
Dietary Cholesterol Differentially Regulates the Muscle Lipidomics of Farmed Turbot and Tiger Puffer


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Diets, Fish Rearing and Sampling
2.2. Muscle Lipidomic Assay
2.3. Total Lipid Content and Fatty Acid Profile
2.4. Statistics
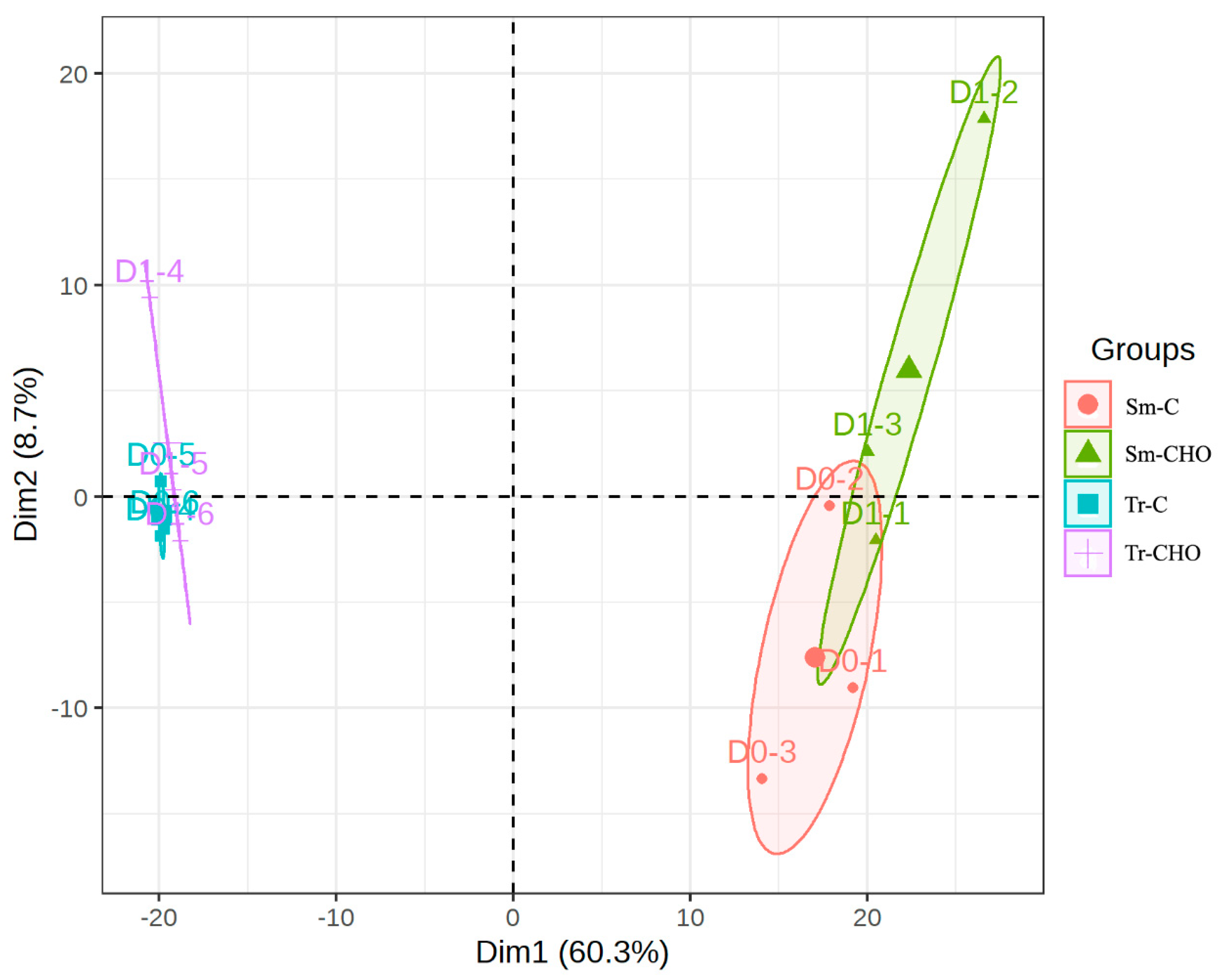
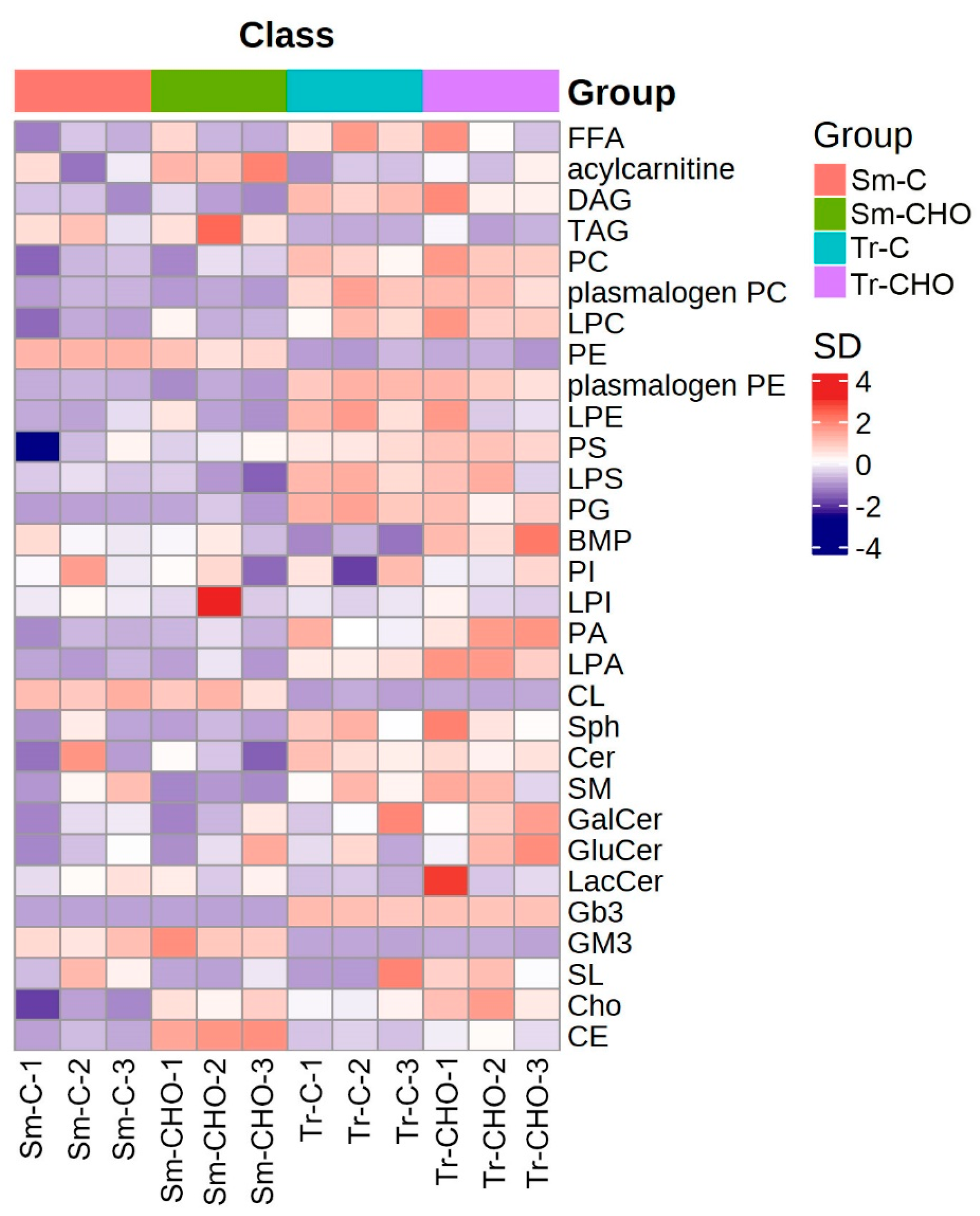
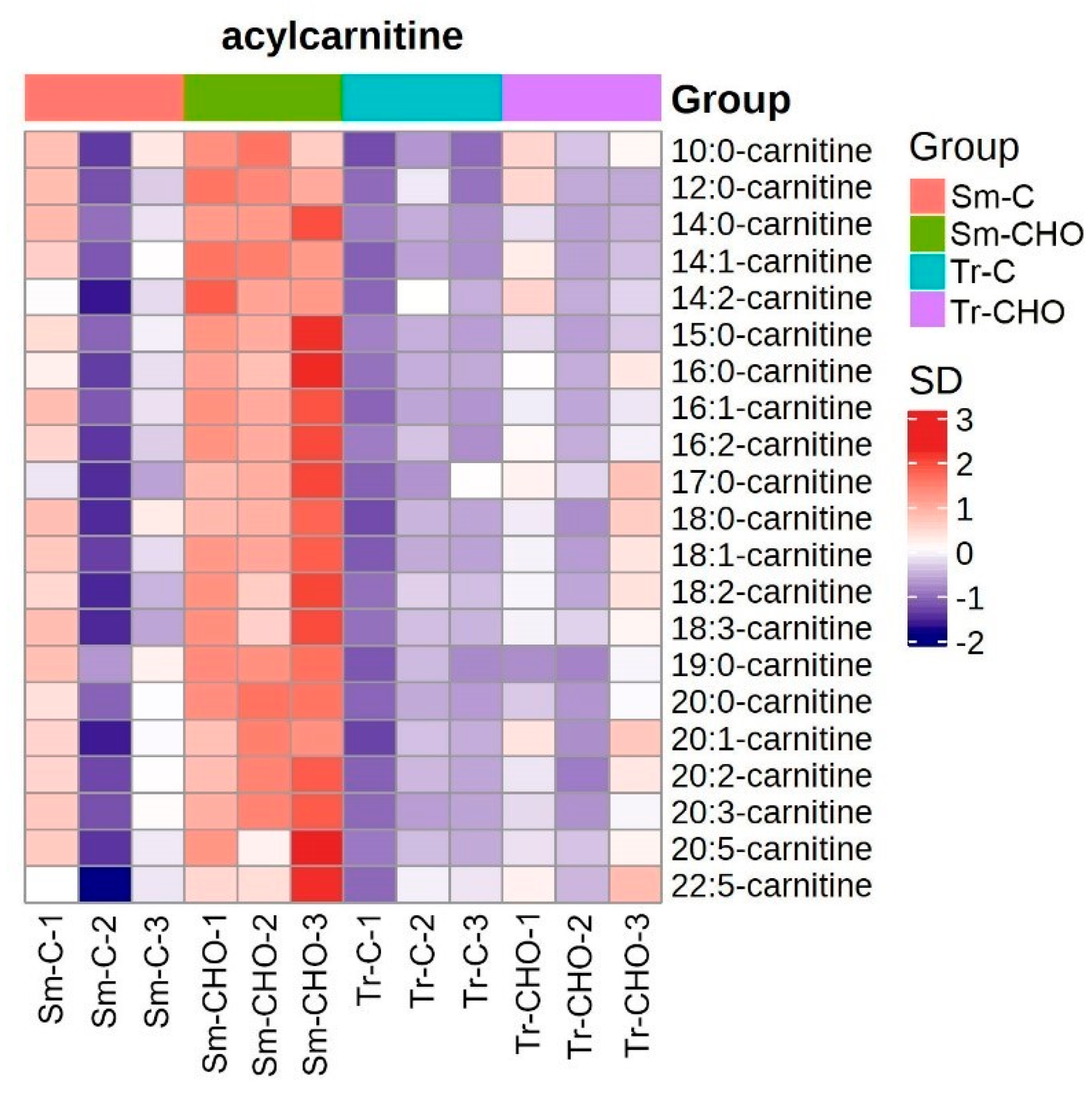
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, Z.; Hardy, R. Protein and lipid sources affect cholesterol concentrations of juvenile Pacific white shrimp, Litopenaeus vannamei (Boone). J. Anim. Sci. 2004, 82, 1136–1145. [Google Scholar] [CrossRef] [PubMed]
- Hernández, P.V.; Olvera-Novoa, M.A.; Rouse, D.B. Effect of dietary cholesterol on growth and survival of juvenile redclaw crayfish Cherax quadricarinatus under laboratory conditions. Aquaculture 2004, 236, 405–411. [Google Scholar] [CrossRef]
- Holme, M.-H.; Zeng, C.; Southgate, P.C. The effects of supplemental dietary cholesterol on growth, development and survival of mud crab, Scylla serrata, megalopa fed semi-purified diets. Aquaculture 2006, 261, 1328–1334. [Google Scholar] [CrossRef]
- Meng, X.; Bi, Q.; Cao, L.; Ma, Q.; Wei, Y.; Duan, M.; Liang, M.; Xu, H. Evaluation of necessity of cholesterol supplementation in diets of two marine teleosts, turbot (Scophthalmus maximus) and tiger puffer (Takifugu rubripes): Effects on growth and lipid metabolism. Aquac. Nutr. 2022, 2022, 4160991. [Google Scholar] [CrossRef]
- Twibell, R.G.; Wilson, R.P. Preliminary evidence that cholesterol improves growth and feed intake of soybean meal-based diets in aquaria studies with juvenile channel catfish, Ictalurus punctatus. Aquaculture 2004, 236, 539–546. [Google Scholar] [CrossRef]
- Deng, J.; Mai, K.; Ai, Q.; Zhang, W.; Wang, X.; Tan, B.; Xu, W.; Liufu, Z.; Ma, H. Interactive effects of dietary cholesterol and protein sources on growth performance and cholesterol metabolism of Japanese flounder (Paralichthys olivaceus). Aquac. Nutr. 2010, 16, 419–429. [Google Scholar] [CrossRef]
- Yun, B.; Mai, K.; Zhang, W.; Xu, W. Effects of dietary cholesterol on growth performance, feed intake and cholesterol metabolism in juvenile turbot (Scophthalmus maximus L.) fed high plant protein diets. Aquaculture 2011, 319, 105–110. [Google Scholar] [CrossRef]
- Rios-Duran, M.G.; Valencia, I.; Ross, L.; Martinez-Palacios, C.A. Nutritional evaluation of autoclaved Salicornia bigelovii Torr. seed meal supplemented with varying levels of cholesterol on growth, nutrient utilization and survival of the Nile tilapia (Oreochromis niloticus). Aquac. Int. 2013, 21, 1355–1371. [Google Scholar] [CrossRef]
- Deng, J.; Bi, B.; Kang, B.; Kong, L.; Wang, Q.; Zhang, X. Improving the growth performance and cholesterol metabolism of rainbow trout (Oncorhynchus mykiss) fed soyabean meal-based diets using dietary cholesterol supplementation. Br. J. Nutr. 2013, 110, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Zhang, X.; Long, X.; Tao, L.; Wang, Z.; Niu, G.; Kang, B. Effects of dietary cholesterol supplementation on growth and cholesterol metabolism of rainbow trout (Oncorhynchus mykiss) fed diets with cottonseed meal or rapeseed meal. Fish Physiol. Biochem. 2014, 40, 1827–1838. [Google Scholar] [CrossRef]
- Zhu, T.; Ai, Q.; Mai, K.; Xu, W.; Zhou, H.; Liufu, Z. Feed intake, growth performance and cholesterol metabolism in juvenile turbot (Scophthalmus maximus L.) fed defatted fish meal diets with graded levels of cholesterol. Aquaculture 2014, 428, 290–296. [Google Scholar] [CrossRef]
- Bjerkeng, B. Cholesterol and short-chain fatty acids in diets for Atlantic salmon Salmo salar (L.): Effects on growth, organ indices, macronutrient digestibility, and fatty acid composition. Aquac. Nutr. 1999, 5, 181–191. [Google Scholar] [CrossRef]
- Sealey, W.; Craig, S.; Gatlin, D.M. Dietary cholesterol and lecithin have limited effects on growth and body composition of hybrid striped bass (Morone chrysops× M. saxatilis). Aquac. Nutr. 2001, 7, 25–31. [Google Scholar] [CrossRef]
- Kortner, T.M.; Björkhem, I.; Krasnov, A.; Timmerhaus, G.; Krogdahl, Å. Dietary cholesterol supplementation to a plant-based diet suppresses the complete pathway of cholesterol synthesis and induces bile acid production in Atlantic salmon (Salmo salar L.). Br. J. Nutr. 2014, 111, 2089–2103. [Google Scholar] [CrossRef]
- Wu, T.M.; Jiang, J.J.; Lu, R.M.; Lin, Y.H. Effects of dietary inclusion of soybean meal and cholesterol on the growth, cholesterol status and metabolism of the giant grouper (Epinephelus lanceolatus). Aquac. Nutr. 2020, 26, 351–357. [Google Scholar] [CrossRef]
- Vega-Moreno, J.; Tirado-Cortes, A.; Álvarez, R.; Irles, C.; Mas-Oliva, J.; Ortega, A. Cholesterol depletion uncouples β-dystroglycans from discrete sarcolemmal domains, reducing the mechanical activity of skeletal muscle. Cell. Physiol. Biochem. 2012, 29, 905–918. [Google Scholar] [CrossRef]
- Hissa, B.; Oakes, P.W.; Pontes, B.; Juan, R.-S.; Gardel, M.L. Cholesterol depletion impairs contractile machinery in neonatal rat cardiomyocytes. Sci. Rep. 2017, 7, 43764. [Google Scholar] [CrossRef]
- Ignatz, E.H.; Sandrelli, R.M.; Tibbetts, S.M.; Colombo, S.M.; Zanuzzo, F.S.; Loveless, A.M.; Parrish, C.C.; Rise, M.L.; Gamperl, A.K. Influence of Supplemental Dietary Cholesterol on Growth Performance, Indices of Stress, Fillet Pigmentation, and Upper Thermal Tolerance of Female Triploid Atlantic Salmon (Salmo salar). Aquac. Nutr. 2022, 2022, 6336060. [Google Scholar] [CrossRef]
- Xu, H.; Bi, Q.; Pribytkova, E.; Wei, Y.; Sun, B.; Jia, L.; Liang, M. Different lipid scenarios in three lean marine teleosts having different lipid storage patterns. Aquaculture 2021, 536, 736448. [Google Scholar] [CrossRef]
- Xu, H.; Luo, X.; Wei, Y.; Liang, M. Dietary lysophosphatidylcholine regulates diacylglycerol, cardiolipin and free fatty acid contents in the fillet of turbot. Food Chem. X 2022, 14, 100293. [Google Scholar] [CrossRef]
- Lam, S.M.; Zhang, C.; Wang, Z.; Ni, Z.; Zhang, S.; Yang, S.; Huang, X.; Mo, L.; Li, J.; Lee, B. A multi-omics investigation of the composition and function of extracellular vesicles along the temporal trajectory of COVID-19. Nat. Metab. 2021, 3, 909–922. [Google Scholar] [CrossRef]
- Eisinger, K.; Liebisch, G.; Schmitz, G.; Aslanidis, C.; Krautbauer, S.; Buechler, C. Lipidomic analysis of serum from high fat diet induced obese mice. Int. J. Mol. Sci. 2014, 15, 2991–3002. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-F.; Wu, W.-Y.; Qiu, G.-K.; Wang, H.; Li, W.-S.; Wang, Y.-L.; Jiang, Q.-Q.; Han, M.-F.; Ning, Q. Plasma lipidomics identifies novel biomarkers in patients with hepatitis B virus-related acute-on-chronic liver failure. Metabolomics 2017, 13, 76. [Google Scholar] [CrossRef]
- Pousinis, P.; Gowler, P.R.; Burston, J.J.; Ortori, C.A.; Chapman, V.; Barrett, D. Lipidomic identification of plasma lipids associated with pain behaviour and pathology in a mouse model of osteoarthritis. Metabolomics 2020, 16, 32. [Google Scholar] [CrossRef]
- Uhl, O.; Fleddermann, M.; Hellmuth, C.; Demmelmair, H.; Koletzko, B. Phospholipid species in newborn and 4 month old infants after consumption of different formulas or breast milk. PLoS ONE 2016, 11, e0162040. [Google Scholar] [CrossRef]
- Kay, J.G.; Grinstein, S. Sensing phosphatidylserine in cellular membranes. Sensors 2011, 11, 1744–1755. [Google Scholar] [CrossRef] [PubMed]
- Kooijman, E.E.; Burger, K.N. Biophysics and function of phosphatidic acid: A molecular perspective. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2009, 1791, 881–888. [Google Scholar] [CrossRef]
- Cummings, R.; Parinandi, N.; Wang, L.; Usatyuk, P.; Natarajan, V. Phospholipase D/phosphatidic acid signal transduction: Role and physiological significance in lung. Mol. Cell. Biochem. 2002, 234, 99–109. [Google Scholar] [CrossRef]
- Wang, T.; Wang, X.; Zhou, H.; Jiang, H.; Mai, K.; He, G. The mitotic and metabolic effects of phosphatidic acid in the primary muscle cells of turbot (Scophthalmus maximus). Front. Endocrinol. 2018, 9, 221. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Sun, B.; Zhang, Q.; Jia, L.; Wei, Y.; Liang, M.; Xu, H. Dietary bile acids regulate the hepatic lipid homeostasis in tiger puffer fed normal or high-lipid diets. Aquaculture 2020, 519, 734935. [Google Scholar] [CrossRef]
- Indiveri, C.; Iacobazzi, V.; Tonazzi, A.; Giangregorio, N.; Infantino, V.; Convertini, P.; Console, L.; Palmieri, F. The mitochondrial carnitine/acylcarnitine carrier: Function, structure and physiopathology. Mol. Asp. Med. 2011, 32, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Jarrell, Z.R.; Smith, M.R.; Hu, X.; Orr, M.; Liu, K.H.; Quyyumi, A.A.; Jones, D.P.; Go, Y.-M. Plasma acylcarnitine levels increase with healthy aging. Aging (Albany NY) 2020, 12, 13555. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, K.; Takami, T.; Shintani, H.; Sasai, N.; Matsumoto, T.; Yamamoto, N.; Sakaida, I. Seasonal variations in photoperiod affect hepatic metabolism of medaka (Oryzias latipes). FEBS Open Bio 2021, 11, 1029–1040. [Google Scholar] [CrossRef]
- Akgoc, Z.; Sena-Esteves, M.; Martin, D.R.; Han, X.; d’Azzo, A.; Seyfried, T.N. Bis (monoacylglycero) phosphate: A secondary storage lipid in the gangliosidoses. J. Lipid Res. 2015, 56, 1005–1006. [Google Scholar] [CrossRef] [PubMed]
- Gallala, H.D.; Sandhoff, K. Biological function of the cellular lipid BMP—BMP as a key activator for cholesterol sorting and membrane digestion. Neurochem. Res. 2011, 36, 1594–1600. [Google Scholar] [CrossRef]
- Hullin-Matsuda, F.; Luquain-Costaz, C.; Bouvier, J.; Delton-Vandenbroucke, I. Bis (monoacylglycero) phosphate, a peculiar phospholipid to control the fate of cholesterol: Implications in pathology. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 313–324. [Google Scholar] [CrossRef]
- Chevallier, J.; Chamoun, Z.; Jiang, G.; Prestwich, G.; Sakai, N.; Matile, S.; Parton, R.G.; Gruenberg, J. Lysobisphosphatidic acid controls endosomal cholesterol levels. J. Biol. Chem. 2008, 283, 27871–27880. [Google Scholar] [CrossRef]
- van Meer, G.; Wolthoorn, J.; Degroote, S. The fate and function of glycosphingolipid glucosylceramide. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2003, 358, 869–873. [Google Scholar] [CrossRef]
- Hashimoto, N.; Matsumoto, I.; Takahashi, H.; Ashikawa, H.; Nakamura, H.; Murayama, T. Cholesterol-dependent increases in glucosylceramide synthase activity in Niemann-Pick disease type C model cells: Abnormal trafficking of endogenously formed ceramide metabolites by inhibition of the enzyme. Neuropharmacology 2016, 110, 458–469. [Google Scholar] [CrossRef]
- Varela, A.R.; Couto, A.S.; Fedorov, A.; Futerman, A.H.; Prieto, M.; Silva, L.C. Glucosylceramide reorganizes cholesterol-containing domains in a fluid phospholipid membrane. Biophys. J. 2016, 110, 612–622. [Google Scholar] [CrossRef]
- Cheng, K.; Bou, M.; Ruyter, B.; Pickova, J.; Ehtesham, E.; Du, L.; Venegas, C.; Moazzami, A. Reduced dietary levels of EPA and DHA have a major impact on the composition of skin membrane lipids in Atlantic salmon (Salmo salar L.). J. Agric Food Chem. 2018, 66, 8876–8884. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | Diet | Turbot Muscle | Tiger Puffer Muscle | |||
|---|---|---|---|---|---|---|
| Control | CHO | Control | CHO | Control | CHO | |
| 14:0 | 2.81 | 2.71 | 1.32 ± 0.09 | 1.19 ± 0.05 | 0.42 ± 0.01 | 0.45 ± 0.02 |
| 16:0 | 17.4 | 17.0 | 21.0 ± 0.14 | 21.7 ± 0.09 * | 19.7 ± 0.14 | 20.3 ± 0.13 * |
| 18:0 | 5.09 | 5.02 | 8.61 ± 0.02 | 8.13 ± 0.07 | 12.5 ± 0.08 | 12.1 ± 0.18 |
| ∑SFA | 26.4 | 25.8 | 31.6 ± 0.13 | 31.7 ± 0.10 | 33.2 ± 0.18 | 33.5 ± 0.09 |
| 16:1n-7 | 3.23 | 3.26 | 1.49 ± 0.08 | 1.33 ± 0.02 | 0.65 ± 0.01 | 0.71 ± 0.03 |
| 18:1n-9 | 24.7 | 24.2 | 18.3 ± 0.40 | 17.5 ± 0.12 * | 13.3 ± 0.13 | 13.9 ± 0.10 * |
| 20:1n-9 | 0.62 | 0.60 | 0.56 ± 0.02 | 0.60 ± 0.03 | 0.78 ± 0.02 | 0.79 ± 0.01 |
| ∑MUFA | 28.8 | 28.2 | 20.9 ± 0.51 | 20.0 ± 0.10 * | 15.2 ± 0.16 | 16.0 ± 0.08 * |
| 18:2n-6 | 23.3 | 23.4 | 19.3 ± 0.21 | 19.5 ± 0.21 | 21.3 ± 0.15 | 21.5 ± 0.22 |
| 20:2n-6 | 0.00 | 0.00 | 0.73 ± 0.05 | 0.79 ± 0.03 | 1.43 ± 0.03 | 1.31 ± 0.08 |
| 20:4n-6 | 0.45 | 0.45 | 1.15 ± 0.05 | 1.25 ± 0.01 * | 1.34 ± 0.03 | 1.28 ± 0.04 |
| ∑n-6PUFA | 24.0 | 24.1 | 21.2 ± 0.20 | 21.5 ± 0.22 | 24.1 ± 0.15 | 24.1 ± 0.25 |
| 18:3n-3 | 13.7 | 14.3 | 6.70 ± 0.14 | 6.51 ± 0.08 | 3.59 ± 0.10 | 3.72 ± 0.15 |
| 20:5n-3 | 4.32 | 4.67 | 6.32 ± 0.10 | 6.47 ± 0.10 | 5.21 ± 0.07 | 5.33 ± 0.06 |
| 22:5n-3 | 0.60 | 0.63 | 2.20 ± 0.01 | 2.27 ± 0.05 | 5.76 ± 0.09 | 5.01 ± 0.15 * |
| 22:6n-3 | 2.13 | 2.28 | 10.5 ± 0.41 | 11.0 ± 0.23 | 12.1 ± 0.25 | 11.7 ± 0.42 |
| ∑n-3PUFA | 20.8 | 21.9 | 26.2 ± 0.31 | 26.8 ± 0.39 | 27.5 ± 0.32 | 26.5 ± 0.29 * |
| n-3/n-6 | 0.87 | 0.91 | 1.24 ± 0.01 | 1.25 ± 0.03 | 1.14 ± 0.02 | 1.10 ± 0.02 |
| Lipid | p Value | Fold | Lipid | p Value | Fold |
|---|---|---|---|---|---|
| Turbot | Tiger puffer | ||||
| Class | Class | ||||
| CE | 0.0001 | 6.23↑ | BMP | 0.0203 | 2.01↑ |
| Cholesterol | 0.0127 | 1.26↑ | PS | 0.0273 | 1.03↑ |
| Specific lipid | Specific lipid | ||||
| PE36:1 | 0.0011 | 0.81↓ | PA38:3(18:0_20:3) | 0.0006 | 1.26↑ |
| PG38:4(18:0_20:4) | 0.0058 | 1.31↑ | CE-20:1 | 0.0084 | 3.06↑ |
| TAG52:4(16:1) | 0.0061 | 1.48↑ | PC40:6(22:5_18:1) | 0.0086 | 0.91↓ |
| SM d18:1/23:0 | 0.0081 | 0.70↓ | PS38:6 | 0.0087 | 1.13↑ |
| PG36:4(16:0_20:4) | 0.0090 | 1.36↑ | PC36:2p(18:0p_18:2) | 0.0099 | 1.13↑ |
| TAG52:6(16:1) | 0.0094 | 1.55↑ | PA40:6(18:0_22:6) | 0.0112 | 1.59↑ |
| SM d18:1/22:1 | 0.0104 | 0.78↓ | LPA16:0 | 0.0133 | 1.31↑ |
| PE38:5 | 0.0123 | 0.88↓ | PA 36:1(18:0_18:1) | 0.0152 | 1.94↑ |
| TAG52:6(18:3) | 0.0125 | 1.52↑ | LPI20:4 | 0.0175 | 0.78↓ |
| PC42:5 | 0.0129 | 1.32↑ | PC34:1(16:1_18:0) | 0.0189 | 1.14↑ |
| TAG52:5(16:0) | 0.0133 | 1.51↑ | PC36:1p(18:0p_18:1) | 0.0195 | 1.07↑ |
| CL72:5(18:2) | 0.0148 | 0.90↓ | BMP36:3(18:2_18:1) | 0.0203 | 2.01↑ |
| TAG56:7(22:5) | 0.0166 | 1.65↑ | LPA16:1 | 0.0205 | 1.26↑ |
| 17:0-carnitine | 0.0194 | 2.31↑ | PS38:3 | 0.0213 | 1.22↑ |
| PE38:5p | 0.0217 | 0.81↓ | PE40:5p(18:0p_22:5) | 0.0224 | 0.81↓ |
| CE-18:2 | 0.0221 | 7.69↑ | GluCer d18:1/15:0 | 0.0284 | 2.44↑ |
| TAG52:5(16:1) | 0.0235 | 1.49↑ | PC34:3 | 0.0288 | 1.12↑ |
| SM d18:1/22:0 | 0.0237 | 0.70↓ | 10:0-carnitine | 0.0312 | 1.64↑ |
| TAG52:4(16:0) | 0.0239 | 1.38↑ | PA38:4(18:0_20:4) | 0.0316 | 1.51↑ |
| PE40:6(20:2_20:4) | 0.0249 | 1.21↑ | PA40:5(18:0_22:5) | 0.0369 | 1.39↑ |
| PC34:2p | 0.0263 | 0.83↓ | PS38:4 | 0.0375 | 1.22↑ |
| TAG48:4(16:1) | 0.0297 | 1.82↑ | PC32:1 | 0.0377 | 1.09↑ |
| CE-22:3 | 0.0299 | 7.59↑ | LPI22:5 | 0.0387 | 0.67↓ |
| PE36:3p(16:0p_20:3) | 0.0300 | 0.81↓ | CL74:7(18:2) | 0.0392 | 1.10↑ |
| PI38:5 | 0.0302 | 0.85↓ | PA38:3 | 0.0414 | 1.43↑ |
| CE-22:5 | 0.0333 | 3.85↑ | TAG58:9(22:5) | 0.0432 | 0.77↓ |
| PC38:2p | 0.0337 | 0.86↓ | PE40:4p | 0.0470 | 0.85↓ |
| CE-18:3 | 0.0340 | 10.9↑ | PS38:3(18:0_20:3) | 0.0479 | 0.90↓ |
| PG36:4(16:1_20:3) | 0.0346 | 1.24↑ | PC34:2p(16:0p_18:2) | 0.0484 | 1.15↑ |
| TAG52:4(16:2) | 0.0350 | 1.33↑ | PC42:7(22:5_20:2) | 0.0498 | 0.81↓ |
| CE-20:3 | 0.0357 | 14.5↑ | |||
| PS32:1 | 0.0364 | 1.10↑ | |||
| TAG52:4(18:1) | 0.0376 | 1.42↑ | |||
| CE-22:6 | 0.0377 | 3.44↑ | |||
| TAG52:5(18:3) | 0.0382 | 1.48↑ | |||
| PE34:1 | 0.0396 | 0.82↓ | |||
| SM d18:1/14:1 | 0.0420 | 1.24↑ | |||
| CE-18:1 | 0.0428 | 14.9↑ | |||
| PC40:3p | 0.0428 | 0.89↓ | |||
| CE-20:5 | 0.0432 | 5.23↑ | |||
| 15:0-carnitine | 0.0433 | 2.34↑ | |||
| PC32:1 | 0.0443 | 1.04↑ | |||
| CE-20:2 | 0.0467 | 14.4↑ | |||
| SM d18:1/23:1 | 0.0467 | 0.80↓ | |||
| 14:2-carnitine | 0.0486 | 1.97↑ | |||
| CE-20:4 | 0.0487 | 5.45↑ | |||
| TAG50:3(18:2) | 0.0489 | 1.38↑ | |||
| TAG48:3(16:0) | 0.0491 | 1.66↑ | |||
| 16:0-carnitine | 0.0497 | 2.45↑ | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, X.; Bi, Q.; Ma, Q.; Wei, Y.; Li, Y.; Liang, M.; Xu, H. Dietary Cholesterol Differentially Regulates the Muscle Lipidomics of Farmed Turbot and Tiger Puffer. Animals 2023, 13, 1632. https://doi.org/10.3390/ani13101632
Meng X, Bi Q, Ma Q, Wei Y, Li Y, Liang M, Xu H. Dietary Cholesterol Differentially Regulates the Muscle Lipidomics of Farmed Turbot and Tiger Puffer. Animals. 2023; 13(10):1632. https://doi.org/10.3390/ani13101632
Chicago/Turabian StyleMeng, Xiaoxue, Qingzhu Bi, Qiang Ma, Yuliang Wei, Yanlu Li, Mengqing Liang, and Houguo Xu. 2023. "Dietary Cholesterol Differentially Regulates the Muscle Lipidomics of Farmed Turbot and Tiger Puffer" Animals 13, no. 10: 1632. https://doi.org/10.3390/ani13101632