
Integrated Transcriptomic and Metabolomic Analyses Reveal Low-Temperature Tolerance Mechanism in Giant Freshwater Prawn Macrobrachium rosenbergii
 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Low-Temperature Stress Treatment and Sampling
2.2. Transcriptomic Analysis
2.2.1. Total RNA Extraction, Library Preparation, and Sequencing
2.2.2. Transcript Assembly and Annotation
2.2.3. Differentially Expressed Gene Analysis
2.3. Metabolomic Analysis
2.3.1. Extraction of Metabolites and LC-MS Analysis
2.3.2. Metabolome Data Processing
2.4. Combination of Transcriptomic and Metabolomic Analyses
2.5. Validation of Selected Genes by RT-qPCR
2.6. Statistics Analysis
3. Results
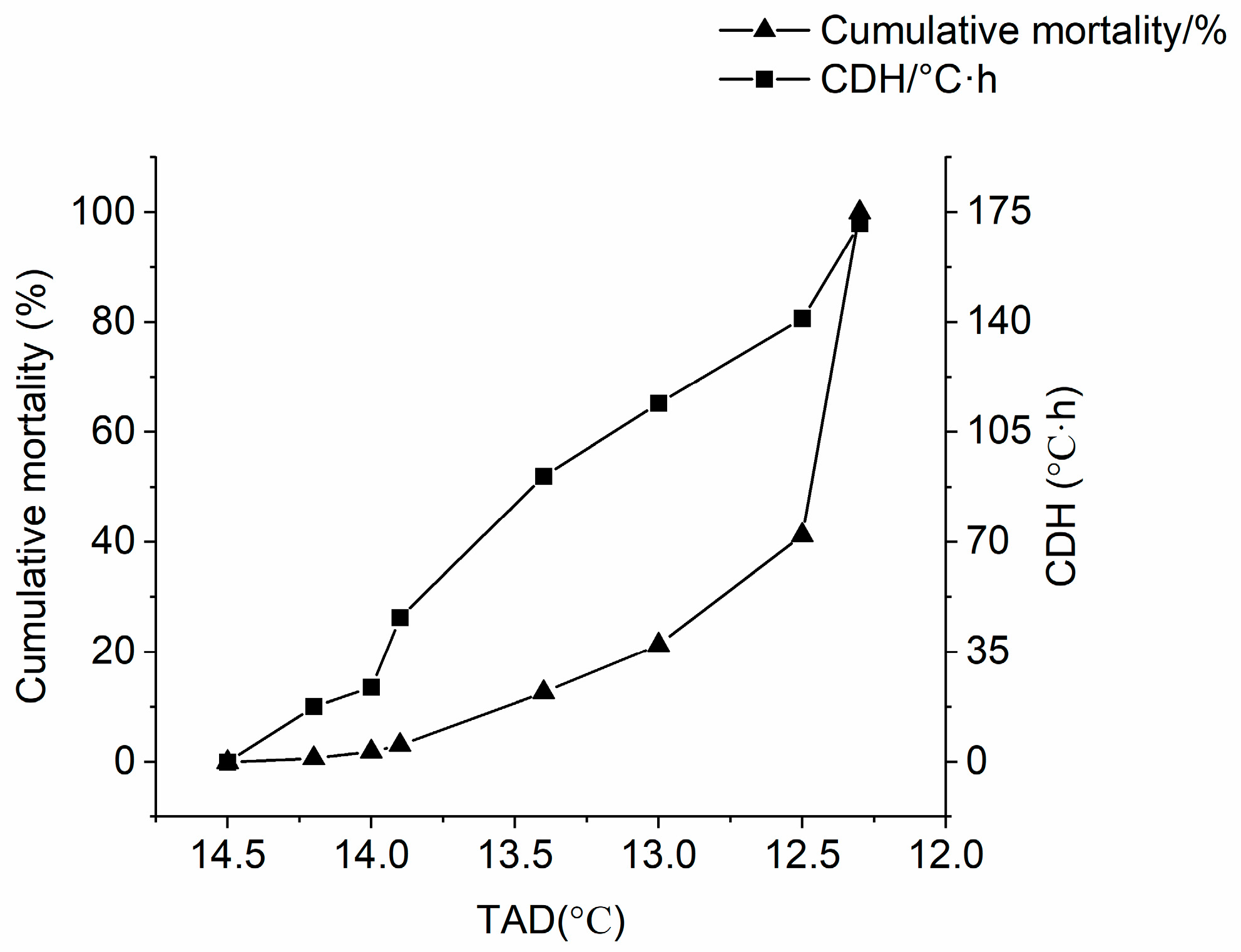
3.1. Evaluation of Low-Temperature Tolerance
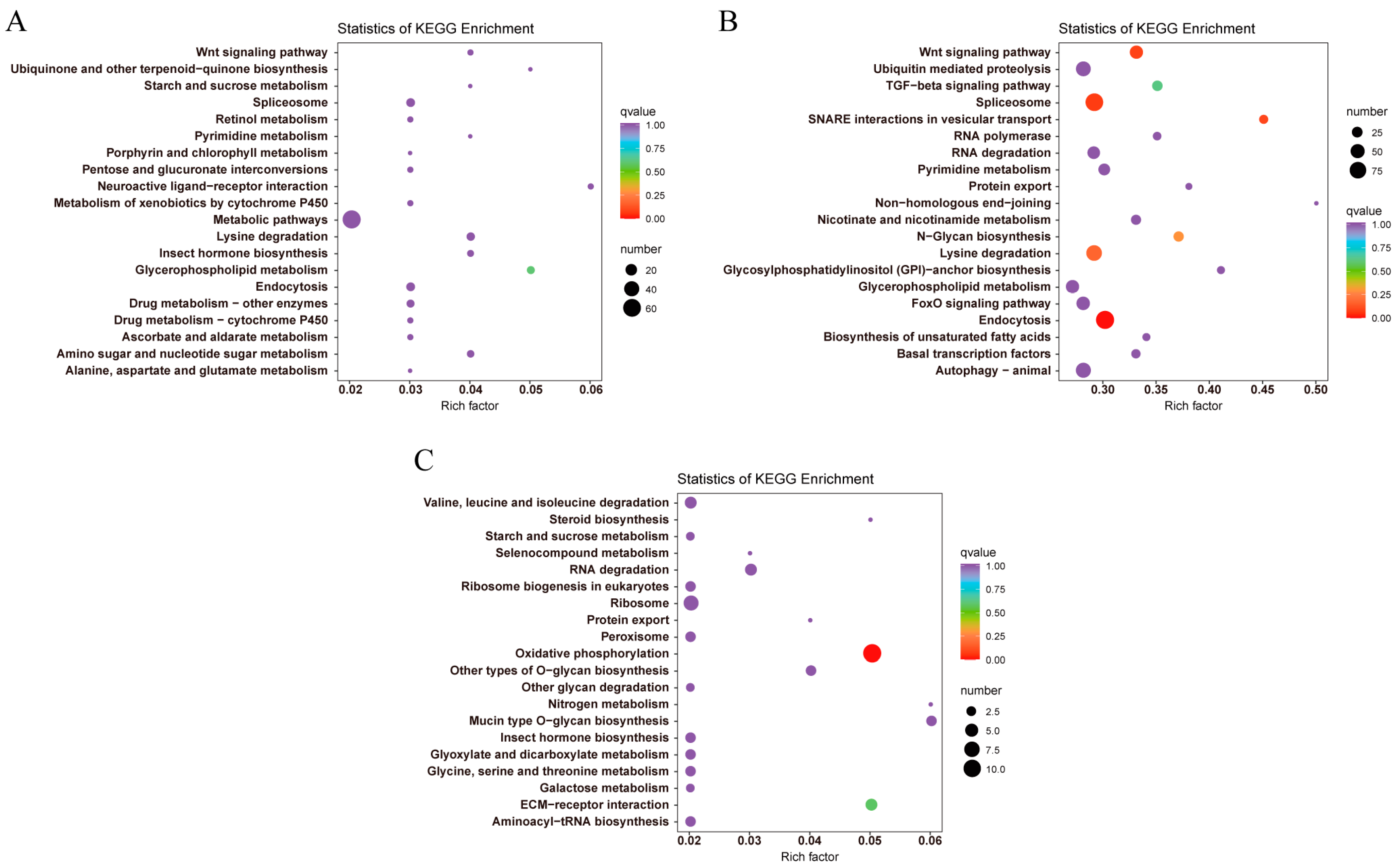
3.2. Identification, GO, and KEGG Classification of DEGs under Low-Temperature Stress
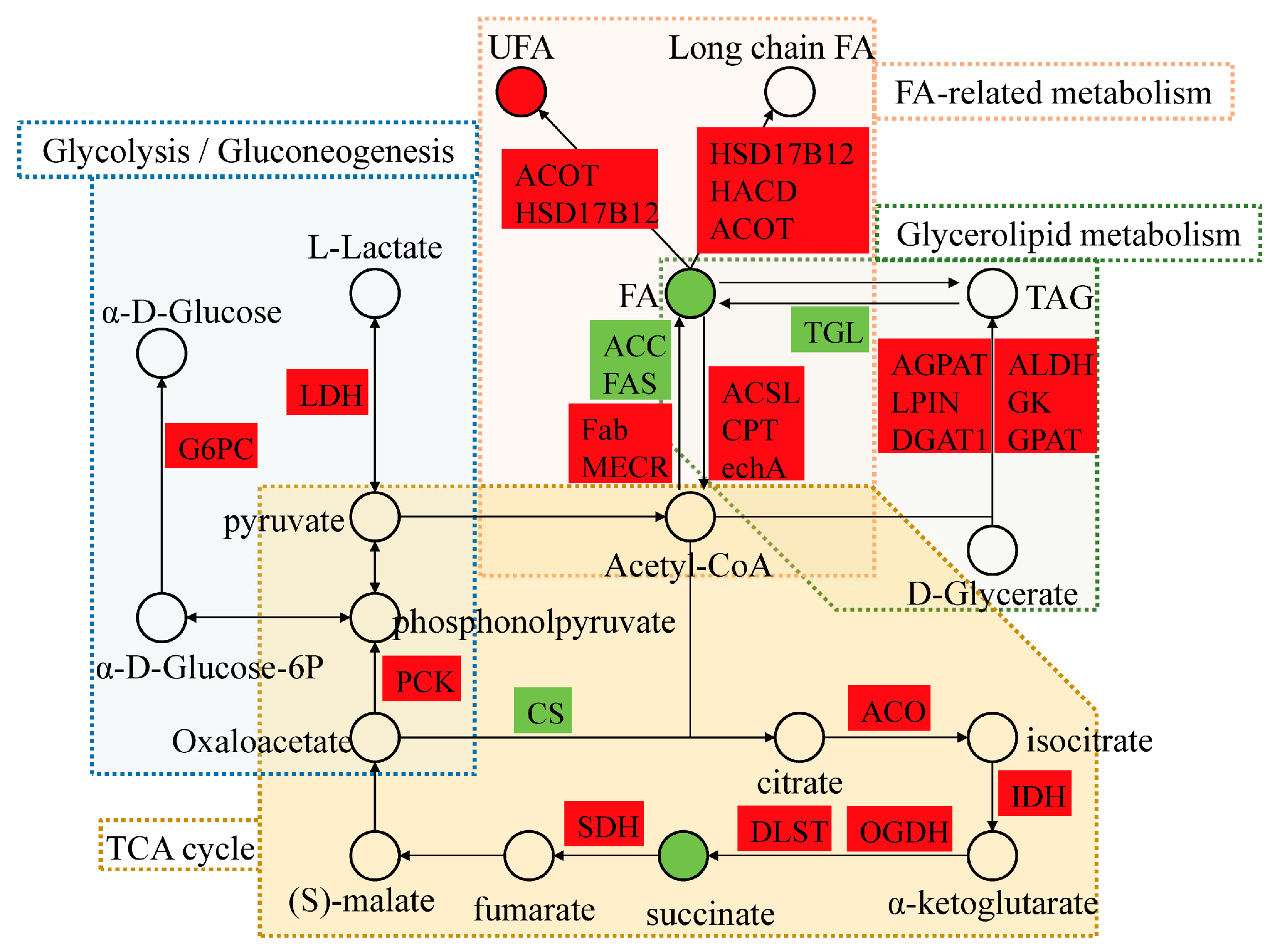
3.3. DEGs Related to Lipid and Energy Metabolism
3.4. Dynamics of Metabolites under Low-Temperature Stress
3.5. Differential Metabolites Related to Lipid and Energy Metabolism
3.6. Correlation Analysis of Transcriptomic and Metabolomic Data
3.7. Validation of Significant DEGs by qRT-PCR
4. Discussion
4.1. Alterations of Lipid Metabolism Associated with Low-Temperature Stress
4.2. Low-Temperature Stress-Induced Energy Metabolism of GFP
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- New, M.B.; Valenti, W.C.; Tidwell, J.H.; D’Abramo, L.R.; Kutty, M.N. Freshwater Prawns Biology and Farming; Wiley-Blackwell: West Sussex County, UK, 2010; pp. 1–528. [Google Scholar]
- FAO. Fisheries and Aquaculture Software Fishstatj-Software for Fishery and Aquaculture Statistical Time Series. Available online: http://www.fao.org/fishery/statistics/software/fishstatj/en (accessed on 18 November 2021).
- New, M.B. Freshwater prawn culture: A review. Aquaculture 1990, 88, 99–143. [Google Scholar] [CrossRef]
- Xing, Q.; Tu, H.; Yang, M.; Chen, G.; Tang, Q.; Yi, S.; Gao, Q.; Ibrahim, S.; Liu, Y.; Xia, Z.; et al. Evaluation of cold tolerance and gene expression patterns associated with low-temperature stress in giant freshwater prawn Macrobrachium rosenbergii. Aquac. Rep. 2022, 24, 101172. [Google Scholar] [CrossRef]
- Xu, Z.; Regenstein, J.M.; Xie, D.; Lu, W.; Ren, X.; Yuan, J.; Mao, L. The oxidative stress and antioxidant responses of Litopenaeus vannamei to low temperature and air exposure. Fish Shellfish Immunol. 2018, 72, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Wang, W.N.; Wang, L.J.; Liu, Y.F.; Wang, A.L. Oxidative stress, DNA damage and osmolality in the Pacific white shrimp, Litopenaeus vannamei exposed to acute low temperature stress. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 154, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Huang, M.; Zhou, Y.; Liu, C.; Han, C.; Gao, Q.; Dong, Y.; Dong, S. Effects of different temperatures on seawater acclimation in rainbow trout Oncorhynchus mykiss: Osmoregulation and branchial phospholipid fatty acid composition. J. Comp. Physiol. B 2021, 191, 669–679. [Google Scholar] [CrossRef]
- Lin, F.; Lin, J.; Liu, X.; Yuan, Y.; Liu, G.; Ye, X. Effects of temperature on muscle growth and collagen deposition in zebrafish (Danio rerio). Aquac. Rep. 2022, 22, 100952. [Google Scholar] [CrossRef]
- Hsu, C.Y.; Chiu, Y.C. Ambient temperature influences aging in an annual fish (Nothobranchius rachovii). Aging Cell 2009, 8, 726–737. [Google Scholar] [CrossRef]
- Cheng, C.H.; Liang, H.Y.; Luo, S.W.; Wang, A.L.; Ye, C.X. The protective effects of vitamin C on apoptosis, DNA damage and proteome of pufferfish (Takifugu obscurus) under low temperature stress. J. Therm. Biol. 2018, 71, 128–135. [Google Scholar] [CrossRef]
- Sun, Z.; Tan, X.; Liu, Q.; Ye, H.; Zou, C.; Xu, M.; Zhang, Y.; Ye, C. Physiological, immune responses and liver lipid metabolism of orange-spotted grouper (Epinephelus coioides) under cold stress. Aquaculture 2019, 498, 545–555. [Google Scholar] [CrossRef]
- Wen, X.; Hu, Y.; Zhang, X.; Wei, X.; Wang, T.; Yin, S. Integrated application of multi-omics provides insights into cold stress responses in pufferfish Takifugu fasciatus. BMC Genom. 2019, 20, 563. [Google Scholar] [CrossRef]
- Wu, D.; Huang, Y.; Chen, Q.; Jiang, Q.; Li, Y.; Zhao, Y. Effects and transcriptional responses in the hepatopancreas of red claw crayfish Cherax quadricarinatus under cold stress. J. Therm. Biol. 2019, 85, 102404. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhou, Y.L.; Guo, X.F.; Wei, W.Y.; Li, Z.; Zhou, L.; Wang, Z.W.; Gui, J.F. Comparative transcriptomes and metabolomes reveal different tolerance mechanisms to cold stress in two different catfish species. Aquaculture 2022, 560, 738543. [Google Scholar] [CrossRef]
- Ren, X.; Yu, Z.; Xu, Y.; Zhang, Y.; Mu, C.; Liu, P.; Li, J. Integrated transcriptomic and metabolomic responses in the hepatopancreas of kuruma shrimp (Marsupenaeus japonicus) under cold stress. Ecotoxicol. Environ. Saf. 2020, 206, 111360. [Google Scholar] [CrossRef]
- Xiang, Q.Q.; Yan, H.; Luo, X.W.; Kang, Y.H.; Hu, J.M.; Chen, L.Q. Integration of transcriptomics and metabolomics reveals damage and recovery mechanisms of fish gills in response to nanosilver exposure. Aquat. Toxicol. 2021, 237, 105895. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Davidson, N.M.; Oshlack, A. Corset: Enabling differential gene expression analysis for de novo assembled transcriptomes. Genome Biol. 2014, 15, 410. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Prakash, A.; Jeffryes, M.; Bateman, A.; Finn, R.D. The HMMER web server for protein sequence similarity search. Curr. Protoc. Bioinform. 2017, 60, 3–15. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from rna-seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Hugo, V.; Loraine, B.G.; Coppée, J.Y.; Dillies, M.A. SARTools: A DESeq2- and EdgeR-based r pipeline for comprehensive differential analysis of RNA-seq data. PLoS ONE 2016, 11, e0157022. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Kessner, D.; Chambers, M.; Burke, R.; Agus, D.; Mallick, P. ProteoWizard: Open source software for rapid proteomics tools development. Bioinformatics 2008, 24, 2534–2536. [Google Scholar] [CrossRef] [PubMed]
- Tautenhahn, R.; Patti, G.J.; Rinehart, D.; Siuzdak, G. XCMS Online: A web-based platform to process untargeted metabolomic data. Anal. Chem. 2012, 84, 5035–5039. [Google Scholar] [CrossRef] [PubMed]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef] [PubMed]
- Allen, F.; Pon, A.; Wilson, M.; Greiner, R.; Wishart, D. CFM-ID: A web server for annotation, spectrum prediction and metabolite identification from tandem mass spectra. Nucleic Acids Res. 2014, 42, W94–W99. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Wang, R.; Xiong, X.; Yin, Y.; Cai, Y.; Ma, Z.; Liu, N.; Zhu, Z.J. Metabolic reaction network-based recursive metabolite annotation for untargeted metabolomics. Nat. Commun. 2019, 10, 1516. [Google Scholar] [CrossRef]
- Kohl, M.; Wiese, S.; Warscheid, B. Cytoscape: Software for visualization and analysis of biological networks. Methods Mol. Biol. 2011, 696, 291–303. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Cnaani, A.; Gall, G.A.E.; Hulata, G. Cold tolerance of tilapia species and hybrids. Aquac. Int. 2000, 8, 289–298. [Google Scholar] [CrossRef]
- Lu, Q.X.; Lin, Y.; Bin, S.Y.; Tang, Z.S.; Peng, T. Analysis of cold tolerance measure at low-temperature in six tilapia families. J. Guangxi Norm. Univ. Nat. Sci. Ed. 2011, 29, 104–109. [Google Scholar] [CrossRef]
- Xu, D.; Zheng, X.; Li, C.; Wu, J.; Sun, L.; Qin, X.; Fan, X. Insights into the response mechanism of Litopenaeus vannamei exposed to cold stress during live transport combining untargeted metabolomics and biochemical assays. J. Therm. Biol. 2022, 104, 103200. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, W.; Qiao, H.; Jiang, S.; Xiong, Y.; Jin, S.; Gong, Y.; Fu, H. Integrated metabolomics and transcriptomic analysis of hepatopancreas in different living status Macrobrachium nipponense in response to hypoxia. Antioxidants 2021, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.C.; Kang, C.K.; Tang, C.H.; Lee, T.H. Transcriptomic analysis of metabolic pathways in milkfish that respond to salinity and temperature changes. PLoS ONE 2015, 10, e0134959. [Google Scholar] [CrossRef]
- Fahy, E.; Subramaniam, S.; Brown, H.A.; Glass, C.K.; Merrill, A.H., Jr.; Murphy, R.C.; Raetz, C.R.; Russell, D.W.; Seyama, Y.; Shaw, W.; et al. A comprehensive classification system for lipids. J. Lipid Res. 2005, 46, 839–861. [Google Scholar] [CrossRef]
- Sargent, J.R.; Tocher, D.R.; Bell, J.G. The lipids. Fish Nutr. 2003, 3, 181–257. [Google Scholar] [CrossRef]
- Chu, P.; Wang, T.; Sun, Y.R.; Chu, M.X.; Wang, H.Y.; Zheng, X.; Yin, S. Effect of cold stress on the MAPK pathway and lipidomics on muscle of Takifugu fasciatus. Aquaculture 2021, 540, 736691. [Google Scholar] [CrossRef]
- Wakil, S.J.; Stoops, J.K.; Joshi, V.C. Fatty acid synthesis and its regulation. Annu. Rev. Biochem. 1983, 52, 537–579. [Google Scholar] [CrossRef]
- Clarke, S.D.; Jump, D.B. Regulation of gene transcription by polyunsaturated fatty acids. Prog. Lipid Res. 1993, 32, 139–149. [Google Scholar] [CrossRef]
- Wu, D.; Liu, Z.; Yu, P.; Huang, Y.; Cai, M.; Zhang, M.; Zhao, Y. Cold stress regulates lipid metabolism via AMPK signalling in Cherax quadricarinatus. J. Therm. Biol. 2020, 92, 102693. [Google Scholar] [CrossRef]
- Hazel, J.R.; Williams, E.E. The role of alterations in membrane lipid composition in enabling physiological adaptation of organisms to their physical environment. Prog. Lipid Res. 1990, 29, 167–227. [Google Scholar] [CrossRef]
- Johnston, P.V.; Roots, B.I. Brain lipid fatty acids and temperature acclimation. Comp. Biochem. Physiol. 1964, 11, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Wijekoon, M.P.A. Effect of Water Temperature and Diet on Cell Membrane Fluidity and Fatty Acid Composition of Muscle, Liver, Gill and Intestine Mucosa of Adult and Juvenile Steelhead Trout, Oncorhynchus mykiss. Ph.D. Thesis, Memorial University of Newfoundland, St. John’s, NL, Canada, 2012. [Google Scholar]
- Hazel, J.R. Thermal adaptation in biological membranes: Is homeoviscousadaptation the explanation. Annu. Rev. Physiol. 1995, 57, 19–42. [Google Scholar] [CrossRef] [PubMed]
- Mejri, S.C.; Tremblay, R.; Audet, C.; Wills, P.S.; Riche, M. Essential fatty acid requirements in tropical and cold-water marine fish larvae and juveniles. Front. Mar. Sci. 2021, 8, 680003. [Google Scholar] [CrossRef]
- Ibarz, A.; Blasco, J.; Beltrán, M.; Gallardo, M.A.; Sánchez, J.; Sala, R.; Fernández-Borràs, J. Cold-induced alterations on proximate composition and fatty acid profiles of several tissues in gilthead sea bream (Sparus aurata). Aquaculture 2005, 249, 477–486. [Google Scholar] [CrossRef]
- Parrish, C.C. Essential fatty acids in aquatic food webs. In Lipids in Aquatic Ecosystems; Michael, T.A., Bruce, C.W., Eds.; Springer: New York, NY, USA, 2009; pp. 309–326. [Google Scholar]
- Rayan, P.; Stenzel, D.; McDonnell, P.A. The effects of saturated fatty acids on Giardia duodenalis trophozoites in vitro. Parasitol. Res. 2005, 97, 191–200. [Google Scholar] [CrossRef]
- Hall, L.M.; Murphy, R.C. Activation of human polymorphonuclear leukocytes by products derived from the peroxidation of human red blood cell membranes. Chem. Res. Toxicol. 1998, 11, 1024–1031. [Google Scholar] [CrossRef]
- Powell, W.S.; Gravelle, F.; Gravel, S. Metabolism of 5(S)-hydroxy-6,8,11,14-eicosatetraenoic acid and other 5(S)-hydroxyeicosanoids by a specific dehydrogenase in human polymorphonuclear leukocytes. J. Biol. Chem. 1992, 267, 19233–19241. [Google Scholar] [CrossRef]
- de Sousa Abreu, R.; Penalva, L.O.; Marcotte, E.M.; Vogel, C. Global signatures of protein and mRNA expression levels. Mol. Biosyst. 2009, 5, 1512–1526. [Google Scholar] [CrossRef]
- Jawad, L.A. Sparidae: Biology and Aquaculture of Gilthead Sea Bream and Other Species; Wiley-Blackwell: West Sussex County, UK, 2012; pp. 1–412. [Google Scholar] [CrossRef]
- Papa, S.; Martino, P.L.; Capitanio, G.; Gaballo, A.; De Rasmo, D.; Signorile, A.; Petruzzella, V. The oxidative phosphorylation system in mammalian mitochondria. Adv. Exp. Med. Biol. 2012, 942, 3–37. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Y.; Zheng, Z.; Deng, Y.; Jiao, Y.; Du, X. Adaptive response of pearl oyster Pinctada fucata martensii to low water temperature stress. Fish Shellfish Immunol. 2018, 78, 310–315. [Google Scholar] [CrossRef]
- Jungermann, K. Metabolic zonation of liver parenchyma. Semin. Liver Dis. 1988, 8, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Shohet, R.V.; Garcia, J.A. Keeping the engine primed: HIF factors as key regulators of cardiac metabolism and angiogenesis during ischemia. J. Mol. Med. 2007, 85, 1309–1315. [Google Scholar] [CrossRef] [PubMed]
- Kerner, J.; Hoppel, C. Fatty acid import into mitochondria. Biochim. Biophys. Acta 2000, 1486, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mcgarry, J.D.; Brown, N.F. The mitochondrial carnitine palmitoyltransferase system. Eur. J. Biochem. 1997, 244, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Li, L.O.; Klett, E.L.; Coleman, R.A. Acyl-CoA synthesis, lipid metabolism and lipotoxicity. Biochim. Biophys. Acta 2010, 1801, 246–251. [Google Scholar] [CrossRef]
- Ma, F.; Zou, Y.; Ma, L.; Ma, R.; Chen, X. Evolution, characterization, and immune response function of long-chain acyl-CoA synthetase genes in rainbow trout (Oncorhynchus mykiss) under hypoxic stress. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2022, 260, 110737. [Google Scholar] [CrossRef]
- Huang, D.; Liang, H.; Zhu, J.; Ren, M.; Ge, X. Transcriptome reveals insights into hepatic nutritional metabolism and gill immune responses adapted to cold stress in genetically improved farmed tilapia (GIFT: Oreochromis niloticus). Aquac. Rep. 2022, 26, 101297. [Google Scholar] [CrossRef]
- Jie, Y.K.; Cheng, C.H.; Wang, L.C.; Ma, H.L.; Deng, Y.Q.; Liu, G.X.; Feng, J.; Guo, Z.X.; Ye, L.T. Hypoxia-induced oxidative stress and transcriptome changes in the mud crab (Scylla paramamosain). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 245, 109039. [Google Scholar] [CrossRef]
- Mendez-Lucas, A.; Duarte, J.A.; Sunny, N.E.; Satapati, S.; He, T.; Fu, X.; Bermudez, J.; Burgess, S.C.; Perales, J.C. PEPCK-M expression in mouse liver potentiates, not replaces, PEPCK-C mediated gluconeogenesis. J. Hepatol. 2013, 59, 105–113. [Google Scholar] [CrossRef]
- Rognstad, R. Rate-limiting steps in metabolic pathways. J. Biol. Chem. 1979, 254, 1875–1878. [Google Scholar] [CrossRef]
- Mizock, B.A. Alterations in carbohydrate-metabolism during stress: A review of the literature. Am. J. Med. 1995, 98, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Plaxton, W.C. The organization and regulation of plant glycolysis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 185–214. [Google Scholar] [CrossRef] [PubMed]
- Everse, J.; Kaplan, N.O. Lactate dehydrogenases: Structure and function. Adv. Enzymol. Relat. Areas Mol. Biol. 1973, 37, 61–133. [Google Scholar] [CrossRef]
- Shan, H.; Geng, Z.; Ma, S.; Wang, T. Comparative study of the key enzymes and biochemical substances involved in the energy metabolism of Pacific white shrimp, Litopenaeus vannamei, with different ammonia-N tolerances. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 221, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Hägerhäll, C. Succinate: Quinone oxidoreductases variations on a conserved theme. Biochim. Biophys. Acta 1997, 1320, 107–141. [Google Scholar] [CrossRef]
- Xu, D.; Wu, J.; Sun, L.; Qin, X.; Fan, X.; Zheng, X. Energy metabolism response of Litopenaeus vannamei to combined stress of acute cold exposure and waterless duration: Implications for physiological regulation and waterless live transport. J. Therm. Biol. 2022, 104, 103149. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Minimum | Maximum | Mean ± SD |
|---|---|---|---|
| TAD/°C | 12.30 | 14.20 | 12.54 ± 0.41 |
| CDH/°C·h | 0.00 | 171.28 | 127.39 ± 38.28 |
| BW/g | 21.62 | 88.05 | 47.73 ± 12.73 |
| BL/mm | 70.20 | 414.87 | 123.60 ± 16.90 |
| Pathway | ko ID | The Number of DEGs | ||
|---|---|---|---|---|
| LS vs. Con | LT vs. Con | LT vs. LS | ||
| Fatty acid biosynthesis | ko00061 | 0 | 8 | 0 |
| Fatty acid elongation | ko00062 | 1 | 11 | 0 |
| Fatty acid degradation | ko00071 | 0 | 18 | 1 |
| Glycerolipid metabolism | ko00561 | 3 | 15 | 0 |
| Glycerophospholipid metabolism | ko00564 | 8 | 43 | 1 |
| Pyruvate metabolism | ko00620 | 1 | 16 | 0 |
| Biosynthesis of unsaturated fatty acids | ko01040 | 1 | 13 | 0 |
| Fatty acid metabolism | ko01212 | 1 | 24 | 1 |
| Glycolysis/Gluconeogenesis | ko00010 | 3 | 18 | 0 |
| Citrate cycle (TCA cycle) | ko00020 | 1 | 9 | 0 |
| Pentose phosphate pathway | ko00030 | 2 | 10 | 0 |
| Oxidative phosphorylation | ko00190 | 3 | 50 | 11 |
| Pathway | ko ID | The Number of DMs | ||
|---|---|---|---|---|
| LS vs. Con | LT vs. Con | LT vs. LS | ||
| Fatty acid biosynthesis | ko00061 | 3 | 1 | 0 |
| Glycerophospholipid metabolism | ko00564 | 0 | 0 | 1 |
| Arachidonic acid metabolism | ko00590 | 1 | 0 | 1 |
| Linoleic acid metabolism | ko00591 | 1 | 0 | 0 |
| alpha-Linolenic acid metabolism | ko00592 | 2 | 1 | 0 |
| Pyruvate metabolism | ko00620 | 3 | 1 | 0 |
| Butanoate metabolism | ko00650 | 2 | 2 | 0 |
| Biosynthesis of unsaturated fatty acids | ko01040 | 4 | 0 | 1 |
| Regulation of lipolysis in adipocytes | ko04923 | 1 | 1 | 0 |
| Oxidative phosphorylation | ko00190 | 1 | 1 | 0 |
| Citrate cycle (TCA cycle) | ko00020 | 2 | 1 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tu, H.; Peng, X.; Yao, X.; Tang, Q.; Xia, Z.; Li, J.; Yang, G.; Yi, S. Integrated Transcriptomic and Metabolomic Analyses Reveal Low-Temperature Tolerance Mechanism in Giant Freshwater Prawn Macrobrachium rosenbergii. Animals 2023, 13, 1605. https://doi.org/10.3390/ani13101605
Tu H, Peng X, Yao X, Tang Q, Xia Z, Li J, Yang G, Yi S. Integrated Transcriptomic and Metabolomic Analyses Reveal Low-Temperature Tolerance Mechanism in Giant Freshwater Prawn Macrobrachium rosenbergii. Animals. 2023; 13(10):1605. https://doi.org/10.3390/ani13101605
Chicago/Turabian StyleTu, Haihui, Xin Peng, Xinyi Yao, Qiongying Tang, Zhenglong Xia, Jingfen Li, Guoliang Yang, and Shaokui Yi. 2023. "Integrated Transcriptomic and Metabolomic Analyses Reveal Low-Temperature Tolerance Mechanism in Giant Freshwater Prawn Macrobrachium rosenbergii" Animals 13, no. 10: 1605. https://doi.org/10.3390/ani13101605