
First Evidence of Microplastics in the Yolk and Embryos of Common Cuttlefish (Sepia officinalis) from the Central Adriatic Sea: Evaluation of Embryo and Hatchling Structural Integrity and Development

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples Collection
2.2. Ethical Statement
2.3. Histological Analysis
2.4. Microplastic Extraction Protocol
2.5. Microplastics Quantification and Identification
2.6. Quality Assurance and Control (QA/QC)
2.7. Statistical Analysis
3. Results
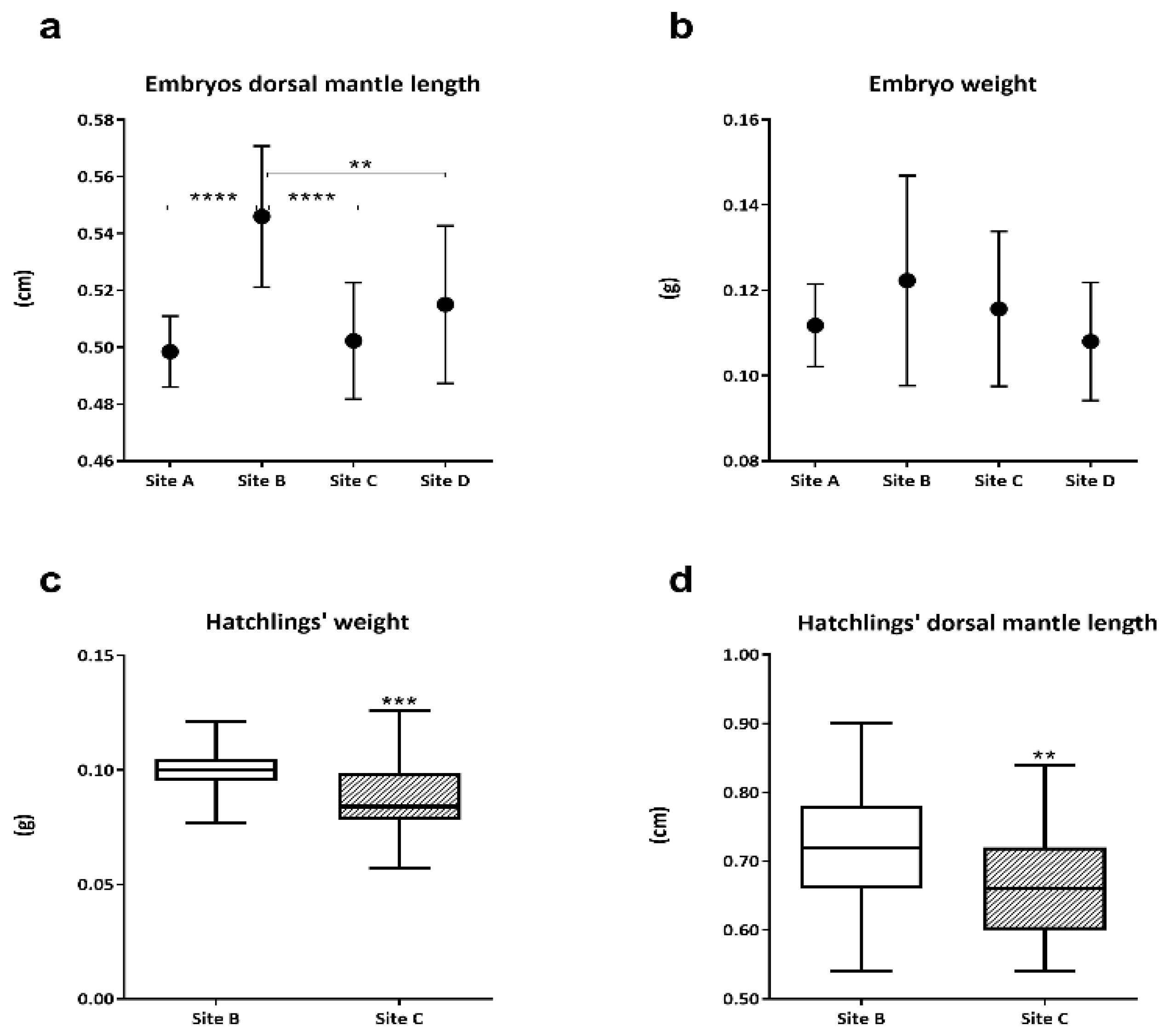
3.1. Embryos and Hatchlings Biometric Measurements
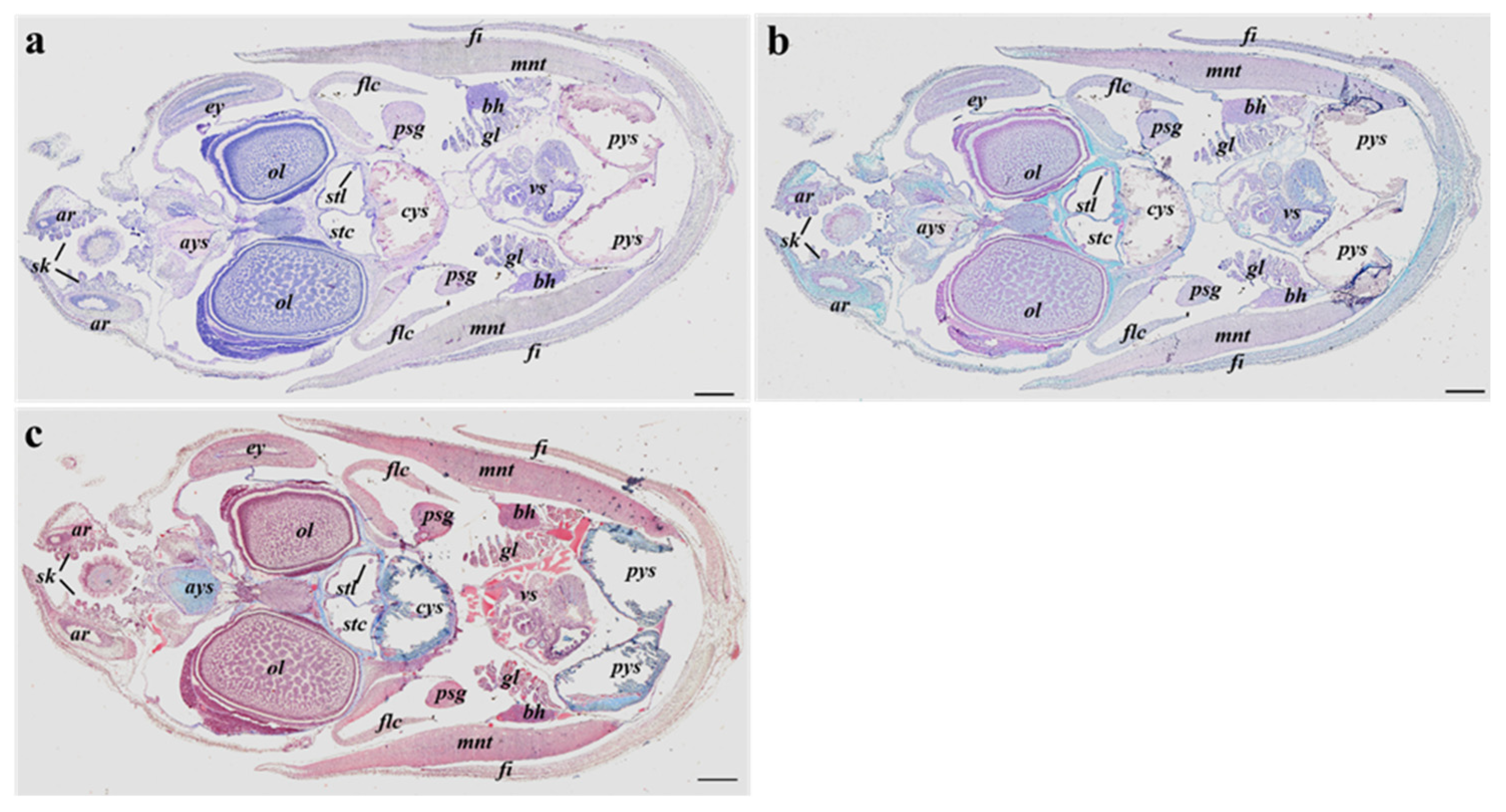
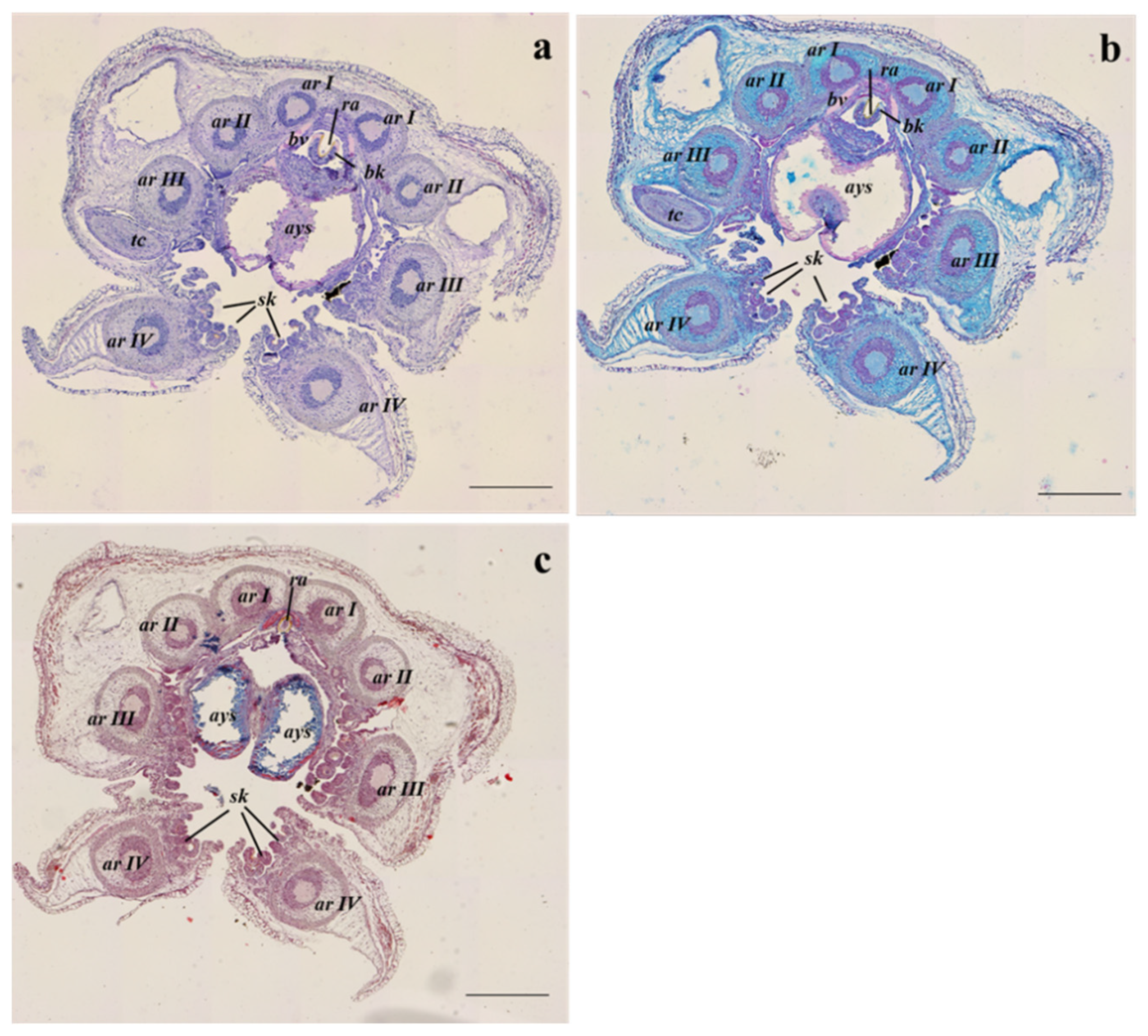
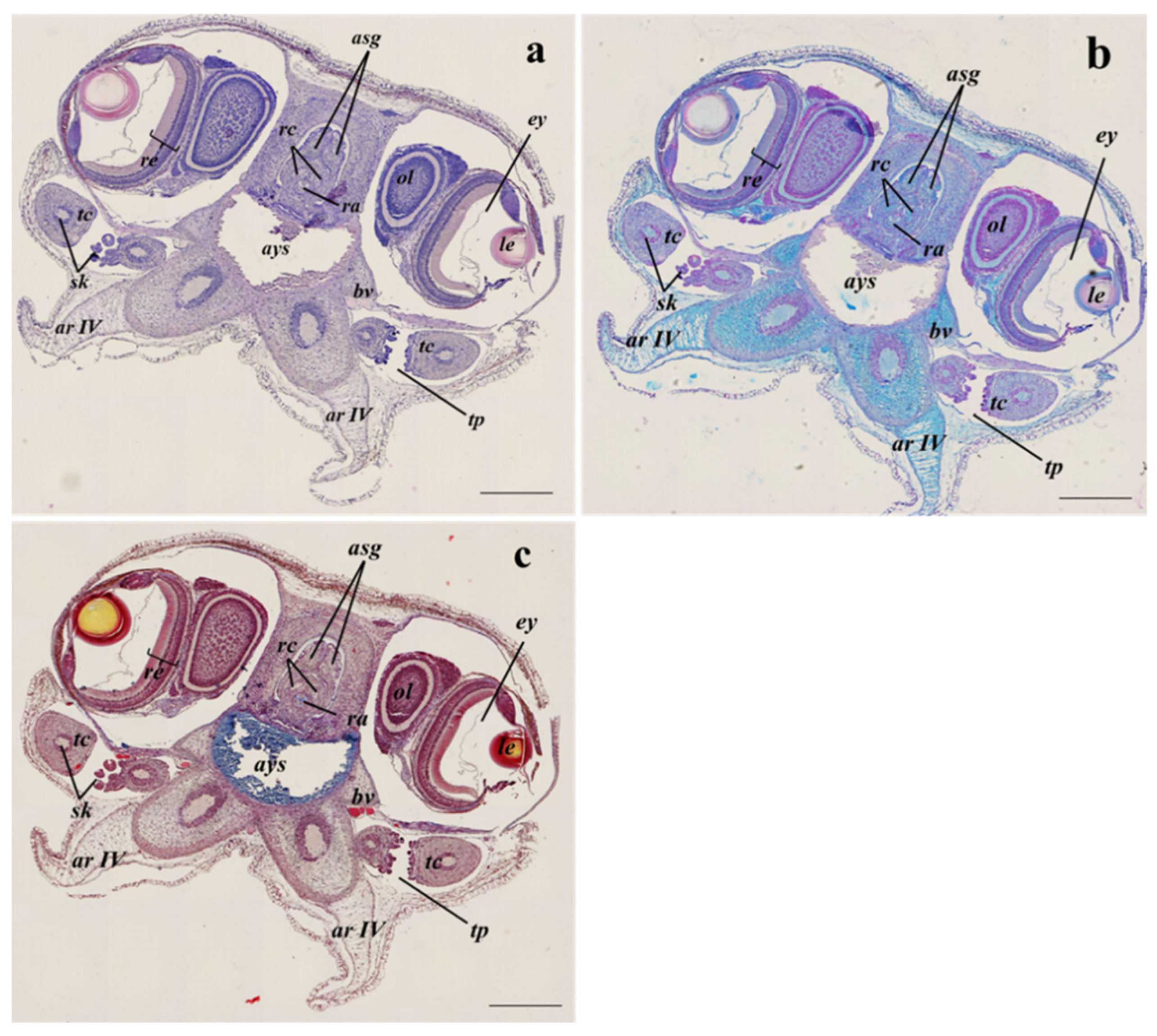
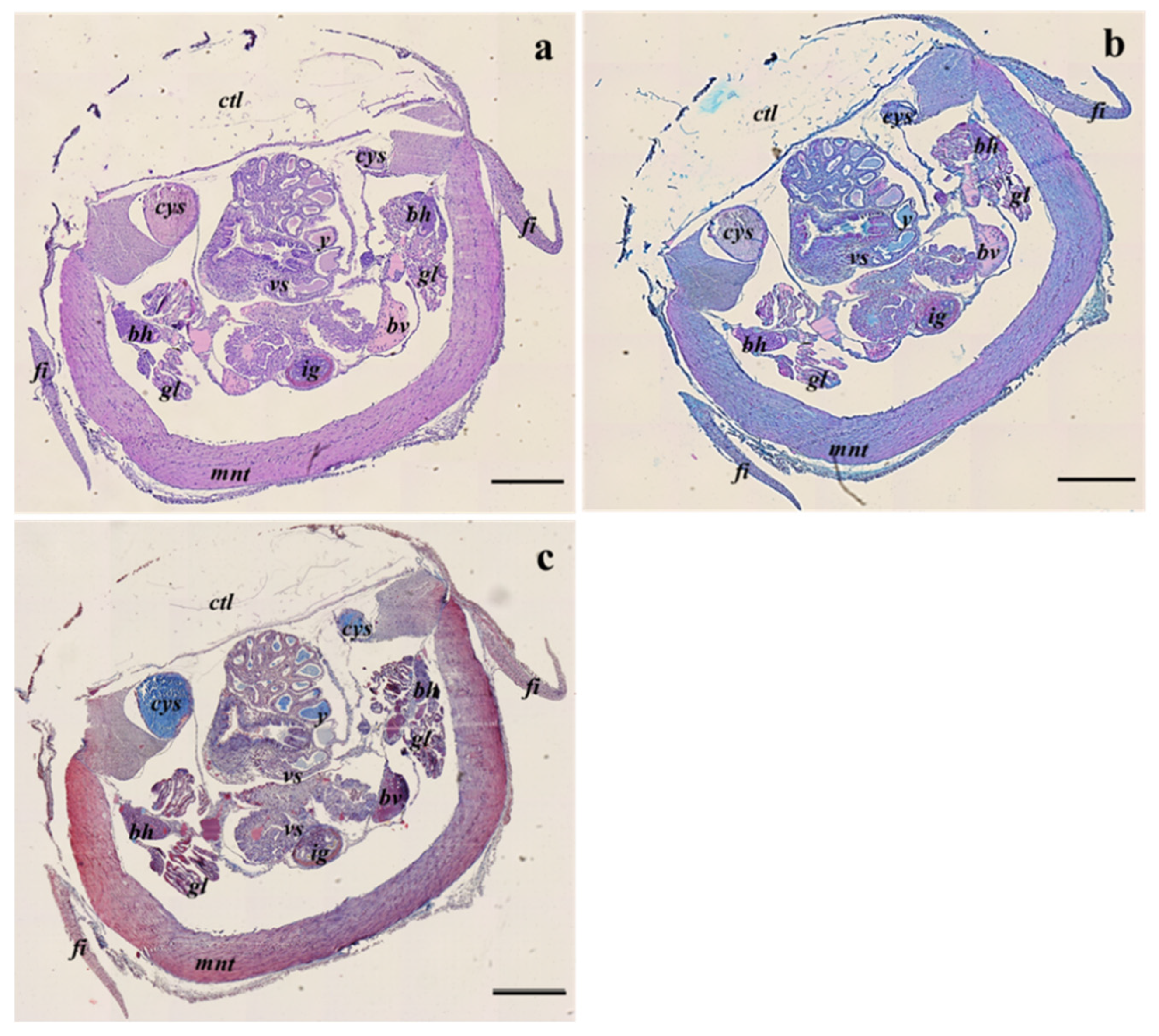
3.2. Histological Analyses
3.3. Microplastics Quantification and Identification
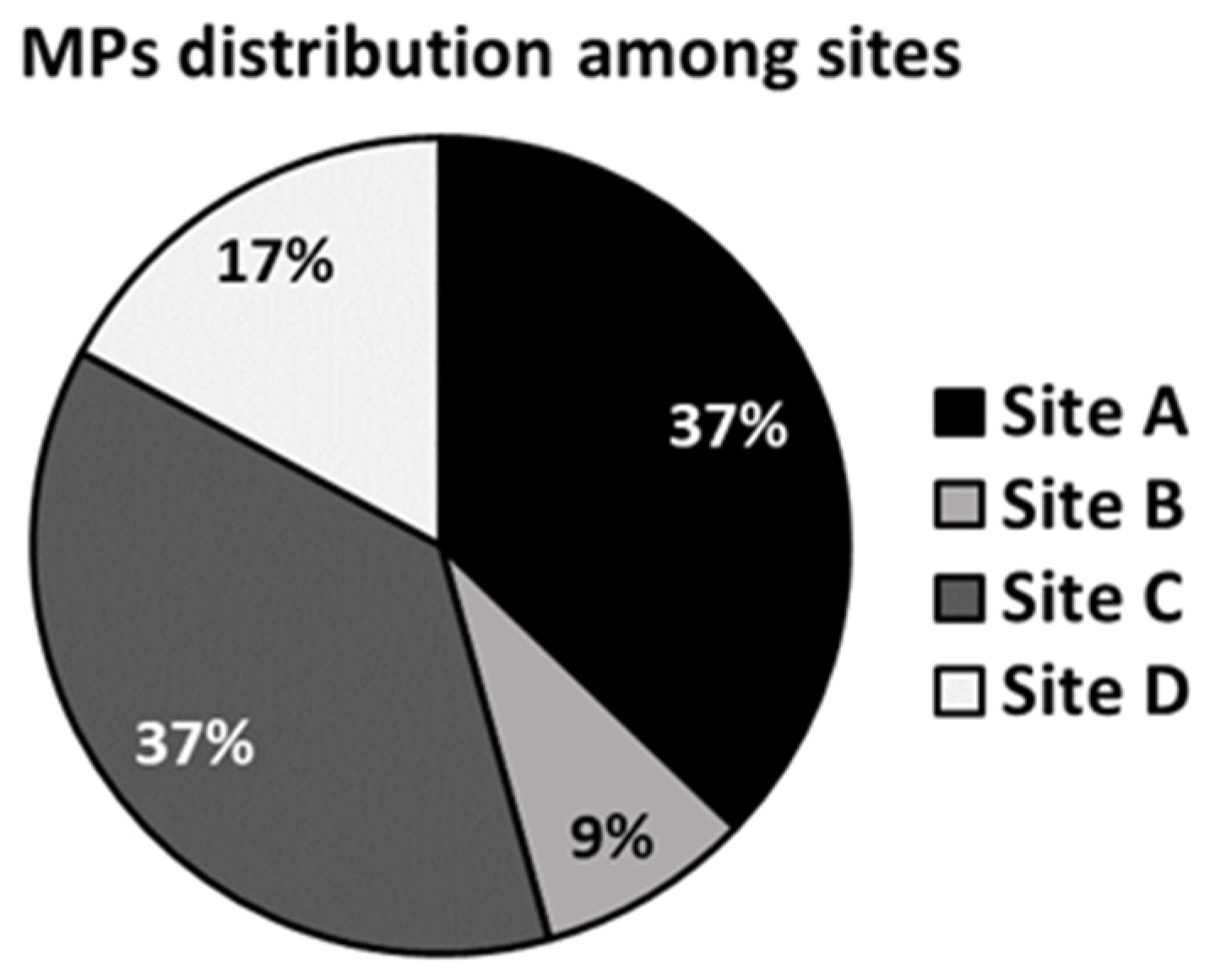
3.3.1. MP Detection in Yolk and Embryos in Each Sampling Site
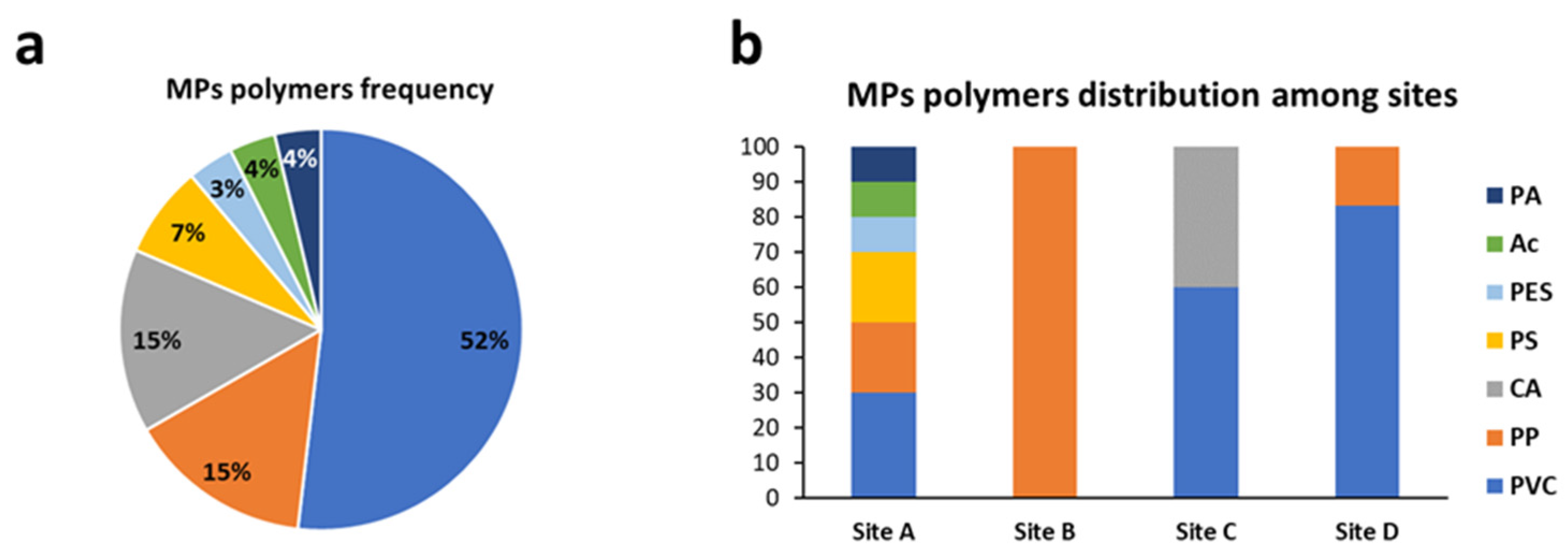
3.3.2. MP Polymers Identification in Yolk and Embryos in Each Sampling Site
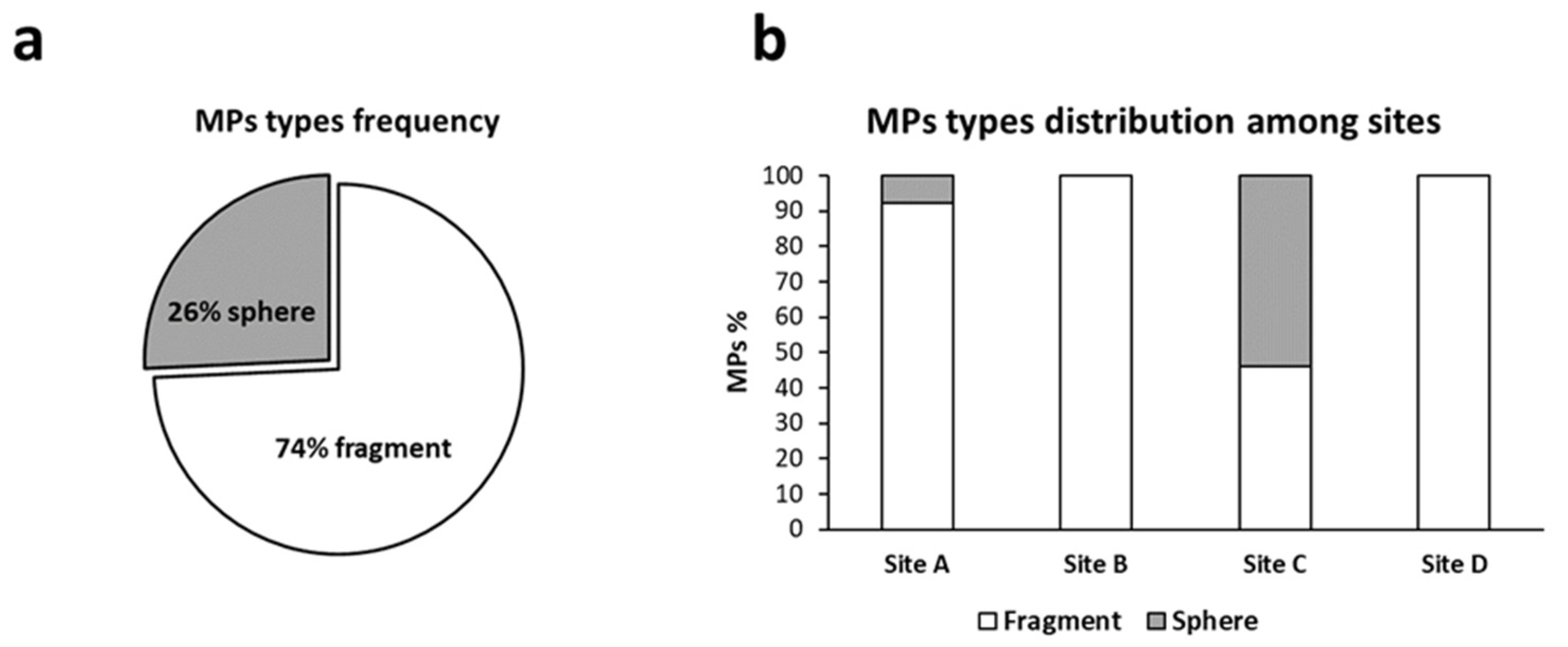
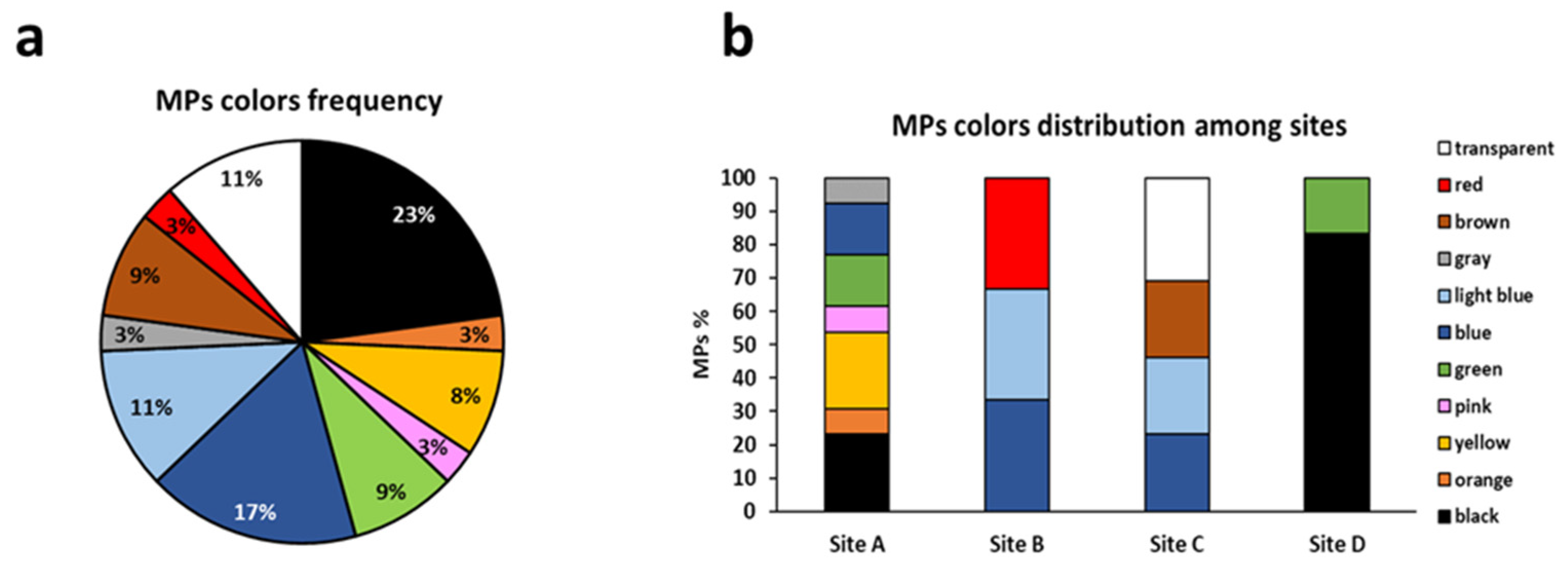
3.3.3. Total Amount of Microplastics and Distribution of Polymers and Colors among Sampling Sites
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elgarahy, A.M.; Akhdhar, A.; Elwakeel, K.Z. Microplastics prevalence, interactions, and remediation in the aquatic environment: A critical review. J. Environ. Chem. Eng. 2021, 9, 106224. [Google Scholar] [CrossRef]
- Frias, J.P.G.L.; Nash, R. Microplastics: Finding a consensus on the definition. Mar. Pollut. Bull. 2019, 138, 145–147. [Google Scholar] [CrossRef] [PubMed]
- GESAMP Sources, Fate and Effects of Microplastics in the Marine Environment (Part 2). J. Ser. GESAMP Reports Stud. 2016, 93, 221.
- Rochman, C.M. Microplastics research-from sink to source. Science 2018, 360, 28–29. [Google Scholar] [CrossRef] [PubMed]
- Zeng, E.Y. Microplastic Contamination in Aquatic Environments: An Emerging Matter of Environmental Urgency; Zeng, E.Y.E., Ed.; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Miller, M.E.; Hamann, M.; Kroon, F.J. Bioaccumulation and biomagnification of microplastics in marine organisms: A review and meta-analysis of current data. PLoS ONE 2020, 15, e0240792. [Google Scholar] [CrossRef]
- Oliveira, A.R.; Sardinha-Silva, A.; Andrews, P.L.R.; Green, D.; Cooke, G.M.; Hall, S.; Blackburn, K.; Sykes, A.V. Microplastics presence in cultured and wild-caught cuttlefish, Sepia officinalis. Mar. Pollut. Bull. 2020, 160, 111553. [Google Scholar] [CrossRef]
- Jovanović, B. Ingestion of microplastics by fish and its potential consequences from a physical perspective. Integr. Environ. Assess. Manag. 2017, 13, 510–515. [Google Scholar] [CrossRef]
- Amélineau, F.; Bonnet, D.; Heitz, O.; Mortreux, V.; Harding, A.M.A.; Karnovsky, N.; Walkusz, W.; Fort, J.; Grémillet, D. Microplastic pollution in the Greenland Sea: Background levels and selective contamination of planktivorous diving seabirds. Environ. Pollut. 2016, 219, 1131–1139. [Google Scholar] [CrossRef]
- Abidli, S.; Lahbib, Y.; Trigui El Menif, N. Microplastics in commercial molluscs from the lagoon of Bizerte (Northern Tunisia). Mar. Pollut. Bull. 2019, 142, 243–252. [Google Scholar] [CrossRef]
- Wan, Z.; Wang, C.; Zhou, J.; Shen, M.; Wang, X.; Fu, Z.; Jin, Y. Effects of polystyrene microplastics on the composition of the microbiome and metabolism in larval zebrafish. Chemosphere 2019, 217, 646–658. [Google Scholar] [CrossRef]
- Limonta, G.; Mancia, A.; Benkhalqui, A.; Bertolucci, C.; Abelli, L.; Fossi, M.C.; Panti, C. Microplastics induce transcriptional changes, immune response and behavioral alterations in adult zebrafish. Sci. Rep. 2019, 9, 15775. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Gundlach, M.; Yang, S.; Jiang, J.; Velki, M.; Yin, D.; Hollert, H. Quantitative investigation of the mechanisms of microplastics and nanoplastics toward zebrafish larvae locomotor activity. Sci. Total Environ. 2017, 584, 1022–1031. [Google Scholar] [CrossRef] [PubMed]
- Qiao, R.; Sheng, C.; Lu, Y.; Zhang, Y.; Ren, H.; Lemos, B. Microplastics induce intestinal inflammation, oxidative stress, and disorders of metabolome and microbiome in zebrafish. Sci. Total Environ. 2019, 662, 246–253. [Google Scholar] [CrossRef]
- Rochman, C.M.; Brookson, C.; Bikker, J.; Djuric, N.; Earn, A.; Bucci, K.; Athey, S.; Huntington, A.; McIlwraith, H.; Munno, K.; et al. Rethinking microplastics as a diverse contaminant suite. Environ. Toxicol. Chem. 2019, 38, 703–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercogliano, R.; Avio, C.G.; Regoli, F.; Anastasio, A.; Colavita, G.; Santonicola, S. Occurrence of microplastics in commercial seafood under the perspective of the human food chain. A review. J. Agric. Food Chem. 2020, 68, 5296–5301. [Google Scholar] [CrossRef] [PubMed]
- Belcari, P.; Sartor, P.; Sanchez, P.; Demestre, M.; Tsangridis, A.; Leondarakis, P.; Lefkaditou, E.; Papaconstantinou, C. Exploitation patterns of the cuttlefish, Sepia officinalis (Cephalopoda, Sepiidae), in the Mediterranean Sea. Bull. Mar. Sci. 2002, 71, 187–196. [Google Scholar]
- Dunn, M.R. Aspects of the stock dynamics and exploitation of cuttlefish, Sepia officinalis (Linnaeus, 1758), in the English Channel. Fish. Res. 1999, 40, 277–293. [Google Scholar] [CrossRef]
- Melli, V.; Riginella, E.; Nalon, M.; Mazzoldi, C. From Trap to Nursery. Mitigating the Impact of an Artisanal Fishery on Cuttlefish Offspring. PLoS ONE 2014, 9, e90542. [Google Scholar] [CrossRef] [Green Version]
- Russo, E.; Monti, M.A.; Mangano, M.C.; Raffaetà, A.; Sarà, G.; Silvestri, C.; Pranovi, F. Temporal and spatial patterns of trawl fishing activities in the Adriatic Sea (Central Mediterranean Sea, GSA17). Ocean Coast. Manag. 2020, 192, 105231. [Google Scholar] [CrossRef]
- Rosas-Luis, R. Description of plastic remains found in the stomach contents of the jumbo squid Dosidicus gigas landed in Ecuador during 2014. Mar. Pollut. Bull. 2016, 113, 302–305. [Google Scholar] [CrossRef]
- Ferreira, G.V.B.; Justino, A.K.S.; Eduardo, L.N.; Lenoble, V.; Fauvelle, V.; Schmidt, N.; Junior, T.V.; Frédou, T.; Lucena-Frédou, F. Plastic in the inferno: Microplastic contamination in deep-sea cephalopods (Vampyroteuthis infernalis and Abralia veranyi) from the southwestern Atlantic. Mar. Pollut. Bull. 2022, 174, 113309. [Google Scholar] [CrossRef] [PubMed]
- Ragusa, A.; Notarstefano, V.; Svelato, A.; Belloni, A.; Gioacchini, G.; Blondeel, C.; Zucchelli, E.; De Luca, C.; Avino, S.D.; Gulotta, A.; et al. Raman Microspectroscopy Detection and Characterisation of Microplastics in Human Breastmilk. Polymers 2022, 14, 2700. [Google Scholar] [CrossRef]
- Boletzky, S.V.; Andouche, A.; Bonnaud-Ponticelli, L. A developmental table of embryogenesis in sepia officinalis. Vie Milieu-Life Environ. 2016, 66, 11–23. [Google Scholar]
- Chemello, G.; Zarantoniello, M.; Randazzo, B.; Gioacchini, G.; Truzzi, C.; Cardinaletti, G.; Riolo, P.; Olivotto, I. Effects of black soldier fly (Hermetia illucens) enriched with Schizochytrium sp. on zebrafish (Danio rerio) reproductive performances. Aquaculture 2022, 550, 737853. [Google Scholar] [CrossRef]
- Di Renzo, L.; Mascilongo, G.; Berti, M.; Bogdanović, T.; Listeš, E.; Brkljača, M.; Notarstefano, V.; Gioacchini, G.; Giorgini, E.; Olivieri, V.; et al. Potential Impact of Microplastics and Additives on the Health Status of Loggerhead Turtles (Caretta caretta) Stranded Along the Central Adriatic Coast. Water Air Soil Pollut. 2021, 232, 1–20. [Google Scholar] [CrossRef]
- Dong, M.; Zhang, Q.; Xing, X.; Chen, W.; She, Z.; Luo, Z. Raman spectra and surface changes of microplastics weathered under natural environments. Sci. Total Environ. 2020, 739, 139990. [Google Scholar] [CrossRef]
- Microplastic Spectral Libraries—Rochman Lab. Available online: https://rochmanlab.wordpress.com/spectral-libraries-for-microplastics-research/ (accessed on 11 July 2022).
- Rocha, F.; Guerra, A.; González, A.F. A review of reproductive strategies in cephalopods. Biol. Rev. Camb. Philos. Soc. 2001, 76, 291–304. [Google Scholar] [CrossRef]
- Boyle, P.R. Cephalopod biology in the fisheries context. Fish. Res. 1990, 8, 303–321. [Google Scholar] [CrossRef]
- Bloor, I.S.M.; Attrill, M.J.; Jackson, E.L. A Review of the Factors Influencing Spawning, Early Life Stage Survival and Recruitment Variability in the Common Cuttlefish (Sepia officinalis). Adv. Mar. Biol. 2013, 65, 1–65. [Google Scholar]
- Boletzky, S.V. Encapsulation of Cephalopod Embryos—A Search for Functional Correlations. Am. Malacol. Bull. 1986, 4, 217–227. [Google Scholar]
- Zatylny-Gaudin, C.; Henry, J. Egg-Laying in the Cuttlefish Sepia officinalis. In Biological Resources of Water; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Chen, C.; Li, H.W.; Ku, W.L.; Lin, C.J.; Chang, C.F.; Wu, G.C. Two distinct vitellogenin genes are similar in function and expression in the bigfin reef squid Sepioteuthis lessoniana. Biol. Reprod. 2018, 99, 1034–1044. [Google Scholar] [CrossRef] [PubMed]
- Gaudron, S.M.; Zanuttini, B.; Henry, J. Yolk protein in the cephalopod, sepia officinalis: A strategy for structural characterisation. Invertebr. Reprod. Dev. 2005, 48, 129–135. [Google Scholar] [CrossRef]
- Costa, P.M.; Rodrigo, A.P.; Costa, M.H. Microstructural and histochemical advances on the digestive gland of the common cuttlefish, Sepia officinalis L. Zoomorphology 2014, 133, 59–69. [Google Scholar] [CrossRef]
- Pitt, J.A.; Trevisan, R.; Massarsky, A.; Kozal, J.S.; Levin, E.D.; Di Giulio, R.T. Maternal transfer of nanoplastics to offspring in zebrafish (Danio rerio): A case study with nanopolystyrene. Sci. Total Environ. 2018, 643, 324–334. [Google Scholar] [CrossRef]
- Veneman, W.J.; Spaink, H.P.; Brun, N.R.; Bosker, T.; Vijver, M.G. Pathway analysis of systemic transcriptome responses to injected polystyrene particles in zebrafish larvae. Aquat. Toxicol. 2017, 190, 112–120. [Google Scholar] [CrossRef]
- Ragusa, A.; Svelato, A.; Santacroce, C.; Catalano, P.; Notarstefano, V.; Carnevali, O.; Papa, F.; Rongioletti, M.C.A.; Baiocco, F.; Draghi, S.; et al. Plasticenta: First evidence of microplastics in human placenta. Environ. Int. 2021, 146, 106274. [Google Scholar] [CrossRef]
- Digka, N.; Tsangaris, C.; Kaberi, H.; Adamopoulou, A.; Zeri, C. Microplastic Abundance and Polymer Types in a Mediterranean Environment. In Proceedings of the International Conference on Microplastic Pollution in the Mediterranean Sea; Springer International Publishing: Cham, Switzerland, 2018; pp. 17–24. [Google Scholar]
- Suaria, G.; Avio, C.G.; Mineo, A.; Lattin, G.L.; Magaldi, M.G.; Belmonte, G.; Moore, C.J.; Regoli, F.; Aliani, S. The Mediterranean Plastic Soup: Synthetic polymers in Mediterranean surface waters. Sci. Rep. 2016, 6, 37551. [Google Scholar] [CrossRef] [Green Version]
- Cincinelli, A.; Martellini, T.; Guerranti, C.; Scopetani, C.; Chelazzi, D.; Giarrizzo, T. A potpourri of microplastics in the sea surface and water column of the Mediterranean Sea. TrAC—Trends Anal. Chem. 2019, 110, 321–326. [Google Scholar] [CrossRef]
- Erni-Cassola, G.; Zadjelovic, V.; Gibson, M.I.; Christie-Oleza, J.A. Distribution of plastic polymer types in the marine environment; A meta-analysis. J. Hazard. Mater. 2019, 369, 691–698. [Google Scholar] [CrossRef]
- Lim, K.P.; Lim, P.E.; Yusoff, S.; Sun, C.; Ding, J.; Loh, K.H. A Meta-Analysis of the Characterisations of Plastic Ingested by Fish Globally. Toxics 2022, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Sykes, A.V.; Almansa, E.; Lorenzo, A.; Andrade, J.P. Lipid characterization of both wild and cultured eggs of cuttlefish (Sepia officinalis L.) throughout the embryonic development. Aquac. Nutr. 2009, 15, 38–53. [Google Scholar] [CrossRef]
- Boletzky, S.V. Recent Studies on Spawning, Embryonic Development, and Hatching in the Cephalopoda. Adv. Mar. Biol. 1989, 25, 85–115. [Google Scholar]
- Boletzky, S.V.; Erlwein, B.; Hofmann, D.K. The Sepia Egg: A showcase of cephalopod embryology. Vie Milieu 2006, 56, 191–201. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | #Mps/Yolk | MPs/Yolk (MPs/g) | #MPs/Embryo | MPs/Embryo (MPs/g) |
|---|---|---|---|---|
| A | 0.45 ± 0.07 a | 14.17 ± 2.33 a | 0.2 ± 0.03 a | 2.35 ± 0.18 a |
| B | 0.15 ± 0.07 b | 4.99 ± 2.13 b | 0.00 ± 0.00 b | 0.00 ± 0.00 b |
| C | 0.40 ± 0.01 a | 18.64 ± 0.18 a | 0.25 ± 0.07 a | 2.35 ± 0.67 a |
| D | 0.25 ± 0.07 ab | 10.59 ± 0.78 c | 0.05 ± 0.07 b | 0.62 ± 0.88 ab |
| Site | Polymer (Yolk) | Polymer (Embryo) |
|---|---|---|
| A | PES, PVC, PA, PP | Ac, PP, PS |
| B | PP | ND |
| C | CA, PVC | PVC |
| D | PVC | PP |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chemello, G.; Faraoni, V.; Notarstefano, V.; Maradonna, F.; Carnevali, O.; Gioacchini, G. First Evidence of Microplastics in the Yolk and Embryos of Common Cuttlefish (Sepia officinalis) from the Central Adriatic Sea: Evaluation of Embryo and Hatchling Structural Integrity and Development. Animals 2023, 13, 95. https://doi.org/10.3390/ani13010095
Chemello G, Faraoni V, Notarstefano V, Maradonna F, Carnevali O, Gioacchini G. First Evidence of Microplastics in the Yolk and Embryos of Common Cuttlefish (Sepia officinalis) from the Central Adriatic Sea: Evaluation of Embryo and Hatchling Structural Integrity and Development. Animals. 2023; 13(1):95. https://doi.org/10.3390/ani13010095
Chicago/Turabian StyleChemello, Giulia, Viola Faraoni, Valentina Notarstefano, Francesca Maradonna, Oliana Carnevali, and Giorgia Gioacchini. 2023. "First Evidence of Microplastics in the Yolk and Embryos of Common Cuttlefish (Sepia officinalis) from the Central Adriatic Sea: Evaluation of Embryo and Hatchling Structural Integrity and Development" Animals 13, no. 1: 95. https://doi.org/10.3390/ani13010095