
Improving Fatty Acid Profile in Native Breed Pigs Using Dietary Strategies: A Review


Abstract
:Simple Summary
Abstract
1. Introduction
2. Factors Affecting Intramuscular Fat Content (IMF) and Fatty Acid Profile
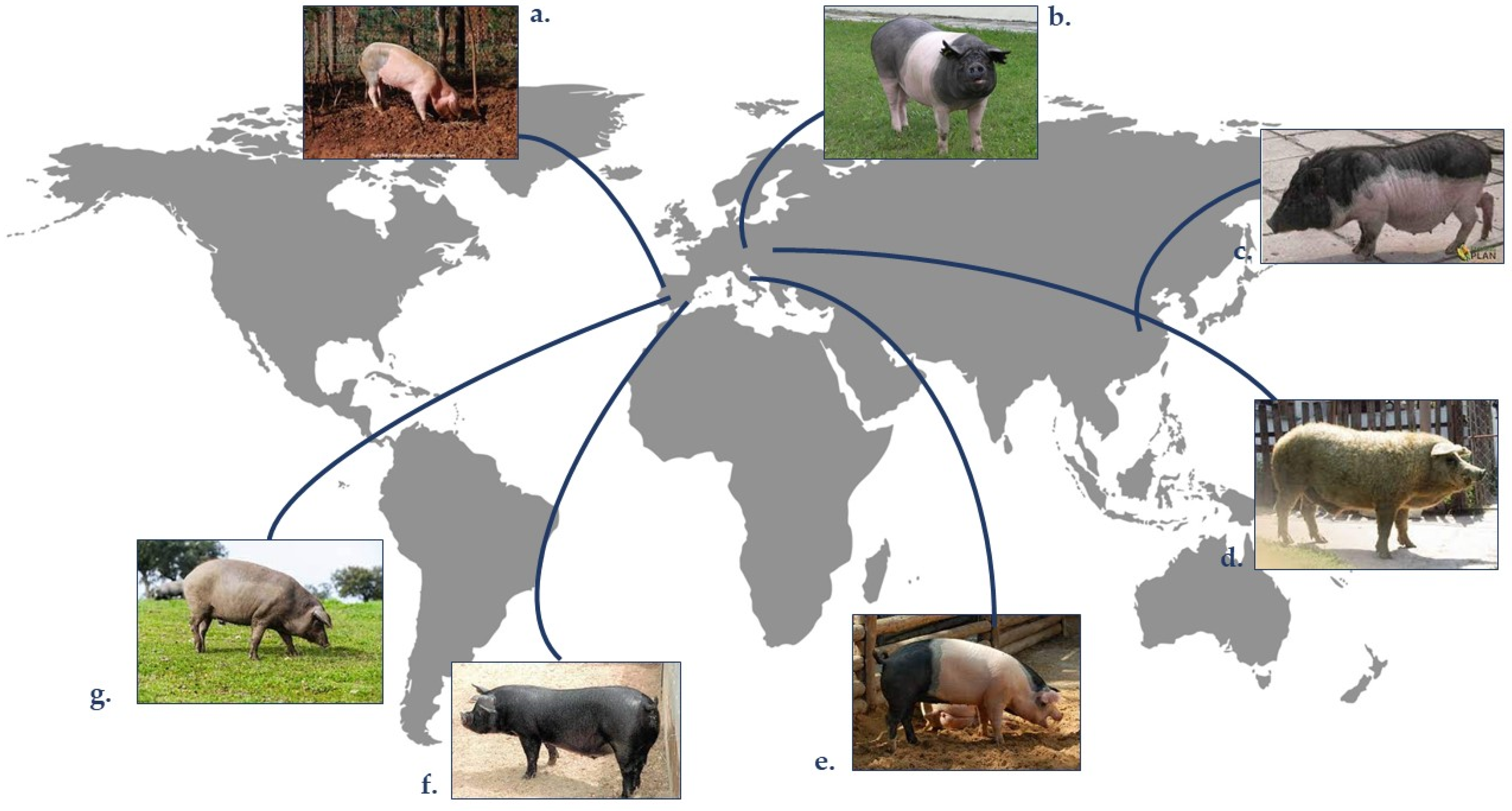
3. Indigenous Pork Fat
3.1. Intramuscular Fat
3.2. Fatty Acid Profile
4. Strategies to Improve the Fatty Acid Profile of Autochthonous Pork Meat
4.1. Acorns
4.2. Crude Olives and By-Products
4.3. Chestnuts
4.4. Sunflower Oil
4.5. Linseed
4.6. Algae
4.7. Insects
5. Conclusions: Lessons and Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leite, A.; Vasconcelos, L.; Ferreira, I.; Sarmiento-García, A.; Domínguez, R.; Santos, E.M.; Campagnol, P.C.B.; Rodrigues, S.; Lorenzo, J.M.; Teixeira, A. Chemical, Physicochemical and Sensorial Characterization of Nitrite-Free Dry-Cured Bísaro Shoulders. Foods 2022, 11, 3079. [Google Scholar] [CrossRef]
- Benítez, R.; Núñez, Y.; Fernández, A.; Isabel, B.; Rodríguez, C.; Daza, A.; López-Bote, C.; Silió, L.; Óvilo, C. Adipose Tissue Transcriptional Response of Lipid Metabolism Genes in Growing Iberian Pigs Fed Oleic Acid v. Carbohydrate Enriched Diets. Animal 2016, 10, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Vieira, C.; Sarmiento-García, A.; García, J.; Rubio, B.; Mart, B. Quality and Shelf Life of Fresh Meat from Iberian Pigs as Affected by a New Form of Presentation of Oleic Acid and an Organic-Acid Mix in the Diet. Foods 2021, 10, 985. [Google Scholar] [CrossRef] [PubMed]
- Alfaia, C.M.; Lopes, P.A.; Madeira, M.S.; Pestana, J.M.; Coelho, D.; Toldrá, F.; Prates, J.A.M. Current Feeding Strategies to Improve Pork Intramuscular Fat Content and Its Nutritional Quality. Adv. Food Nutr. Res. 2019, 89, 53–94. [Google Scholar] [CrossRef] [PubMed]
- Benítez, R.; Núñez, Y.; Ayuso, M.; Isabel, B.; Fernández-Barroso, M.A.; De Mercado, E.; Gómez-Izquierdo, E.; García-Casco, J.M.; López-Bote, C.; Óvilo, C. Changes in Biceps Femoris Transcriptome along Growth in Iberian Pigs Fed Different Energy Sources and Comparative Analysis with Duroc Breed. Animals 2021, 11, 3505. [Google Scholar] [CrossRef] [PubMed]
- Lebret, B.; Čandek-Potokar, M. Review: Pork Quality Attributes from Farm to Fork. Part I. Carcass and Fresh Meat. Animal 2022, 16, 100402. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, C.; Sirtori, F. Quality of Meat and Meat Products Produced from Southern European Pig Breeds. Meat Sci. 2012, 90, 511–518. [Google Scholar] [CrossRef]
- Szyndler-Nędza, M.; Światkiewicz, M.; Migdał, Ł.; Migdał, W. The Quality and Health-Promoting Value of Meat from Pigs of the Native Breed as the Effect of Extensive Feeding with Acorns. Animals 2021, 11, 789. [Google Scholar] [CrossRef]
- Nevrkla, P.; Kapelański, W.; Václavková, E.; Hadaš, Z.; Cebulska, A.; Horký, P. Meat Quality and Fatty Acid Profile of Pork and Backfat from an Indigenous Breed and a Commercial Hybrid of Pigs. Ann. Anim. Sci. 2017, 17, 1215–1227. [Google Scholar] [CrossRef]
- Sarmiento-García, A.; Revilla; Palacios-Riocerezo, C.; González-Martín, I. Effect of Weather Conditions on the Fatty Acid Composition of Medium-Growth Chicken Reared in Organic Production System. Braz. J. Poult. Sci. 2021, 23, 1–8. [Google Scholar] [CrossRef]
- García-Gudiño, J.; Blanco-Penedo, I.; Gispert, M.; Brun, A.; Perea, J.; Font-i-Furnols, M. Understanding Consumers’ Perceptions towards Iberian Pig Production and Animal Welfare. Meat Sci. 2021, 172, 108317. [Google Scholar] [CrossRef] [PubMed]
- Bedia, M.; Serrano, R.; Bañón, S. Use of Rustic Pigs to Manufacture Pork Salami Free of Colour and Taste Enhancers. Ital. J. Food Sci. 2012, 24, 24–2012. [Google Scholar]
- Tejeda, J.F.; Hernández-Matamoros, A.; Paniagua, M.; González, E. Effect of Free-Range and Low-Protein Concentrated Diets on Growth Performance, Carcass Traits, and Meat Composition of Iberian Pig. Animals 2020, 10, 273. [Google Scholar] [CrossRef] [PubMed]
- Nieto, R.; García-Casco, J.; Lara, L.; Palma-Granados, P.; Izquierdo, M.; Hernandez, F.; Dieguez, E.; Luis Duarte, J.; Batorek-Lukač, N. Ibérico (Iberian) Pig. In European Local Pig Breeds—Diversity and Performance. A Study of Project TREASURE; IntechOpen: London, UK, 2019. [Google Scholar]
- Franco, D.; Vazquez, J.A.; Lorenzo, J.M. Growth Performance, Carcass and Meat Quality of the Celta Pig Crossbred with Duroc and Landrance Genotypes. Meat Sci. 2014, 96, 195–202. [Google Scholar] [CrossRef]
- Gómez, M.; Lorenzo, J.M. Effect of Fat Level on Physicochemical, Volatile Compounds and Sensory Characteristics of Dry-Ripened “Chorizo” from Celta Pig Breed. Meat Sci. 2013, 95, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Auqui, S.M.; Egea, M.; Peñaranda, I.; Garrido, M.D.; Linares, M.B. Rustic Chato Murciano Pig Breed: Effect of the Weight on Carcass and Meat Quality. Meat Sci. 2019, 156, 105–110. [Google Scholar] [CrossRef]
- Peñaranda, I.; Auqui, S.M.; Egea, M.; Linares, M.B.; Garrido, M.D. Effects of Dietary Rosemary Extract Supplementation on Pork Quality of Chato Murciano Breed during Storage. Animals 2021, 11, 2295. [Google Scholar] [CrossRef]
- Leite, A.; Domínguez, R.; Vasconcelos, L.; Ferreira, I.; Pereira, E.; Pinheiro, V.; Outor-Monteiro, D.; Rodrigues, S.; Lorenzo, J.M.; Santos, E.M.; et al. Can the Introduction of Different Olive Cakes Affect the Carcass, Meat and Fat Quality of Bisaro Pork? Foods 2022, 11, 1650. [Google Scholar] [CrossRef]
- Serra, A.; Conte, G.; Giovannetti, M.; Casarosa, L.; Agnolucci, M.; Ciucci, F.; Palla, M.; Bulleri, E.; Cappucci, A.; Servili, M.; et al. Olive Pomace in Diet Limits Lipid Peroxidation of Sausages from Cinta Senese Swine. Eur. J. Lipid Sci. Technol. 2018, 120, 1700236. [Google Scholar] [CrossRef]
- Serra, A.; Buccioni, A.; Rodriguez-Estrada, M.T.; Conte, G.; Cappucci, A.; Mele, M. Fatty Acid Composition, Oxidation Status and Volatile Organic Compounds in “Colonnata” Lard from Large White or Cinta Senese Pigs as Affected by Curing Time. Meat Sci. 2014, 97, 504–512. [Google Scholar] [CrossRef]
- Gül, E.T.; Olgun, O.; Yıldız, A.; Tüzün, A.E.; Sarmiento-García, A. Use of Maca Powder (Lepidium meyenii) as Feed Additive in Diets of Laying Quails at Different Ages: Its Effect on Performance, Eggshell Quality, Serum, Ileum, and Bone Properties. Vet. Sci. 2022, 9, 418. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.D.; Richardson, R.I.; Nute, G.R.; Fisher, A.V.; Campo, M.M.; Kasapidou, E.; Sheard, P.R.; Enser, M. Effects of Fatty Acids on Meat Quality: A Review. Meat Sci. 2004, 66, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Burnett, D.D.; Legako, J.F.; Phelps, K.J.; Gonzalez, J.M. Biology, Strategies, and Fresh Meat Consequences of Manipulating the Fatty Acid Composition of Meat. J. Anim. Sci. 2020, 98, 1–12. [Google Scholar] [CrossRef]
- González, E.; Hernández-Matamoros, A.; Tejeda, J.F. Two By-Products of the Olive Oil Extraction Industry as Oleic Acid Supplement Source for Iberian Pigs: Effect on the Meat’s Chemical Composition and Induced Lipoperoxidation. J. Sci. Food Agric. 2012, 92, 2543–2551. [Google Scholar] [CrossRef] [PubMed]
- Rey, A.I.; Daza, A.; López-Carrasco, C.; López-Bote, C.J. Feeding Iberian Pigs with Acorns and Grass in Either Free-Range or Confinement Affects the Carcass Characteristics and Fatty Acids and Tocopherols Accumulation in Longissimus Dorsi Muscle and Backfat. Meat Sci. 2006, 73, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Daza, A.; Rey, A.I.; Ruiz, J.; Lopez-Bote, C.J. Effects of Feeding in Free-Range Conditions or in Confinement with Different Dietary MUFA/PUFA Ratios and α-Tocopheryl Acetate, on Antioxidants Accumulation and Oxidative Stability in Iberian Pigs. Meat Sci. 2005, 69, 151–163. [Google Scholar] [CrossRef]
- Kušec, G.; Komlenić, M.; Gvozdanović, K.; Sili, V.; Krvavica, M.; Radišić, Ž.; Kušec, I.D. Carcass Composition and Physicochemical Characteristics of Meat from Pork Chains Based on Native and Hybrid Pigs. Processes 2022, 10, 370. [Google Scholar] [CrossRef]
- Yi, W.; Huang, Q.; Wang, Y.; Shan, T. Lipo-Nutritional Quality of Pork: The Lipid Composition, Regulation, and Molecular Mechanisms of Fatty Acid Deposition. Anim. Nutr. 2023, 13, 373–385. [Google Scholar] [CrossRef]
- Morales, J.; Baucells, M.D.; Pérez, J.F.; Mourot, J.; Gasa, J. Body Fat Content, Composition and Distribution in Landrace and Iberian Finishing Pigs given Ad Libitum Maize- and Acorn-Sorghum-Maize-Based Diets. Anim. Sci. 2003, 77, 215–224. [Google Scholar] [CrossRef]
- Li, M.; Ouyang, H.; Yuan, H.; Li, J.; Xie, Z.; Wang, K.; Yu, T.; Liu, M.; Chen, X.; Tang, X.; et al. Site-Specific Fat-1 Knock-In Enables Significant Decrease of n-6PUFAs/n-3PUFAs Ratio in Pigs. G3 Genes Genomes Genet. 2018, 8, 1747–1754. [Google Scholar] [CrossRef]
- Daza, A.; Menoyo, D.; Olivares, A.; Cordero, G.; López-Bote, C.J. Effect of Iberian Pig Feeding System on Tissue Fatty-Acid Composition and Backfat Rheological Properties. J. Anim. Feed. Sci. 2007, 16, 408–419. [Google Scholar] [CrossRef]
- Turner, T.D.; Mapiye, C.; Aalhus, J.L.; Beaulieu, A.D.; Patience, J.F.; Zijlstra, R.T.; Dugan, M.E.R. Flaxseed Fed Pork: N-3 Fatty Acid Enrichment and Contribution to Dietary Recommendations. Meat Sci. 2014, 96, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Garrido, N.; Izquierdo, M.; Hernández-García, F.I.; Núñez, Y.; García-Torres, S.; Benítez, R.; Padilla, J.Á.; Óvilo, C. Differences in Muscle Lipogenic Gene Expression, Carcass Traits and Fat Deposition among Three Iberian Pig Strains Finished in Two Different Feeding Systems. Animals 2023, 13, 1138. [Google Scholar] [CrossRef] [PubMed]
- Cebulska, A.; Václavková, E.; Bocian, M.; Dybała, J.; Wiśniewska, J.; Kapelański, W. Quality and Dietary Value of Pork Meat of the Puławska and Złotnicka Spotted Breeds, and Commercial Fattening Pigs. Ann. Anim. Sci. 2018, 18, 281–291. [Google Scholar] [CrossRef]
- Tomović, V.M.; Šević, R.; Jokanović, M.; Šojić, B.; Škaljac, S.; Tasić, T.; Ikonić, P.; Polak, M.L.; Polak, T.; Demšar, L. Quality Traits of Longissimus lumborum Muscle from White Mangalica, Duroc × White Mangalica and Large White Pigs Reared under Intensive Conditions and Slaughtered at 150 kg Live Weight: A Comparative Study. Arch. Anim. Breed. 2016, 59, 401–415. [Google Scholar] [CrossRef]
- Huang, Y.; Zhou, L.; Zhang, J.; Liu, X.; Zhang, Y.; Cai, L.; Zhang, W.; Cui, L.; Yang, J.; Ji, J.; et al. A Large-Scale Comparison of Meat Quality and Intramuscular Fatty Acid Composition among Three Chinese Indigenous Pig Breeds. Meat Sci. 2020, 168, 108182. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products Nutrition and Allergies. Scientific Opinion on Dietary Reference Values for Fats, Including Saturated Fatty Acids, Polyunsaturated Fatty Acids, Monounsaturated Fatty Acids, Trans Fatty Acids, and Cholesterol. EFSA J. 2010, 8, 107. [Google Scholar] [CrossRef]
- Dugan, M.E.R.; Vahmani, P.; Turner, T.D.; Mapiye, C.; Juárez, M.; Prieto, N.; Beaulieu, A.D.; Zijlstra, R.T.; Patience, J.F.; Aalhus, J.L. Pork as a Source of Omega-3 (n-3) Fatty Acids. J. Clin. Med. 2015, 4, 1999–2011. [Google Scholar] [CrossRef]
- Teixeira, A.; Rodrigues, S. Consumer Perceptions towards Healthier Meat Products. Curr. Opin. Food Sci. 2021, 38, 147–154. [Google Scholar] [CrossRef]
- Lebret, B.; Dourmad, J.Y.; Mourot, J.; Pollet, P.Y.; Gondret, F. Production Performance, Carcass Composition, and Adipose Tissue Traits of Heavy Pigs: Influence of Breed and Production System. J. Anim. Sci. 2014, 92, 3543–3556. [Google Scholar] [CrossRef]
- Robina, A.; Viguera, J.; Perez-Palacios, T.; Mayoral, A.I.; Vivo, J.M.; Guillen, M.T.; Ruiz, J. Carcass and Meat Quality Traits of Iberian Pigs as Affected by Sex and Crossbreeding with Different Duroc Genetic Lines. Span. J. Agric. Res. 2013, 11, 1057–1067. [Google Scholar] [CrossRef]
- Heras-Molina, A.; Escudero, R.; Pesántez-Pacheco, J.L.; García-Contreras, C.; Vázquez-Gómez, M.; Astiz, S.; Óvilo, C.; González-Bulnes, A.; Isabel, B. Maternal Supplementation with Polyphenols and Omega-3 Fatty Acids during Pregnancy: Prenatal Effects on Fetal Fatty Acid Composition in the Iberian Pig. Animals 2022, 12, 2140. [Google Scholar] [CrossRef]
- Jaturasitha, S.; Chaiwang, N.; Kayan, A.; Kreuzer, M. Nutritional Strategies to Improve the Lipid Composition of Meat, with Emphasis on Thailand and Asia. Meat Sci. 2016, 120, 157–166. [Google Scholar] [CrossRef]
- Furman, M.; Malovrh, S.; Levart, A.; Kovač, M. Fatty Acid Composition of Meat and Adipose Tissue from Krškopolje Pigs and Commercial Fatteners in Slovenia. Arch. Anim. Breed. 2010, 53, 73–84. [Google Scholar] [CrossRef]
- Sarmiento-García, A.; Palacios, C.; González-Martín, I.; Revilla, I. Evaluation of the Production Performance and the Meat Quality of Chickens Reared in Organic System. As Affected by the Inclusion of Calliphora Sp. in the Diet. Animals 2021, 11, 324. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, R.; Warnakula, S.; Kunutsor, S.; Crowe, F.; Ward, H.A.; Johnson, L.; Franco, O.H.; Butterworth, A.S.; Forouhi, N.G.; Thompson, S.G.; et al. Association of Dietary, Circulating, and Supplement Fatty Acids with Coronary Risk: A Systematic Review and Meta-Analysis. Ann. Intern. Med. 2014, 160, 398–406. [Google Scholar] [CrossRef]
- López-Pedrouso, M.; Lorenzo, J.M.; Gullón, B.; Campagnol, P.C.B.; Franco, D. Novel Strategy for Developing Healthy Meat Products Replacing Saturated Fat with Oleogels. Curr. Opin. Food Sci. 2021, 40, 40–45. [Google Scholar] [CrossRef]
- Rossi, R.; Pastorelli, G.; Cannata, S.; Corino, C. Recent Advances in the Use of Fatty Acids as Supplements in Pig Diets: A Review. Anim. Feed. Sci. Technol. 2010, 162, 1–11. [Google Scholar] [CrossRef]
- De Smet, S.; Vossen, E. Meat: The Balance between Nutrition and Health. A Review. Meat Sci. 2016, 120, 145–156. [Google Scholar] [CrossRef]
- Pugliese, C.; Sirtori, F.; Acciaioli, A.; Bozzi, R.; Campodoni, G.; Franci, O. Quality of Fresh and Seasoned Fat of Cinta Senese Pigs as Affected by Fattening with Chestnut. Meat Sci. 2013, 93, 92–97. [Google Scholar] [CrossRef]
- Domínguez, R.; Martínez, S.; Gómez, M.; Carballo, J.; Franco, I. Fatty Acids, Retinol and Cholesterol Composition in Various Fatty Tissues of Celta Pig Breed: Effect of the Use of Chestnuts in the Finishing Diet. J. Food Compos. Anal. 2015, 37, 104–111. [Google Scholar] [CrossRef]
- Palma-Granados, P.; García-Casco, J.M.; Fernández-Barroso, M.A.; López-García, A.; Martínez-Torres, J.M.; Muñoz, M.; González-Sánchez, E. Inclusion of Olive By-Products in Growing Diets Causes Minor Effects on Meat Quality of Iberian Pigs Fattened in a Traditional System. Span. J. Agric. Res. 2022, 20, e0607. [Google Scholar] [CrossRef]
- Verge-Mèrida, G.; Barroeta, A.C.; Guardiola, F.; Verdú, M.; Balart, M.; Font-i-Furnols, M.; Solà-Oriol, D. Crude and Acid Oils from Olive Pomace as Alternative Fat Sources in Growing-Finishing Pigs. Animal 2021, 15, 100389. [Google Scholar] [CrossRef] [PubMed]
- Bermúdez, R.; Franco, I.; Franco, D.; Carballo, J.; Lorenzo, J.M. Influence of Inclusion of Chestnut in the Finishing Diet on Fatty Acid Profile of Dry-Cured Ham from Celta Pig Breed. Meat Sci. 2012, 92, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento-García, A.; Olgun, O.; Kilinç, G.; Sevim, B.; Gökmen, S.A. Reuse of Vegetable Wastes in Animal Feed: The Influence of Red Beet Powder Supplementation on Performance, Egg Quality, and Antioxidant Capacity of Layer Quails. Trop. Anim. Health Prod. 2023, 55, 153. [Google Scholar] [CrossRef]
- Cava, R.; Ventanas, J.; Florencio Tejeda, J.; Ruiz, J.; Antequera, T. Effect of Free-Range Rearing and α-Tocopherol and Copper Supplementation on Fatty Acid Profiles and Susceptibility to Lipid Oxidation of Fresh Meat from Iberian Pigs. Food Chem. 2000, 68, 51–59. [Google Scholar] [CrossRef]
- Liotta, L.; Chiofalo, V.; Lo Presti, V.; Chiofalo, B. In Vivo Performances, Carcass Traits, and Meat Quality of Pigs Fed Olive Cake Processing Waste. Animals 2019, 9, 1155. [Google Scholar] [CrossRef]
- Temperan, S.; Lorenzo, J.M.; Castiñeiras, B.D.; Franco, I.; Carballo, J. Carcass and Meat Quality Traits of Celta Heavy Pigs. Effect of the Inclusion of Chestnuts in the Finishing Diet. Span. J. Agric. Res. 2014, 12, 694–707. [Google Scholar] [CrossRef]
- Benítez, R.; Fernández, A.; Isabel, B.; Núñez, Y.; De Mercado, E.; Gómez-Izquierdo, E.; García-Casco, J.; López-Bote, C.; Óvilo, C. Modulatory Effects of Breed, Feeding Status, and Diet on Adipogenic, Lipogenic, and Lipolytic Gene Expression in Growing Iberian and Duroc Pigs. Int. J. Mol. Sci. 2018, 19, 22. [Google Scholar] [CrossRef]
- Mitchaothai, J.; Everts, H.; Yuangklang, C.; Wittayakun, S.; Vasupen, K.; Wongsuthavas, S.; Srenanul, P.; Hovenier, R.; Beynen, A.C. Meat Quality, Digestibility and Deposition of Fatty Acids in Growing-Finishing Pigs Fed Restricted, Iso-Energetic Amounts of Diets Containing Either Beef Tallow or Sunflower Oil. Asian-Australas. J. Anim. Sci. 2008, 21, 1015–1026. [Google Scholar] [CrossRef]
- Costa, M.; Cardoso, C.; Afonso, C.; Bandarra, N.M.; Prates, J.A.M. Current Knowledge and Future Perspectives of the Use of Seaweeds for Livestock Production and Meat Quality: A Systematic Review. J. Anim. Physiol. Anim. Nutr. 2021, 105, 1075–1102. [Google Scholar] [CrossRef] [PubMed]
- Altmann, B.A.; Neumann, C.; Rothstein, S.; Liebert, F.; Mörlein, D. Do Dietary Soy Alternatives Lead to Pork Quality Improvements or Drawbacks? A Look into Micro-Alga and Insect Protein in Swine Diets. Meat Sci. 2019, 153, 26–34. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Author | Authochonous Breed | Region | Hybrid | Fatty Acid Profile |
|---|---|---|---|---|
| Nevrkla et al. [9] | Prestice Black-Pied | Republic Czech | Large White × Landrace sows × Duroc × Pietrain boars | M. longissimus thoracis et lumborum
|
| Tomović et al. [36] | White Mangalica (M) | Hungaria | Large White (LW) and a crossbreed (LW × M) | M. longissimus lumborum
|
| Kušec et al. [28] | Black Slavonian | Croatia | (Pietrain × Duroc × Pietrain) × Camborough 23. | M. longissimus thoracis et lumborum
|
| Serra et al. [21] | Cinta Senese | Northwest of Tuscany, Italy | Large White | Lard from backfat
|
| Teixeira and Rodrigues [40] | Preto Alentejano | South of Portugal | Large White × Landrace | M. longissimus thoracis et lumborum
|
| Lebret et al. [41] | Basque | North of Spain | Large White | Subcutaneous adipose tissue
|
| Robina et al. [42] | Iberian | Spain | Duroc | Subcutaneous fat
|
| Furman et al. [45] | Krškopolje | Slovenia | Maternal hybrid Large White × Slovenian Landrace (line 11) mated by Pietrain, Duroc, or Piertain × Slovenian Landrace (line 55) sires | M. longissimus dorsi
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarmiento-García, A.; Vieira-Aller, C. Improving Fatty Acid Profile in Native Breed Pigs Using Dietary Strategies: A Review. Animals 2023, 13, 1696. https://doi.org/10.3390/ani13101696
Sarmiento-García A, Vieira-Aller C. Improving Fatty Acid Profile in Native Breed Pigs Using Dietary Strategies: A Review. Animals. 2023; 13(10):1696. https://doi.org/10.3390/ani13101696
Chicago/Turabian StyleSarmiento-García, Ainhoa, and Ceferina Vieira-Aller. 2023. "Improving Fatty Acid Profile in Native Breed Pigs Using Dietary Strategies: A Review" Animals 13, no. 10: 1696. https://doi.org/10.3390/ani13101696