P.; Pagter, E.; Nash, R.; O’Connor, I.; et al. Harmonized Protocol for Monitoring Microplastics in Biota; JPI-Ocean. BASEMAN Project; 2019. [Google Scholar] [CrossRef]
- Bråte, I.L.N.; Hurley, R.; Iversen, K.; Beyer, J.; Thomas, K.V.; Steindal, C.C.; Green, N.W.; Olsen, M.; Lusher, A. Mytilus Spp. as Sentinels for Monitoring Microplastic Pollution in Norwegian Coastal Waters: A Qualitative and Quantitative Study. Environ. Pollut. 2018, 243, 383–393. [Google Scholar] [CrossRef]
- Beer, S.; Garm, A.; Huwer, B.; Dierking, J.; Nielsen, T.G. No Increase in Marine Microplastic Concentration over the Last Three Decades—A Case Study from the Baltic Sea. Sci. Total Environ. 2018, 621, 1272–1279. [Google Scholar] [CrossRef] [PubMed]
- Białowąs, M.; Jonko-Sobuś, K.; Pawlak, J.; Polak-Juszczak, L.; Dąbrowska, A.; Urban-Malinga, B. Plastic in Digestive Tracts and Gills of Cod and Herring from the Baltic Sea. Sci. Total Evriesnviron. 2022, 822, 153333. [Google Scholar] [CrossRef] [PubMed]
- Bråte, I.L.N.; Eidsvoll, D.P.; Steindal, C.C.; Thomas, K.V. Plastic Ingestion by Atlantic Cod (Gadus Morhua) from the Norwegian Coast. Mar. Pollut. Bull. 2016, 112, 105–110. [Google Scholar] [CrossRef] [PubMed]
- de Vries, A.N.; Govoni, D.; Árnason, S.H.; Carlsson, P. Microplastic Ingestion by Fish: Body Size, Condition Factor and Gut Fullness Are Not Related to the Amount of Plastics Consumed. Mar. Pollut. Bull. 2020, 151, 110827. [Google Scholar] [CrossRef]
- Fischer, E.K. Distribution of Microplastics in Marine Species of the Wadden Sea along the Coastline of Schleswig-Holstein, Germany. 2019. Available online: https://www.geo.uni-hamburg.de/geographie/abteilungen/zentrale-einrichtungen/labor/pdf-dokumente/pdf-projekte/fischer-distribution-of-microplastics-in-marine-species-of-the-wadden-sea-germany-2019.pdf (accessed on 8 April 2023).
- Hermsen, E.; Pompe, R.; Besseling, E.; Koelmans, A.A. Detection of Low Numbers of Microplastics in North Sea Fish Using Strict Quality Assurance Criteria. Mar. Pollut. Bull. 2017, 122, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, T.M.; Vethaak, A.D.; Almroth, B.C.; Ariese, F.; van Velzen, M.; Hassellöv, M.; Leslie, H.A. Screening for Microplastics in Sediment, Water, Marine Invertebrates and Fish: Method Development and Microplastic Accumulation. Mar. Pollut. Bull. 2017, 122, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Kühn, S.; van Franeker, J.A.; O’Donoghue, A.M.; Swiers, A.; Starkenburg, M.; van Werven, B.; Foekema, E.; Hermsen, E.; Egelkraut-Holtus, M.; Lindeboom, H. Details of Plastic Ingestion and Fibre Contamination in North Sea Fishes. Environ. Pollut. 2020, 257, 113569. [Google Scholar] [CrossRef] [PubMed]
- Lenz, R.; Enders, K.; Beer, S.; Sørensen, T.K.; Stedmon, C.A. Analysis of Microplastic in the Stomachs of Herring and Cod from the North Sea and Baltic Sea; DTU Aqua National Institute of Aquatic Resources; Technical University of Denmark: Lyngby, Denmark, 2016. [Google Scholar] [CrossRef]
- Lusher, A.L.; McHugh, M.; Thompson, R.C. Occurrence of Microplastics in the Gastrointestinal Tract of Pelagic and Demersal Fish from the English Channel. Mar. Pollut. Bull. 2013, 67, 94–99. [Google Scholar] [CrossRef]
- McGoran, A.R.; Clark, P.F.; Morritt, D. Presence of Microplastic in the Digestive Tracts of European Flounder, Platichthys Flesus, and European Smelt, Osmerus Eperlanus, from the River Thames. Environ. Pollut. 2017, 220, 744–751. [Google Scholar] [CrossRef]
- Murphy, F.; Russell, M.; Ewins, C.; Quinn, B. The Uptake of Macroplastic & Microplastic by Demersal & Pelagic Fish in the Northeast Atlantic around Scotland. Mar. Pollut. Bull. 2017, 122, 353–359. [Google Scholar] [CrossRef]
- Rummel, C.D.; Löder, M.G.J.; Fricke, N.F.; Lang, T.; Griebhermseler, E.-M.; Janke, M.; Gerdts, G. Plastic Ingestion by Pelagic and Demersal Fish from the North Sea and Baltic Sea. Mar. Pollut. Bull. 2016, 102, 134–141. [Google Scholar] [CrossRef]
- Bessa, F.; Barría, P.; Neto, J.M.; Frias, J.P.G.L.; Otero, V.; Sobral, P.; Marques, J.C. Occurrence of Microplastics in Commercial Fish from a Natural Estuarine Environment. Mar. Pollut. Bull. 2018, 128, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Kazour, M.; Jemaa, S.; El Rakwe, M.; Duflos, G.; Hermabassiere, L.; Dehaut, A.; Le Bihanic, F.; Cachot, J.; Cornille, V.; Rabhi, K.; et al. Juvenile Fish Caging as a Tool for Assessing Microplastics Contamination in Estuarine Fish Nursery Grounds. Env. Sci. Pollut. Res. 2020, 27, 3548–3559. [Google Scholar] [CrossRef]
- Catarino, A.I.; Macchia, V.; Sanderson, W.G.; Thompson, R.C.; Henry, T.B. Low Levels of Microplastics (MP) in Wild Mussels Indicate That MP Ingestion by Humans Is Minimal Compared to Exposure via Household Fibres Fallout during a Meal. Environ. Pollut. 2018, 237, 675–684. [Google Scholar] [CrossRef]
- Halbach, M.; Vogel, M.; Tammen, J.K.; Rüdel, H.; Koschorreck, J.; Scholz-Böttcher, B.M. 30 Years Trends of Microplastic Pollution: Mass-Quantitative Analysis of Archived Mussel Samples from the North and Baltic Seas. Sci. Total Environ. 2022, 826, 154179. [Google Scholar] [CrossRef]
- Leslie, H.A.; Brandsma, S.H.; van Velzen, M.J.M.; Vethaak, A.D. Microplastics En Route: Field Measurements in the Dutch River Delta and Amsterdam Canals, Wastewater Treatment Plants, North Sea Sediments and Biota. Environ. Int. 2017, 101, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Green, C.; Reynolds, A.; Shi, H.; Rotchell, J.M. Microplastics in Mussels Sampled from Coastal Waters and Supermarkets in the United Kingdom. Environ. Pollut. 2018, 241, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Van Cauwenberghe, L.; Claessens, M.; Vandegehuchte, M.B.; Janssen, C.R. Microplastics Are Taken up by Mussels (Mytilus Edulis) and Lugworms (Arenicola Marina) Living in Natural Habitats. Environ. Pollut. 2015, 199, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Phuong, N.N.; Poirier, L.; Pham, Q.T.; Lagarde, F.; Zalouk-Vergnoux, A. Factors Influencing the Microplastic Contamination of Bivalves from the French Atlantic Coast: Location, Season and/or Mode of Life? Mar. Pollut. Bull. 2018, 129, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Doyle, D.; Gammell, M.; Frias, J.; Griffin, G.; Nash, R. Low Levels of Microplastics Recorded from the Common Periwinkle, Littorina Littorea on the West Coast of Ireland. Mar. Pollut. Bull. 2019, 149, 110645. [Google Scholar] [CrossRef]
- Bour, A.; Avio, C.G.; Gorbi, S.; Regoli, F.; Hylland, K. Presence of Microplastics in Benthic and Epibenthic Organisms: Influence of Habitat, Feeding Mode and Trophic Level. Environ. Pollut. 2018, 243, 1217–1225. [Google Scholar] [CrossRef]
- Muller-Karanassos, C.; Turner, A.; Arundel, W.; Vance, T.; Lindeque, P.K.; Cole, M. Antifouling Paint Particles in Intertidal Estuarine Sediments from Southwest England and Their Ingestion by the Harbour Ragworm, Hediste Diversicolor. Environ. Pollut. 2019, 249, 163–170. [Google Scholar] [CrossRef]
- European Commission Directorate-General for Environment. Commission Decision (EU) 2017/848 of 17 May 2017 Laying down Criteria and Methodological Standards on Good Environmental Status of Marine Waters and Specifications and Standardised Methods for Monitoring and Assessment, and Repealing Decision 2010/477/EU. 2017. Available online: http://data.europa.eu/eli/dec/2017/848/oj (accessed on 12 May 2023).
- OSPAR Commission. OSPAR’s Second Regional Action Plan for the Prevention and Management of Marine Litter in the North-East Atlantic (RAP ML 2) (2022–2030) (OSPAR Agreement 2022-05). 2023. Available online: https://www.un.org/depts/los//general_assembly/contributions_2016/OSPAR_Contribution_to_ICP_on_marine_debris.pdf (accessed on 12 May 2023).
- HELCOM. Terms of Reference for the HELCOM Expert Group on Marine Litter (EG Marine Litter) 2022–2024. 2022. Available online: https://helcom.fi/wp-content/uploads/2021/12/Terms-of-Reference-for-the-HELCOM-Expert-Group-on-Marine-Litter-2022-2024.pdf (accessed on 12 May 2023).
- Beyer, J.; Green, N.W.; Brooks, S.; Allan, I.J.; Ruus, A.; Gomes, T.; Bråte, I.L.N.; Schøyen, M. Blue Mussels (Mytilus edulis spp.) as Sentinel Organisms in Coastal Pollution Monitoring: A Review. Mar. Environ. Res. 2017, 130, 338–365. [Google Scholar] [CrossRef]
- Li, J.; Lusher, A.L.; Rotchell, J.M.; Deudero, S.; Turra, A.; Bråte, I.L.N.; Sun, C.; Shahadat Hossain, M.; Li, Q.; Kolandhasamy, P.; et al. Using Mussel as a Global Bioindicator of Coastal Microplastic Pollution. Environ. Pollut. 2019, 244, 522–533. [Google Scholar] [CrossRef] [PubMed]
- Kazour, M.; Amara, R. Is Blue Mussel Caging an Efficient Method for Monitoring Environmental Microplastics Pollution? Sci. Total Environ. 2020, 710, 135649. [Google Scholar] [CrossRef] [PubMed]
- Nationalpark Wattenmeer. Willkommen im Nationalpark. 2022. Available online: https://www.nationalpark-wattenmeer.de/wp-content/uploads/2022/02/fb_willkommen_nationalpark_nlpv_202202_web.pdf (accessed on 8 April 2023).
- Behrends, B.; Dittmann, S.; Liebezeit, G.; Kaiser, M.; Knoke, V.; Petri, G.; Rahmel, J.; Roy, M.; Scheiffarth, G.; Wilhelmsen, U. Gesamtsynthese Ökosystemforschung Wattenmeer: Zusammenfassender Bericht zu Forschungsergebnissen und Systemschutz im Deutschen Wattenmeer; Umweltbundesamt: Berlin, Germany, 2004; Volume 3. [Google Scholar]
- DWD (Deutscher Wetterdienst). Norderney. n.d. Available online: https://www.dwd.de/DE/wetter/wetterundklima_vorort/niedersachsen_bremen/norderney/_node.html (accessed on 8 April 2023).
- Strand, J.; Tairova, Z. Microplastic Particles in North Sea Sediments 2015. 2016. Available online: https://dce2.au.dk/pub/sr178.pdf (accessed on 8 April 2023).
- Hengstmann, E.; Weil, E.; Wallbott, P.C.; Tamminga, M.; Fischer, E.K. Microplastics in Lakeshore and Lakebed Sediments–External Influences and Temporal and Spatial Variabilities of Concentrations. Environ. Res. 2021, 197, 111141. [Google Scholar] [CrossRef]
- Tamminga, M.; Hengstmann, E.; Fischer, E.K. Nile Red Staining as a Subsidiary Method for Microplastic Quantifica-Tion: A Comparison of Three Solvents and Factors Influencing Application Reliability. JESES 2017, 2. [Google Scholar] [CrossRef]
- Hengstmann, E.; Tamminga, M.; vom Bruch, C.; Fischer, E.K. Microplastic in Beach Sediments of the Isle of Rügen (Baltic Sea)—Implementing a Novel Glass Elutriation Column. Mar. Pollut. Bull. 2018, 126, 263–274. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Abbatt, J.P.D.; Wang, C. The Atmospheric Chemistry of Indoor Environments. Environ. Sci. Process. Impacts 2020, 22, 25–48. [Google Scholar] [CrossRef]
- Prata, J.C.; Reis, V.; Da Costa, J.P.; Mouneyrac, C.; Duarte, A.C.; Rocha-Santos, T. Contamination Issues as a Challenge in Quality Control and Quality Assurance in Microplastics Analytics. J. Hazard. Mater. 2021, 403, 123660. [Google Scholar] [CrossRef] [PubMed]
- Hardege, J.; Bentley, M.; Snape, L. Sediment Selection by Juvenile Arenicola Marina. Mar. Ecol. Prog. Ser. 1998, 166, 187–195. [Google Scholar] [CrossRef]
- Gebhardt, C.; Forster, S. Size-Selective Feeding of Arenicola Marina Promotes Long-Term Burial of Microplastic Particles in Marine Sediments. Environ. Pollut. 2018, 242, 1777–1786. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, C. Bivalve Filter Feeding Revisited. Mar. Ecol. Prog. Ser. 1996, 142, 287–302. [Google Scholar] [CrossRef]
- Clausen, I.B.; Riisgård, H.U. Growth, Filtration and Respiration in the Mussel Mytilus Edulis: No Evidence for Physiological Regulation of the Filter-Pump to Nutritional Needs. Mar. Ecol. Prog. Ser. 1996, 141, 37–45. [Google Scholar] [CrossRef]
- Kittner, C.; Riisgård, H. Effect of Temperature on Filtration Rate in the Mussel Mytilus Edulis: No Evidence for Temperature Compensation. Mar. Ecol. Prog. Ser. 2005, 305, 147–152. [Google Scholar] [CrossRef]
- Moore, C.; Phillips, C. Plastic Ocean: How a Sea Captain’s Chance Discovery Launched a Quest to Save the Oceans; First trade paperback edition 2012; Avery a member of Penguin Group (USA): New York, NY, USA, 2012; ISBN 978-1-58333-501-7. [Google Scholar]
- Gündoğdu, S.; Çevik, C.; Ayat, B.; Aydoğan, B.; Karaca, S. How Microplastics Quantities Increase with Flood Events? An Example from Mersin Bay NE Levantine Coast of Turkey. Environ. Pollut. 2018, 239, 342–350. [Google Scholar] [CrossRef]
- Wang, J.; Lu, L.; Wang, M.; Jiang, T.; Liu, X.; Ru, S. Typhoons Increase the Abundance of Microplastics in the Marine Environment and Cultured Organisms: A Case Study in Sanggou Bay, China. Sci. Total Environ. 2019, 667, 1–8. [Google Scholar] [CrossRef]
- Hitchcock, J.N. Storm Events as Key Moments of Microplastic Contamination in Aquatic Ecosystems. Sci. Total Environ. 2020, 734, 139436. [Google Scholar] [CrossRef]
- Vandermeersch, G.; Van Cauwenberghe, L.; Janssen, C.R.; Marques, A.; Granby, K.; Fait, G.; Kotterman, M.J.J.; Diogène, J.; Bekaert, K.; Robbens, J.; et al. A Critical View on Microplastic Quantification in Aquatic Organisms. Environ. Res. 2015, 143, 46–55. [Google Scholar] [CrossRef]
- Kazour, M.; Jemaa, S.; Issa, C.; Khalaf, G.; Amara, R. Microplastics Pollution along the Lebanese Coast (Eastern Mediterranean Basin): Occurrence in Surface Water, Sediments and Biota Samples. Sci. Total Environ. 2019, 696, 133933. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Chen, G.; Wang, W.; Wang, J. Microplastic Pollution Research Methodologies, Abundance, Characteristics and Risk Assessments for Aquatic Biota in China. Environ. Pollut. 2020, 266, 115098. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Qu, X.; Su, L.; Zhang, W.; Yang, D.; Kolandhasamy, P.; Li, D.; Shi, H. Microplastics in Mussels along the Coastal Waters of China. Environ. Pollut. 2016, 214, 177–184. [Google Scholar] [CrossRef]
- Müller, C. Not as Bad as It Seems? A Literature Review on the Case of Microplastic Uptake in Fish. Front. Mar. Sci. 2021, 8, 672768. [Google Scholar] [CrossRef]
- Foekema, E.M.; De Gruijter, C.; Mergia, M.T.; van Franeker, J.A.; Murk, A.J.; Koelmans, A.A. Plastic in North Sea Fish. Environ. Sci. Technol. 2013, 47, 8818–8824. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Zhang, H.; Singh, N.; Tang, Y.; Cai, Z. Intertidal Zone Effects on Occurrence, Fate and Potential Risks of Microplastics with Perspectives under COVID-19 Pandemic. J. Chem. Eng. 2022, 429, 132351. [Google Scholar] [CrossRef]
- Setälä, O.; Norkko, J.; Lehtiniemi, M. Feeding Type Affects Microplastic Ingestion in a Coastal Invertebrate Community. Mar. Pollut. Bull. 2016, 102, 95–101. [Google Scholar] [CrossRef]
- Näkki, P.; Setälä, O.; Lehtiniemi, M. Bioturbation Transports Secondary Microplastics to Deeper Layers in Soft Marine Sediments of the Northern Baltic Sea. Mar. Pollut. Bull. 2017, 119, 255–261. [Google Scholar] [CrossRef]
- De Kluijver, M.J.; Ingalsuo, S.S.; de Bruyne, R.H. Marine Species Identification Portal: Arenicola Marina. 2021. Available online: http://species-identification.org/species.php?species_group=macrobenthos_polychaeta&id=438 (accessed on 8 April 2023).
- Krüger, F. Bau Und Leben Des Wattwurmes Arenicola Marina. Helgol. Wiss. Meeresunters. 1971, 22, 149–200. [Google Scholar] [CrossRef]
- OBIS (Oceanic Biodiversity Information System). Littorina littorea (Linnaeus, 1758). 2021. Available online: https://www.marlin.ac.uk/species/detail/1328 (accessed on 8 April 2023).
- OBIS (Oceanic Biodiversity Information System). Mytilus edulis (Linnaeus, 1758). 2021. Available online: https://www.marlin.ac.uk/species/detail/1421 (accessed on 8 April 2023).
- OBIS (Oceanic Biodiversity Information System). Platichthys flesus (Linnaeus, 1758). 2021. Available online: https://www.marlin.ac.uk/species/detail/1495 (accessed on 8 April 2023).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Geographical Exposition | Flow Exposure | Watt Type | Anthropogenic Pressure | River Basin District |
|---|---|---|---|---|---|
| Knockster Tief | west | eastern edge Ems estuary | mixed mudflats/mudflats | low | Ems (transitional waters) |
| Leybucht | north-west | bay location eastern edge Ems estuary | mixed mudflats/mudflats | low | Ems (polyhaline tidal flats) |
| Neuharlingersiel | north | main estuary Neuharlingersiel-low | mixed mudflats/mudflats | high harbor (fishing, boats, ferries, camping site, beach, residential area) | Ems (euhaline tidal flats) |
| Jadebusen | east | western edge Jade | mixed mudflats/mudflats | medium—high camping site, beach, residential area | Weser (euhaline tidal flats) |
| Tettens | east | western edge Weser estuary | mixed mudflats/mudflats | medium camping site, residential area | Weser (transitional waters) |
| Cappel- Neufeld | west | eastern edge Weser estuary | mixed mudflats/mudflats | low camping site | Weser (transitional waters) |
| Species | n (Investigated/Affected) | Affected (%) |
|---|---|---|
| Arenicola marina | 98/89 | 91 |
| Littorina littorea | 43/40 | 94 |
| Mytilus edulis | 142/121 | 85 |
| Platichthys flesus | 310/244 | 79 |
| Species | Fragment | Fiber | Microbead |
|---|---|---|---|
| Arenicola marina | 92 | 5 | 3 |
| Littorina littorea | 91 | 5 | 4 |
| Mytilus edulis | 93 | 3 | 4 |
| Platichthys flesus | 92 | 6 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polt, L.; Motyl, L.; Fischer, E.K. Abundance and Distribution of Microplastics in Invertebrate and Fish Species and Sediment Samples along the German Wadden Sea Coastline. Animals 2023, 13, 1698. https://doi.org/10.3390/ani13101698
Polt L, Motyl L, Fischer EK. Abundance and Distribution of Microplastics in Invertebrate and Fish Species and Sediment Samples along the German Wadden Sea Coastline. Animals. 2023; 13(10):1698. https://doi.org/10.3390/ani13101698
Chicago/Turabian StylePolt, Laura, Larissa Motyl, and Elke Kerstin Fischer. 2023. "Abundance and Distribution of Microplastics in Invertebrate and Fish Species and Sediment Samples along the German Wadden Sea Coastline" Animals 13, no. 10: 1698. https://doi.org/10.3390/ani13101698