
Investigations of Fish Assemblages Using Two Methods in Three Terminal Reservoirs of the East Route of South-to-North Water Transfer Project, China
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
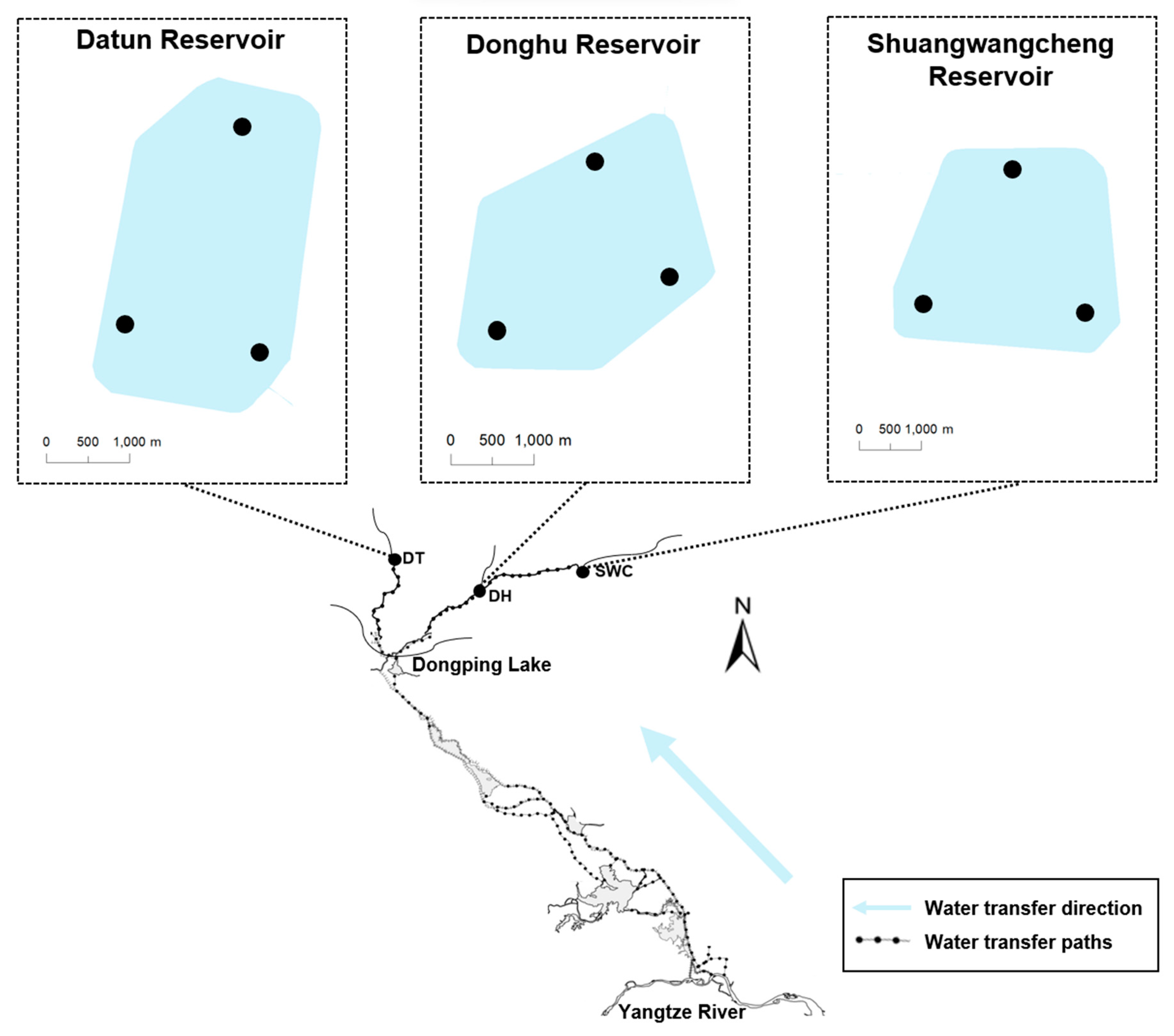
2.1. Study Area
2.2. Samplings Processing of TFL
2.3. eDNA Sample Processing
2.4. Laboratory Processing of eDNA Samples
2.5. Data Analysis
3. Results
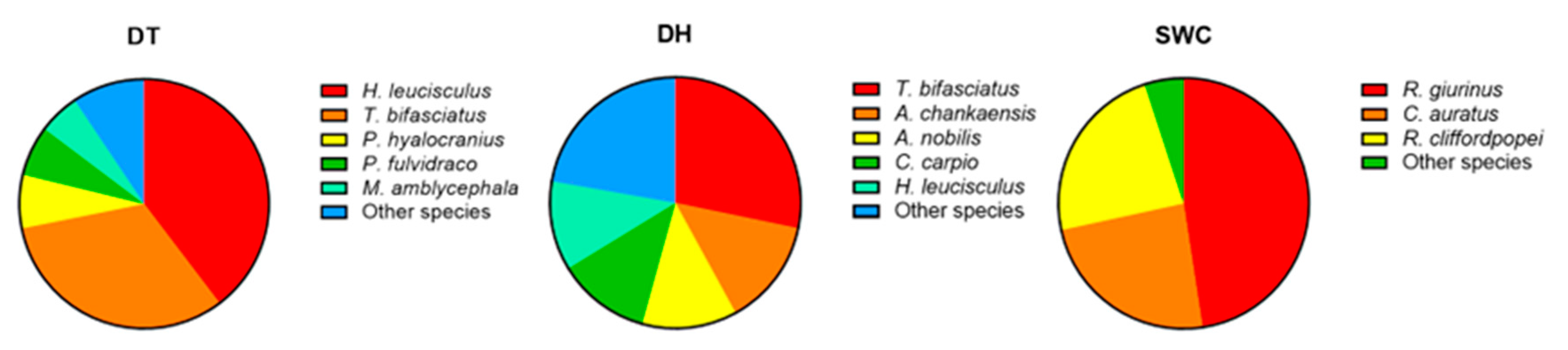
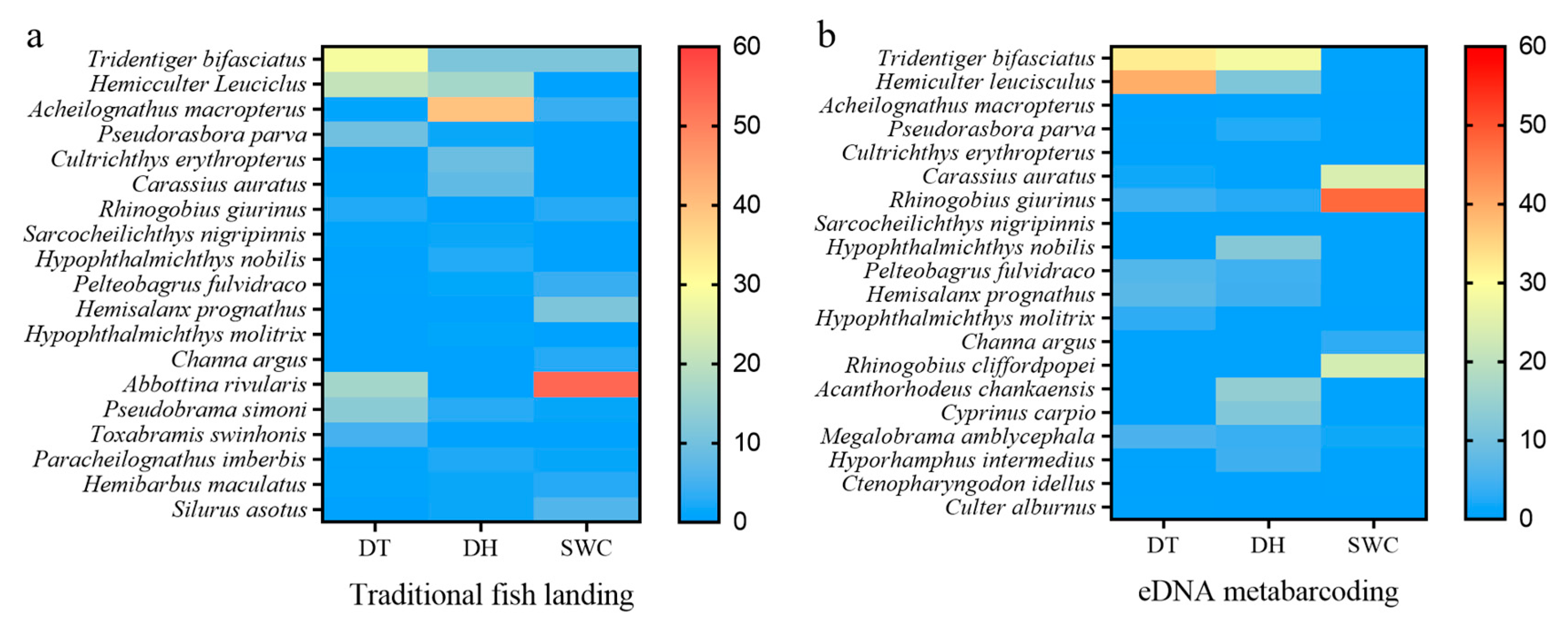
3.1. Fish Assemblages Identified through Traditional Fish Landing (TFL) Analysis
3.2. Fish Assemblages Detected through eDNA Metabarcoding
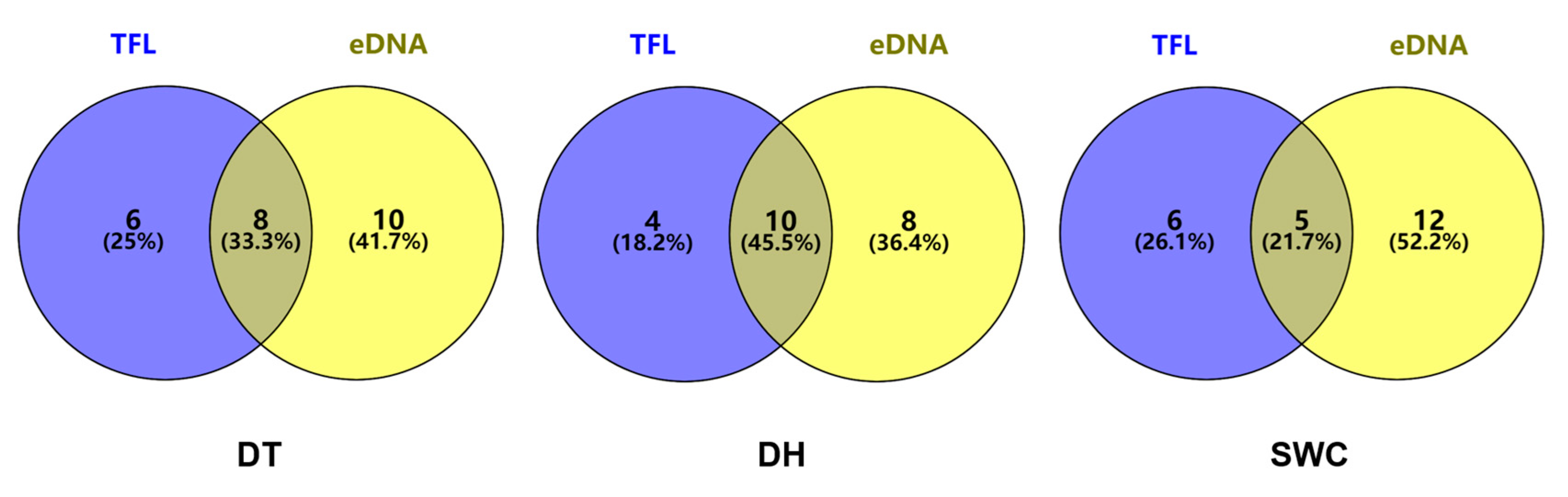
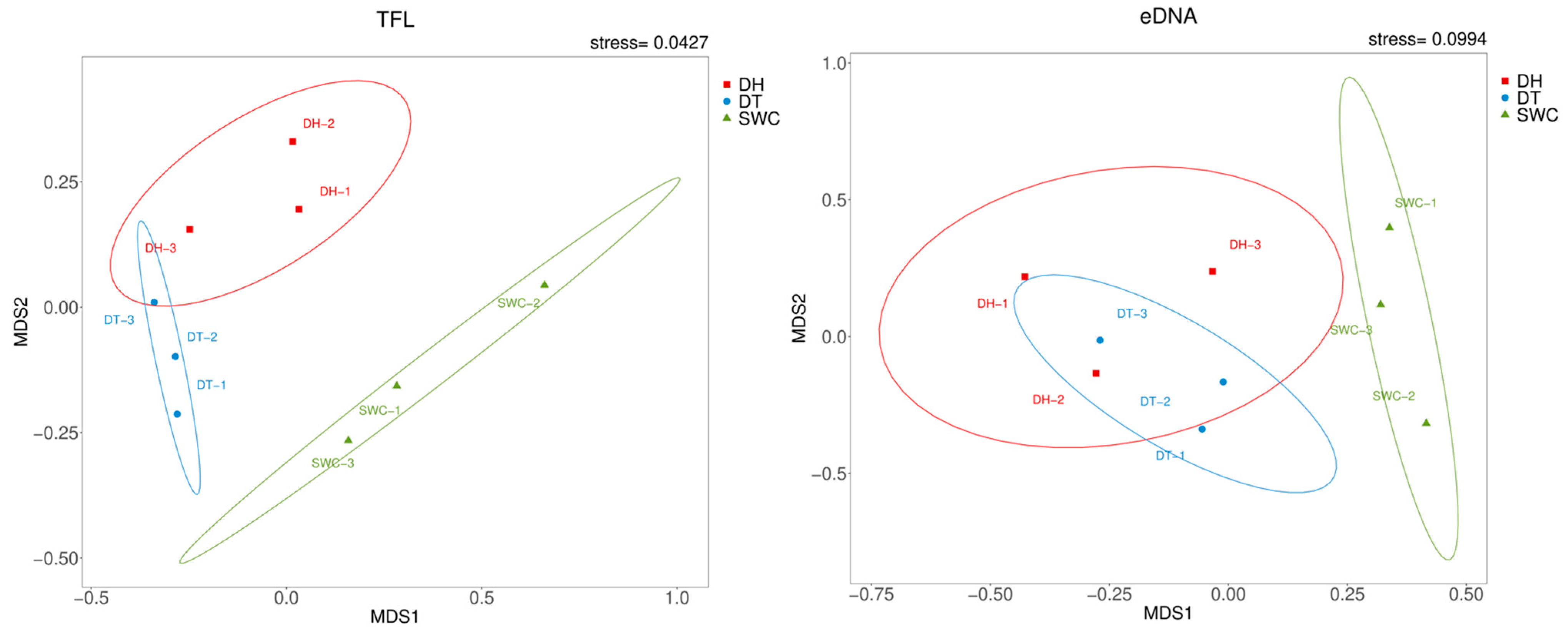
3.3. Comparison of Traditional Fish Landing (TFL) and eDNA Metabarcoding Methods
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, C.; Chen, Y.; Gozlan, R.E.; Liu, H.; Lu, Y.; Qu, X.; Xia, W.; Xiong, F.; Xie, S.; Wang, L. Patterns of fish communities and water quality in impounded lakes of China’s South-To-North Water Diversion Project. Sci. Total Environ. 2020, 713, 136515. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, J.; Zhang, Y.; Shi, J.; Yu, H.; Zhang, X. eDNA biomonitoring revealed the ecological effects of water diversion projects between Yangtze River and Tai Lake. Water Res. 2022, 210, 117994. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, A.; Mirchi, A.; Pirmoradian, R.; Mirabbasi, R.; Tourian, M.J.; Haghighi, A.T.; Madani, K. Battling water limits to growth: Lessons from water trends in the central plateau of Iran. Environ. Manag. 2021, 68, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, B.; Wang, Z.; Ren, P.; Guo, C.; Qin, J.; Cheng, F.; Xie, S. A review of potential factors promoting fish movement in inter-basin water transfers, with emergent patterns from a trait-based risk analysis for a large-scale project in China. Ecol. Freshw. Fish 2020, 29, 790–807. [Google Scholar] [CrossRef]
- Liu, D.; Wang, R.; Gordon, D.R.; Sun, X.; Chen, L.; Wang, Y. Predicting Plant invasions following China’s water diversion project. Environ. Sci. Technol. 2017, 51, 1450–1457. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Jia, C.; Liu, Q.; Li, Z.; Liu, P.; Yang, M.; Zhao, M.; Li, W.; Zhu, H.; Zhang, Q. Dynamics of bacterial community diversity and structure in the terminal reservoir of the South-To-North Water Diversion Project in China. Water 2018, 10, 709. [Google Scholar] [CrossRef]
- Feng, S.; Li, L.X.; Duan, Z.G.; Zhang, J.L. Assessing the impacts of South-To-North Water Transfer Project with decision support systems. Decis. Support Syst. 2007, 42, 1989–2003. [Google Scholar] [CrossRef]
- Xiao, R.; Shi, H.; He, X.; Li, Z.; Jia, D.; Yang, Z. Deformation monitoring of reservoir dams using GNSS: An application to South-to-North Water Diversion Project, China. IEEE Access 2019, 7, 54981–54992. [Google Scholar] [CrossRef]
- Zang, N.; Zhu, J.; Wang, X.; Liao, Y.; Cao, G.; Li, C.; Liu, Q.; Yang, Z. Eutrophication risk assessment considering joint effects of water quality and water quantity for a receiving reservoir in the South-to-North Water Transfer Project, China. J. Clean. Prod. 2022, 331, 129966. [Google Scholar] [CrossRef]
- Ho, L.T.; Goethals, P.L.M. Opportunities and challenges for the sustainability of lakes and reservoirs in relation to the sustainable development goals (SDGs). Water 2021, 13, 3207. [Google Scholar] [CrossRef]
- Deacon, J.R.; Mize, S.V.; Survey, G. Effects of Water Quality and Habitat on Composition of Fish Communities in the Upper Colorado River Basin; US Department of the Interior, US Geological Survey: Asheville, NC, USA, 1997.
- Moyle, P. Biodiversity, Biomonitoring, and the Structure of Stream Fish Communities; LEWIS Publishers Inc.: Boca Raton, FL, USA, 1994; pp. 171–186. [Google Scholar]
- Xie, P.; Liu, J. Practical success of biomanipulation using filter-feeding fish to control cyanobacteria blooms: A synthesis of decades of research and application in a subtropical hypereutrophic lake. Sci. World J. 2001, 1, 337–356. [Google Scholar] [CrossRef] [PubMed]
- Peng, G.; Zhou, X.; Xie, B.; Huang, C.; Uddin, M.M.; Chen, X.; Huang, L. Ecosystem stability and water quality improvement in a eutrophic shallow lake via long-term integrated biomanipulation in Southeast China. Ecol. Eng. 2021, 159, 106119. [Google Scholar] [CrossRef]
- Blabolil, P.; Logez, M.; Ricard, D.; Prchalova, M.; Riha, M.; Sagouis, A.; Peterka, J.; Kubecka, J.; Argillier, C. An assessment of the ecological potential of Central and Western European reservoirs based on fish communities. Fish. Res. 2016, 173, 80–87. [Google Scholar] [CrossRef]
- Al-Baz, A.; Bishop, J.M.; Al-Husaini, M.; Chen, W. Gargoor trap fishery in Kuwait, catch rate and species composition. J. Appl. Ichthyol. 2018, 34, 867–877. [Google Scholar] [CrossRef]
- Meador, M.R.; McIntyre, J.P.; Pollock, K.H. Assessing the efficacy of single-pass backpack electrofishing to characterize fish community structure. Trans. Am. Fish. Soc. 2003, 132, 39–46. [Google Scholar] [CrossRef]
- Bayley, P.B.; Peterson, J.T. An approach to estimate probability of presence and richness of fish species. Trans. Am. Fish. Soc. 2001, 130, 620–633. [Google Scholar] [CrossRef]
- Gu, W.; Swihart, R.K. Absent or undetected? Effects of non-detection of species occurrence on wildlife–habitat models. Biol. Conserv. 2004, 116, 195–203. [Google Scholar] [CrossRef]
- Kubecka, J.; Hohausova, E.; Matena, J.; Peterka, J.; Amarasinghe, U.S.; Bonar, S.A.; Hateley, J.; Hickley, P.; Suuronen, P.; Tereschenko, V.; et al. The true picture of a lake or reservoir fish stock: A review of needs and progress. Fish. Res. 2009, 96, 1–5. [Google Scholar] [CrossRef]
- Olds, B.P.; Jerde, C.L.; Renshaw, M.A.; Li, Y.; Evans, N.T.; Turner, C.R.; Deiner, K.; Mahon, A.R.; Brueseke, M.A.; Shirey, P.D.; et al. Estimating species richness using environmental DNA. Ecol. Evol. 2016, 6, 4214–4226. [Google Scholar] [CrossRef] [PubMed]
- Valentini, A.; Taberlet, P.; Miaud, C.; Civade, R.; Herder, J.; Thomsen, P.F.; Bellemain, E.; Besnard, A.; Coissac, E.; Boyer, F.; et al. Next-generation monitoring of aquatic biodiversity using environmental DNA metabarcoding. Mol. Ecol. 2016, 25, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Andruszkiewicz, E.A.; Starks, H.A.; Chavez, F.P.; Sassoubre, L.M.; Block, B.A.; Boehm, A.B. Biomonitoring of marine vertebrates in Monterey Bay using eDNA metabarcoding. PLoS ONE 2017, 12, e0176343. [Google Scholar] [CrossRef]
- Cilleros, K.; Valentini, A.; Allard, L.; Dejean, T.; Etienne, R.; Grenouillet, G.; Iribar, A.; Taberlet, P.; Vigouroux, R.; Brosse, S. Unlocking biodiversity and conservation studies in high-diversity environments using environmental DNA (eDNA): A test with Guianese freshwater fishes. Mol. Ecol. Resour. 2019, 19, 27–46. [Google Scholar] [CrossRef]
- Pukk, L.; Kanefsky, J.; Heathman, A.L.; Weise, E.M.; Nathan, L.R.; Herbst, S.J.; Sard, N.M.; Scribner, K.T.; Robinson, J.D. eDNA metabarcoding in lakes to quantify influences of landscape features and human activity on aquatic invasive species prevalence and fish community diversity. Divers. Distrib. 2021, 27, 2016–2031. [Google Scholar] [CrossRef]
- Li, H.; Yang, F.; Zhang, R.; Liu, S.; Yang, Z.; Lin, L.; Ye, S. Environmental DNA metabarcoding of fish communities in a small hydropower dam reservoir: A comparison between the eDNA approach and established fishing methods. J. Freshw. Ecol. 2022, 37, 337–358. [Google Scholar] [CrossRef]
- Fujii, K.; Doi, H.; Matsuoka, S.; Nagano, M.; Sato, H.; Yamanaka, H. Environmental DNA metabarcoding for fish community analysis in backwater lakes: A comparison of capture methods. PLoS ONE 2019, 14, e0210357. [Google Scholar] [CrossRef]
- Guo, C.; Chen, Y.; Liu, H.; Lu, Y.; Qu, X.; Yuan, H.; Lek, S.; Xie, S. Modelling fish communities in relation to water quality in the impounded lakes of China’s South-to-North Water Diversion Project. Ecol. Model. 2019, 397, 25–35. [Google Scholar] [CrossRef]
- Eros, T.; Specziar, A.; Biro, P. Assessing fish assemblages in reed habitats of a large shallow lake-a comparison between gillnetting and electric fishing. Fish. Res. 2009, 96, 70–76. [Google Scholar] [CrossRef]
- Chen, Y. Fauna Sinica: Osteichthyes: Cypriniformes II; Science Press: Beijing, China, 1998. [Google Scholar]
- Cheng, Q.; Zheng, B. Systematic Synopsis of Chinese Fishes; Science Press: Beijing, China, 1987. [Google Scholar]
- Chu, X.L.; Zheng, B.S.; Dai, D.Y. Fauna Sinica: Osteichthyes: Siluriformes; Science Press: Beijing, China, 1999. [Google Scholar]
- Yue, P.Q.; Shan, X.H.; Lin, R. Fauna Sinica, Osteichthyes, Cypriniformes III; Science Press: Beijing, China, 2000. [Google Scholar]
- Bista, I.; Carvalho, G.R.; Walsh, K.; Seymour, M.; Hajibabaei, M.; Lallias, D.; Christmas, M.; Creer, S. Annual time-series analysis of aqueous eDNA reveals ecologically relevant dynamics of lake ecosystem biodiversity. Nat. Commun. 2017, 8, 14087. [Google Scholar] [CrossRef] [PubMed]
- Miya, M.; Sato, Y.; Fukunaga, T.; Sado, T.; Poulsen, J.Y.; Sato, K.; Minamoto, T.; Yamamoto, S.; Yamanaka, H.; Araki, H.; et al. MiFish, a set of universal PCR primers for metabarcoding environmental DNA from fishes: Detection of more than 230 subtropical marine species. R. Soc. Open Sci. 2015, 2, 150088. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Wen, Y.; Liu, X.; Yu, J.; Jin, B.; Li, D. Impact of anthropogenic activities on changes of ichthyofauna in the middle and lower Xiang River. Aquac. Fish. 2022, 7, 693–702. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Quince, C.; Lanzen, A.; Davenport, R.J.; Turnbaugh, P.J. Removing noise from pyrosequenced amplicons. BMC Bioinform. 2011, 12, 38. [Google Scholar] [CrossRef]
- Majaneva, M.; Hyytiäinen, K.; Varvio, S.L.; Nagai, S.; Blomster, J. Bioinformatic amplicon read processing strategies strongly affect eukaryotic diversity and the taxonomic composition of communities. PLoS ONE 2015, 10, e0130035. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.P.; Veach, A.M.; Rigdon-Huss, A.R.; Grond, K.; Lickteig, S.K.; Lothamer, K.; Oliver, A.K.; Jumpponen, A. Scraping the bottom of the barrel: Are rare high throughput sequences artifacts? Fungal Ecol. 2015, 13, 221–225. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Qin, J.; Xu, Y.; Ouyang, S.; Wu, X. Biodiversity decline of fish assemblages after the impoundment of the Three Gorges Dam in the Yangtze River Basin, China. Rev. Fish Biol. Fish. 2019, 29, 177–195. [Google Scholar] [CrossRef]
- Lyons, J. Distribution, abundance, and mortality of small littoral-zone fishes in Sparkling Lake, Wisconsin. Environ. Biol. Fishes 1987, 18, 93–107. [Google Scholar] [CrossRef]
- Shan, X.; Jin, X.; Dai, F.; Chen, Y.; Yang, T.; Yao, J. Population dynamics of fish species in a marine ecosystem: A case study in the Bohai Sea, China. Mar. Coast. Fish. 2016, 8, 100–117. [Google Scholar] [CrossRef]
- Stoeckle, M.Y.; Adolf, J.; Charlop-Powers, Z.; Dunton, K.J.; Hinks, G.; VanMorter, S.M. Trawl and eDNA assessment of marine fish diversity, seasonality, and relative abundance in coastal New Jersey, USA. Ices J. Mar. Sci. 2021, 78, 293–304. [Google Scholar] [CrossRef]
- Li, C.; Long, H.; Yang, S.; Zhang, Y.; Tang, F.; Jin, W.; Wang, G.; Chang, W.; Pi, Y.; Gao, L.; et al. eDNA assessment of pelagic fish diversity, distribution, and abundance in the central Pacific Ocean. Reg. Stud. Mar. Sci. 2022, 56, 102661. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Pielou, E. Species-diversity and pattern-diversity in study of ecological succession. J. Theor. Biol. 1966, 10, 370–383. [Google Scholar] [CrossRef]
- Chao, A. Nonparametric-estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Chao, A.; Ma, M.; Yang, M. Stopping Rules and estimation for recapture debugging with unequal failure rates. Biometrika 1993, 80, 193–201. [Google Scholar] [CrossRef]
- Peng, W.; Shazhou, A.; Mei, K.; Chao, L.; Yurong, H.; Jiyong, L.; Shuai, K.; Junpeng, J. Effects of enclosure on plant and soil restoration in the junggar desert. J. Resour. Ecol. 2021, 12, 840–848. [Google Scholar] [CrossRef]
- Thomsen, P.F.; Moller, P.R.; Sigsgaard, E.E.; Knudsen, S.W.; Jorgensen, O.A.; Willerslev, E. Environmental DNA from seawater samples correlate with Trawl Catches of Subarctic, deepwater fishes. PLoS ONE 2016, 11, e0165252. [Google Scholar] [CrossRef]
- Bylemans, J.; Gleeson, D.M.; Lintermans, M.; Hardy, C.M.; Beitzel, M.; Gilligan, D.M.; Furlan, E.M. Monitoring riverine fish communities through eDNA metabarcoding: Determining optimal sampling strategies along an altitudinal and biodiversity gradient. Metabarcoding Metagenomics 2018, 2, e30457. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. A further biodiversity index applicable to species lists: Variation in taxonomic distinctness. Mar. Ecol. Prog. Ser. 2001, 216, 265–278. [Google Scholar] [CrossRef]
- Clarke, K. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P. R Package; Version 2.5–7; Vegan: Community Ecology Package; The R Project for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Singer, G.a.C.; Fahner, N.A.; Barnes, J.G.; McCarthy, A.; Hajibabaei, M. Comprehensive biodiversity analysis via ultra-deep patterned flow cell technology: A case study of eDNA metabarcoding seawater. Sci. Rep. 2019, 9, 5991. [Google Scholar] [CrossRef] [PubMed]
- Sard, N.M.; Herbst, S.J.; Nathan, L.; Uhrig, G.; Kanefsky, J.; Robinson, J.D.; Scribner, K.T. Comparison of fish detections, community diversity, and relative abundance using environmental DNA metabarcoding and traditional gears. Environ. DNA 2019, 1, 368–384. [Google Scholar] [CrossRef]
- Xu, S.; Lyu, P.; Zheng, X.; Yang, H.; Xia, B.; Li, H.; Zhang, H.; Ma, S. Monitoring and control methods of harmful algal blooms in Chinese freshwater system: A review. Environ. Sci. Pollut. Res. 2022, 29, 56908–56927. [Google Scholar] [CrossRef]
- Li, J.; Huang, L.; Zou, L.; Kano, Y.; Sato, T.; Yahara, T. Spatial and temporal variation of fish assemblages and their associations to habitat variables in a mountain stream of north Tiaoxi River, China. Environ. Biol. Fishes 2012, 93, 403–417. [Google Scholar] [CrossRef]
- Cahill, A.E.; Pearman, J.K.; Borja, A.; Carugati, L.; Carvalho, S.; Danovaro, R.; Dashfield, S.; David, R.; Feral, J.-P.; Olenin, S.; et al. A comparative analysis of metabarcoding and morphology-based identification of benthic communities across different regional seas. Ecol. Evol. 2018, 8, 8908–8920. [Google Scholar] [CrossRef]
- Gillet, B.; Cottet, M.; Destanque, T.; Kue, K.; Descloux, S.; Chanudet, V.; Hughes, S. Direct fishing and eDNA metabarcoding for biomonitoring during a 3-year survey significantly improves number of fish detected around a South East Asian reservoir. PLoS ONE 2018, 13, e0208592. [Google Scholar] [CrossRef]
- Czeglédi, I.; Sály, P.; Specziár, A.; Preiszner, B.; Szalóky, Z.; Maroda, Á.; Pont, D.; Meulenbroek, P.; Valentini, A.; Erős, T. Congruency between two traditional and eDNA-based sampling methods in characterising taxonomic and trait-based structure of fish communities and community-environment relationships in lentic environment. Ecol. Indic. 2021, 129, 107952. [Google Scholar] [CrossRef]
- Jiang, P.; Zhang, S.; Xu, S.; Xiong, P.; Cao, Y.; Chen, Z.; Li, M. Comparison of environmental DNA metabarcoding and bottom trawling for detecting seasonal fish communities and habitat preference in a highly disturbed estuary. Ecol. Indic. 2023, 146, 109754. [Google Scholar] [CrossRef]
- Belle, C.C.; Stoeckle, B.C.; Geist, J. Taxonomic and geographical representation of freshwater environmental DNA research in aquatic conservation. Aquat. Conserv.-Mar. Freshw. Ecosyst. 2019, 29, 1996–2009. [Google Scholar] [CrossRef]
- Weigand, H.; Beermann, A.J.; Ciampor, F.; Costa, F.O.; Csabai, Z.; Duarte, S.; Geiger, M.F.; Grabowski, M.; Rimet, F.; Rulik, B.; et al. DNA barcode reference libraries for the monitoring of aquatic biota in Europe: Gap-analysis and recommendations for future work. Sci. Total Environ. 2019, 678, 499–524. [Google Scholar] [CrossRef] [PubMed]
- Barco, A.; Kullmann, B.; Knebelsberger, T.; Sarrazin, V.; Kuhs, V.; Kreutle, A.; Pusch, C.; Thiel, R. Detection of fish species from marine protected areas of the North Sea using environmental DNA. J. Fish Biol. 2022, 101, 722–727. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, M.J.; Collie, J.S.; Hall, S.J.; Jennings, S.; Poiner, I.R. Modification of marine habitats by trawling activities: Prognosis and solutions. Fish Fish. 2002, 3, 114–136. [Google Scholar] [CrossRef]
- Braun, L.-M.; Brucet, S.; Mehner, T. Top-down and bottom-up effects on zooplankton size distribution in a deep stratified lake. Aquat. Ecol. 2021, 55, 527–543. [Google Scholar] [CrossRef]
- Cai, X.; Ye, S.; Li, W.; Fan, H.; Li, Z.; Zhang, T.; Liu, J. Spatio-temporal variability of small fishes related with environmental factors in a typical domestic tap water lake, Eastern China. J. Oceanol. Limnol. 2019, 37, 278–289. [Google Scholar] [CrossRef]
- Alam, A.; Kumar, J.; Sarkar, U.K.; Jha, D.N.; Das, S.C.S.; Srivastava, S.K.; Kumar, V.; Das, B.K. Assessing the influence of environmental factors on fish assemblage and spatial diversity in an unexplored sub-tropical Jargo reservoir of the Ganga River basin. Aquat. Ecosyst. Health Manag. 2021, 24, 46–53. [Google Scholar] [CrossRef]
- Qin, J.; Cheng, F.; Zhang, L.; Schmidt, B.V.; Liu, J.; Xie, S. Invasions of two estuarine gobiid species interactively induced from water diversion and saltwater intrusion. Manag. Biol. Invasions 2019, 10, 139–150. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Orders | Families | Species | Feeding Habits | Water Layer | The Relative Abundance (N%) of TFL | The Relative Abundance (N%) of eDNA | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| DT | DH | SWC | DT | DH | SWC | |||||
| Salmoniformes | Salangidae | Protosalanx hyalocranius * | C | UL | 0.00 | 0.00 | 11.25 | 6.97 | 4.29 | 0.01 |
| Cypriniformes | Cyprinidae | Ctenopharyngodon idella | H | LL | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.03 |
| Toxabramis swinhonis * | P | UL | 4.85 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | ||
| Hemiculter leucisculus * | P | UL | 20.84 | 16.75 | 0.00 | 39.66 | 11.50 | 0.07 | ||
| Cultrichthys erythropterus * | C | UL | 0.16 | 9.14 | 0.00 | <0.01 | <0.01 | <0.01 | ||
| Culter alburnus | C | UL | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.00 | ||
| Megalobrama amblycephala | H | LL | 0.00 | 0.00 | 0.00 | 5.33 | 3.79 | 1.68 | ||
| Pseudobrama simoni * | P | UL | 13.09 | 2.79 | 1.25 | 0.00 | 0.00 | 0.00 | ||
| Acheilognathus macropterus * | O | DM | 0.81 | 39.59 | 3.75 | 0.00 | <0.01 | 0.00 | ||
| Acheilognathus chankaensis * | O | DM | 0.00 | 0.00 | 0.00 | 0.01 | 13.77 | <0.01 | ||
| Paracheilognathus imberbis * | O | DM | 0.81 | 2.03 | 1.25 | 0.00 | 0.00 | 0.00 | ||
| Hemibarbus maculatus | O | DM | 0.65 | 1.52 | 2.50 | 0.00 | 0.00 | 0.00 | ||
| Pseudorasbora parva * | O | DM | 9.53 | 1.52 | 0.00 | 0.50 | 2.61 | 0.02 | ||
| Sarcocheilichthys nigripinnis * | O | DM | 0.81 | 1.52 | 0.00 | <0.01 | 0.04 | 0.01 | ||
| Abbottina rivularis * | O | DM | 16.48 | 0.00 | 53.75 | 0.00 | 0.00 | 0.00 | ||
| Cyprinus carpio | O | DM | 0.00 | 0.00 | 0.00 | 0.01 | 11.97 | 0.01 | ||
| Carassius auratus * | O | DM | 0.81 | 7.61 | 0.00 | 1.64 | 0.03 | 24.02 | ||
| Aristichthys nobilis | P | UL | 0.00 | 2.54 | 0.00 | 0.03 | 12.28 | 0.03 | ||
| Hypophthalmichthys molitrix | P | UL | 0.00 | 1.02 | 0.00 | 3.04 | 0.04 | 0.00 | ||
| Siluriformes | Siluridae | Silurus asotus | C | DM | 0.00 | 1.52 | 6.25 | 0.00 | 0.00 | 0.00 |
| Bagridae | Pelteobagrus fulvidraco * | C | DM | 0.16 | 1.27 | 3.75 | 6.48 | 4.49 | <0.01 | |
| Beloniformes | Hemiramphidae | Hyporhamphus intermedius * | P | UL | 0.00 | 0.00 | 0.00 | <0.01 | 4.41 | 0.05 |
| Perciformes | Gobiidae | Tridentiger bifasciatus * | C | DM | 28.76 | 11.17 | 11.25 | 32.19 | 28.25 | 0.03 |
| Rhinogobius giurinus * | C | DM | 2.26 | 0.00 | 2.50 | 4.07 | 2.52 | 47.57 | ||
| Rhinogobius cliffordpopei * | C | DM | 0.00 | 0.00 | 0.00 | 0.05 | <0.01 | 23.38 | ||
| Channidae | Channa argus | C | LL | 0.00 | 0.00 | 2.50 | 0.02 | <0.01 | 3.07 | |
| Sampling Reservoirs | Dominant Species | IRI (×103) | N (%) | W (%) | F (%) |
|---|---|---|---|---|---|
| DT | Pseudobrama simoni | 5.71 | 13.09 | 44.05 | 100.00 |
| Tridentiger bifasciatus | 3.61 | 28.76 | 7.33 | 100.00 | |
| Hemiculter leucisculus | 2.45 | 20.84 | 3.70 | 100.00 | |
| Abbottina rivularis | 2.08 | 16.48 | 4.30 | 100.00 | |
| Hemibarbus maculatus | 1.35 | 0.65 | 19.66 | 66.67 | |
| Pseudorasbora parva | 1.12 | 9.53 | 1.68 | 100.00 | |
| DH | Acheilognathus macropterus | 5.14 | 39.59 | 11.76 | 100.00 |
| Cultrichthys erythropterus | 4.35 | 9.14 | 34.32 | 100.00 | |
| Hemiculter leucisculus | 3.65 | 16.75 | 19.74 | 100.00 | |
| Carassius auratus | 1.43 | 7.61 | 13.89 | 66.67 | |
| Tridentiger bifasciatus | 1.26 | 11.17 | 1.43 | 100.00 | |
| SWC | Silurus asotus | 4.04 | 6.25 | 54.29 | 66.67 |
| Abbottina rivularis | 3.85 | 53.75 | 4.05 | 66.67 | |
| Protosalanx hyalocranius | 1.16 | 11.25 | 0.34 | 100.00 | |
| Channa argus | 1.16 | 2.50 | 32.21 | 33.33 |
| Sampling Reservoirs | Shannon–Wiener Index | Pielou’s Index | Chao1 Index | ACE Index | ||||
|---|---|---|---|---|---|---|---|---|
| TFL | eDNA | TFL | eDNA | TFL | eDNA | TFL | eDNA | |
| DT | 1.91 | 1.59 | 0.73 | 0.55 | 15.00 | 19.00 | 14.74 | 20.04 |
| DH | 1.94 | 2.12 | 0.74 | 0.74 | 14.00 | 18.00 | 14.00 | 18.00 |
| SWC | 1.63 | 1.23 | 0.68 | 0.44 | 11.25 | 17.00 | 12.51 | 17.53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, H.; Chen, S.; Liu, X.; Cheng, Z.; Schmidt, B.V.; He, W.; Cheng, F.; Xie, S. Investigations of Fish Assemblages Using Two Methods in Three Terminal Reservoirs of the East Route of South-to-North Water Transfer Project, China. Animals 2023, 13, 1614. https://doi.org/10.3390/ani13101614
Yan H, Chen S, Liu X, Cheng Z, Schmidt BV, He W, Cheng F, Xie S. Investigations of Fish Assemblages Using Two Methods in Three Terminal Reservoirs of the East Route of South-to-North Water Transfer Project, China. Animals. 2023; 13(10):1614. https://doi.org/10.3390/ani13101614
Chicago/Turabian StyleYan, Huiguo, Sibao Chen, Xia Liu, Zhenhao Cheng, Bjorn Victor Schmidt, Wenping He, Fei Cheng, and Songguang Xie. 2023. "Investigations of Fish Assemblages Using Two Methods in Three Terminal Reservoirs of the East Route of South-to-North Water Transfer Project, China" Animals 13, no. 10: 1614. https://doi.org/10.3390/ani13101614