
Dietary Use of Methionine Sources and Bacillus amyloliquefaciens CECT 5940 Influences Growth Performance, Hepatopancreatic Histology, Digestion, Immunity, and Digestive Microbiota of Litopenaeus vannamei Fed Reduced Fishmeal Diets

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diet Preparation
2.2. Shrimp, Feeding Trial and Sample Collection
2.3. Growth Performance
- Final weight = (Σ Final individual weight)/Final number of shrimps.
- Weekly weight gain = (Final weight − Initial weight)/Number of weeks.
- Specific growth rate = 100 × (ln final weight − ln initial weight)/days of experiment.
- Survival rate = 100 × (Final number of shrimps/Initial number of shrimps).
- Final biomass = Final weight × Final number of shrimps.
- Feed intake = Feed Input (dry weight) − Feed collected (dry weight).
- Feed conversion rate = Feed intake/Final biomass.
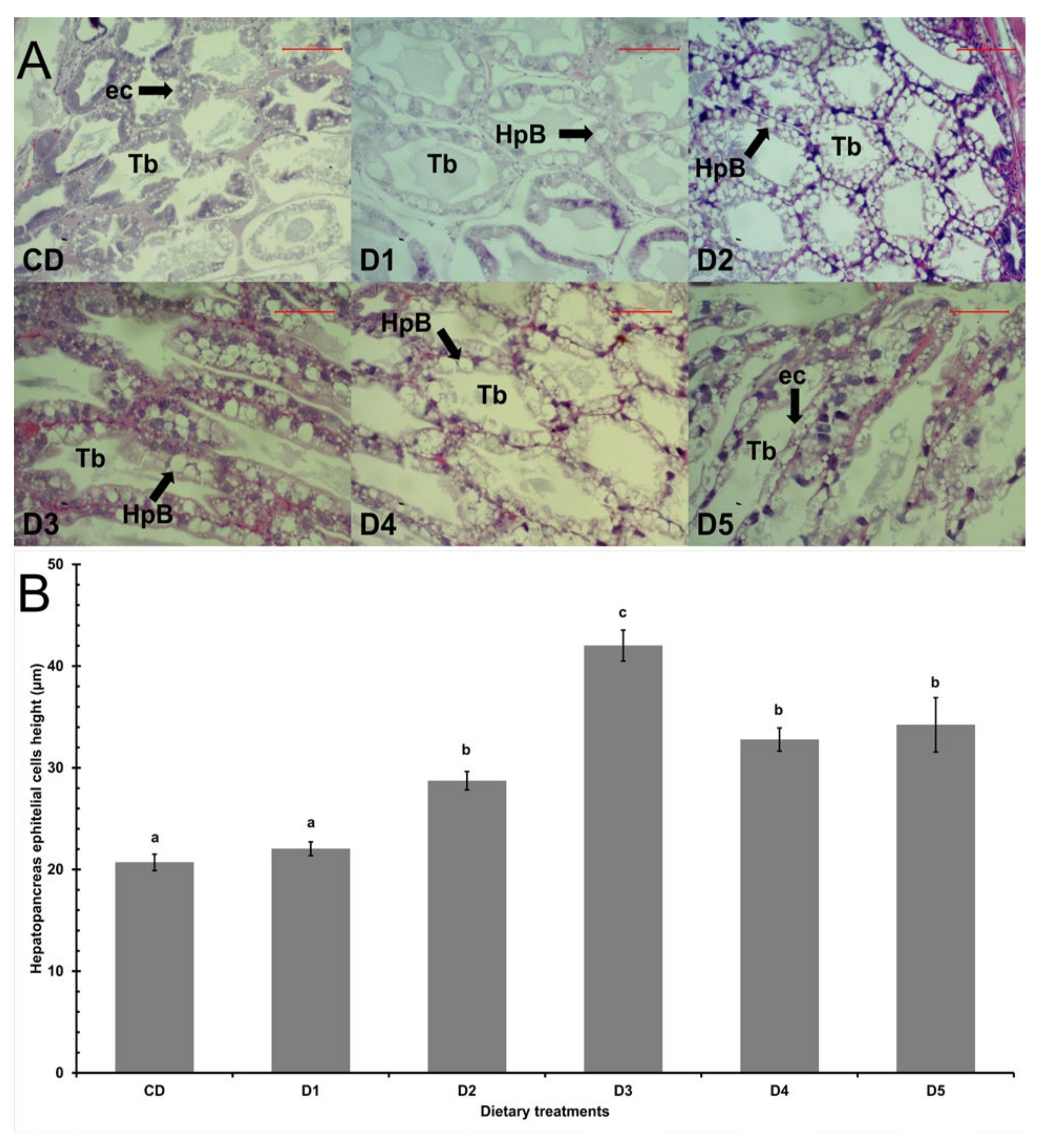
2.4. Hepatopancreas Histology
2.5. Digestive Enzymes’ Activity
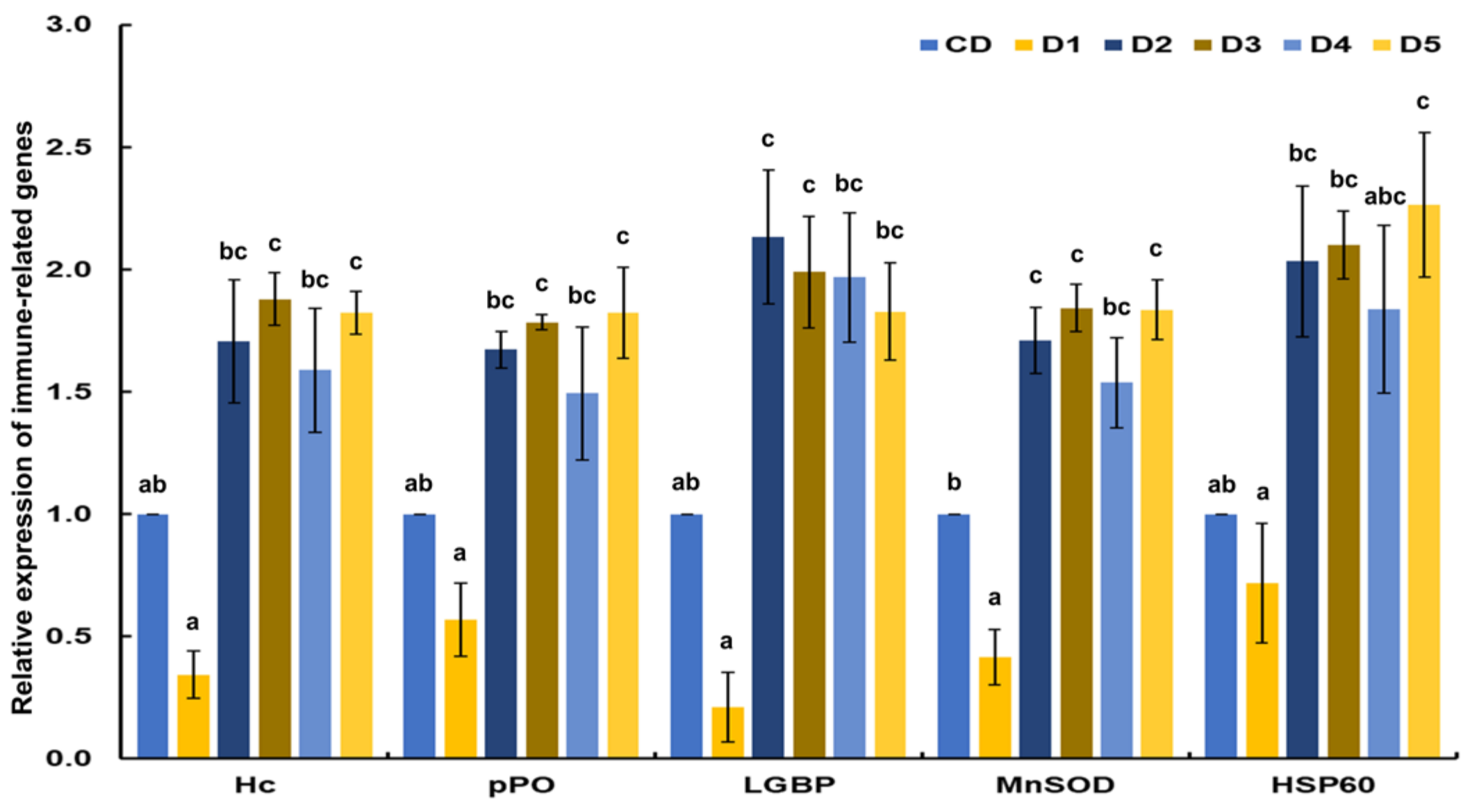
2.6. Transcriptional Response of Immune-Related Genes
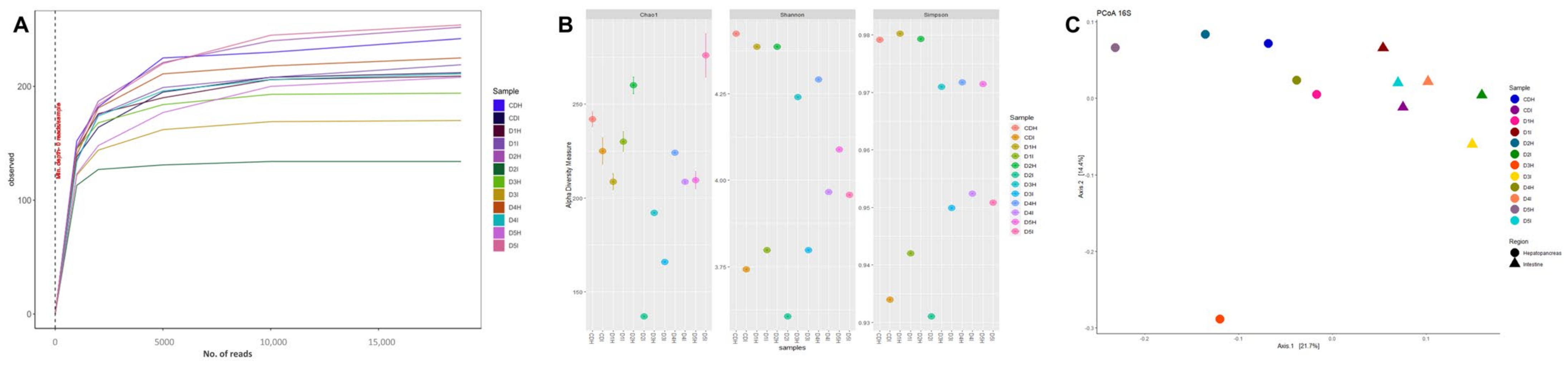
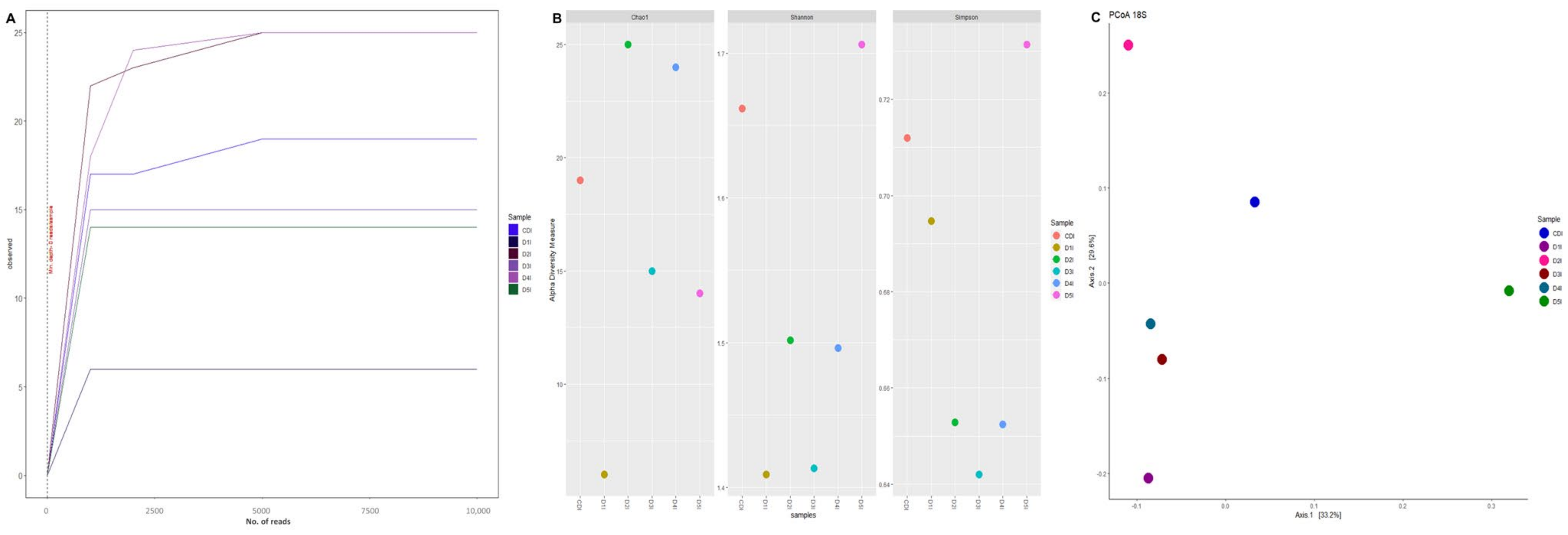
2.7. DNA Extraction and Sequencing Analysis
2.8. Bioinformatic Analysis
2.9. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Hepatopancreatic Histology
3.3. Digestive Enzyme Activity
3.4. Transcriptional Response of Immune-Related Genes
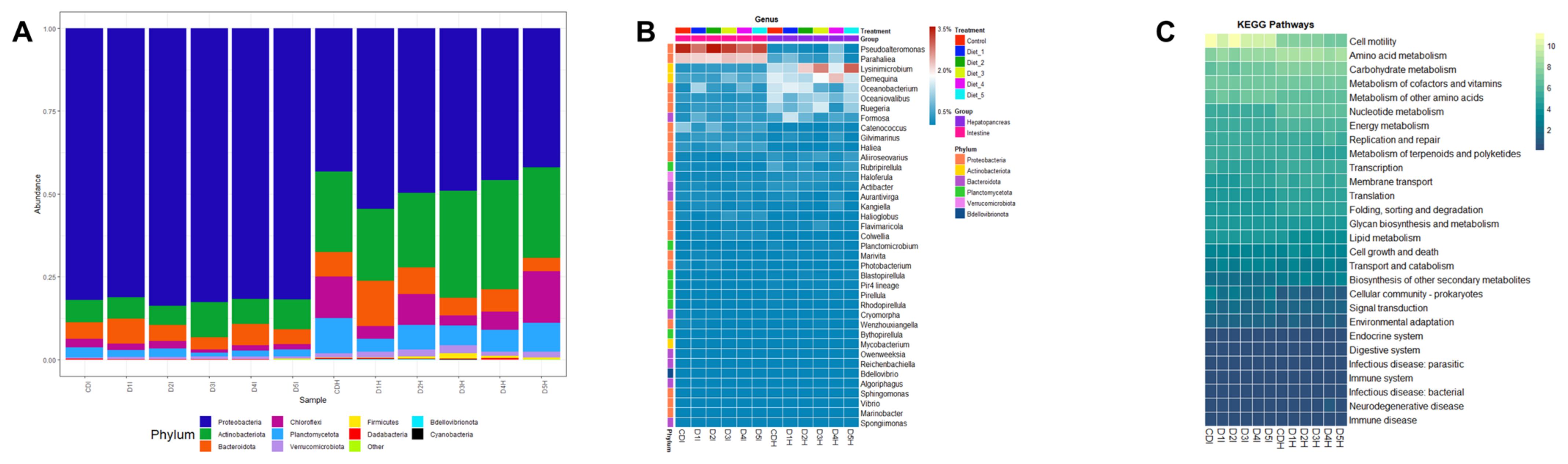
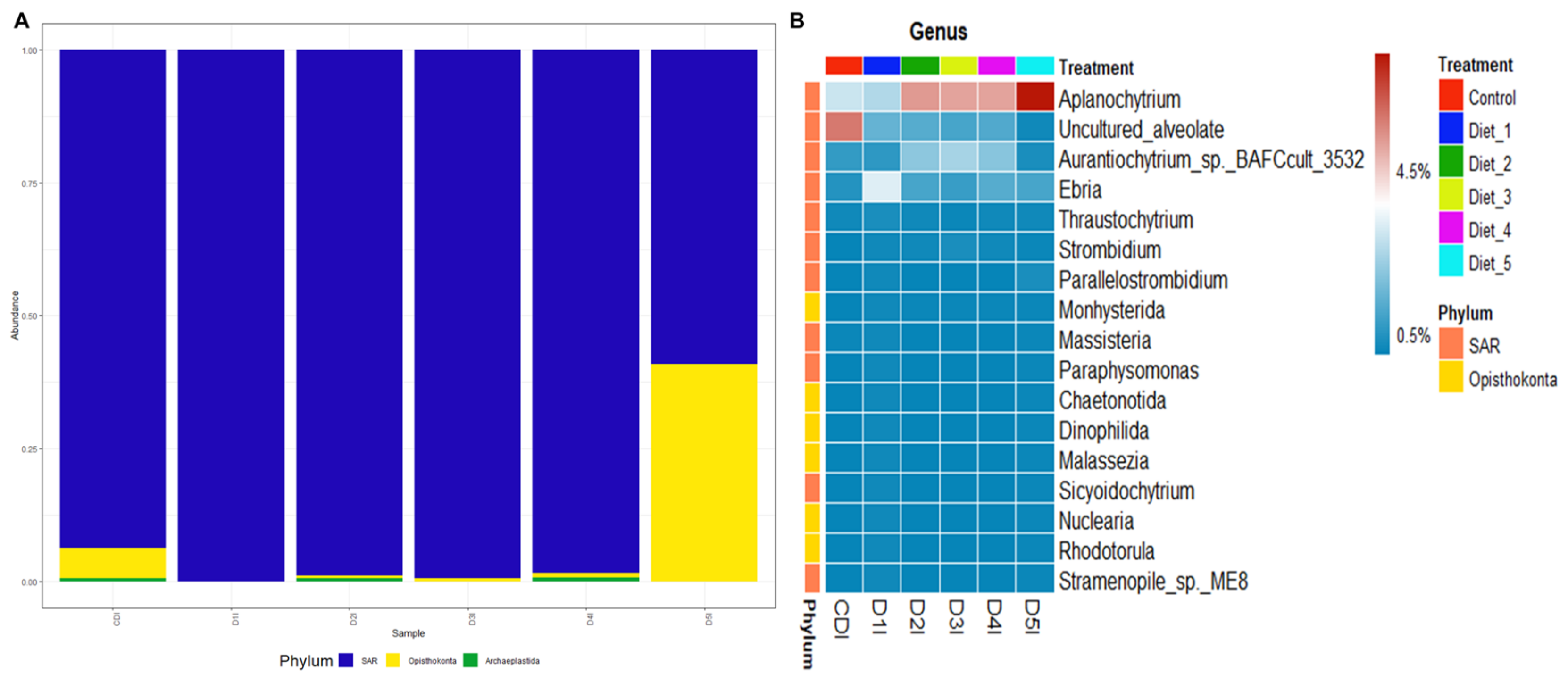
3.5. Digestive Microbiota Analysis and Functional Prediction
4. Discussion
4.1. Growth Performance
4.2. Hepatopancreatic Histology
4.3. Digestive Enzyme Analysis
4.4. Transcriptional Response of Immune-Related Genes
4.5. Digestive Microbiota Analysis and Functional Prediction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy. [CrossRef]
- Xie, J.-J.; Lemme, A.; He, J.-Y.; Yin, P.; Figueiredo-Silva, C.; Liu, Y.-J.; Xie, S.-W.; Niu, J.; Tian, L.-X. Fishmeal levels can be successfully reduced in white shrimp (Litopenaeus vannamei) if supplemented with DL-Methionine (DL-Met) or DL-Methionyl-DL-Methionine (Met-Met). Aquac. Nutr. 2017, 24, 1144–1152. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, X.; Zhou, Q.; Cheng, Y.; Luo, J.; Masagounder, K.; He, S.; Zhu, T.; Yuan, Y.; Shi, B.; et al. Dietary DL-methionyl-DL-methionine supplementation could improve growth performance under low fishmeal strategies by modulating TOR signalling pathway of Litopenaeus vannamei. Aquac. Nutr. 2021, 27, 1921–1933. [Google Scholar] [CrossRef]
- Cho, J.H.; Kim, I.H. Fish meal-nutritive value. J. Anim. Physiol. Anim. Nutr. 2011, 95, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Ye, H.; Jin, F.; Li, H.; Kim, Y.-S.; Xiao, J.; Guo, Z. Effects of fermented soybean meal and guanosine 5′-monophosphate on growth, intestinal health and anti-stress capability of Penaeus vannamei in low fish meal diet. Aquaculture 2022, 548, 737591. [Google Scholar] [CrossRef]
- Yao, W.; Yang, P.; Zhang, X.; Xu, X.; Zhang, C.; Li, X.; Leng, X. Effects of replacing dietary fishmeal with Clostridium autoethanogenum protein on growth and flesh quality of Pacific white shrimp (Litopenaeus vannamei). Aquaculture 2022, 549, 737770. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, H.; Yan, Y.; Zhang, C.; Ye, J.; Sun, Y. Effects of fish origin probiotics on growth performance, immune response and intestinal health of shrimp (Litopenaeus vannamei) fed diets with fish meal partially replaced by soybean meal. Aquac. Nutr. 2020, 26, 1255–1265. [Google Scholar] [CrossRef]
- Ji, R.; Wang, Z.; He, J.; Masagounder, K.; Xu, W.; Mai, K.; Ai, Q. Effects of DL-methionyl-DL-methionine supplementation on growth performance, immune and antioxidative responses of white leg shrimp (Litopenaeus vannamei) fed low fishmeal diet. Aquac. Rep. 2021, 21, 100785. [Google Scholar] [CrossRef]
- Chaklader, R.; Siddik, M.A.B.; Fotedar, R. Total replacement of fishmeal with poultry by-product meal affected the growth, muscle quality, histological structure, antioxidant capacity and immune response of juvenile barramundi, Lates calcarifer. PLoS ONE 2020, 15, e0242079. [Google Scholar] [CrossRef]
- Niu, J.; Lemme, A.; He, J.-Y.; Li, H.-Y.; Xie, S.-W.; Liu, Y.-J.; Yang, H.-J.; Figueiredo-Silva, C.; Tian, L.-X. Assessing the bioavailability of the Novel Met-Met product (AQUAVI® Met-Met) compared to dl-methionine (dl-Met) in white shrimp (Litopenaeus vannamei). Aquaculture 2018, 484, 322–332. [Google Scholar] [CrossRef]
- Skiba-Cassy, S.; Geurden, I.; Panserat, S.; Seiliez, I. Dietary methionine imbalance alters the transcriptional regulation of genes involved in glucose, lipid and amino acid metabolism in the liver of rainbow trout (Oncorhynchus mykiss). Aquaculture 2016, 454, 56–65. [Google Scholar] [CrossRef]
- Machado, M.; Serra, C.R.; Oliva-Teles, A.; Costas, B. Methionine and Tryptophan Play Different Modulatory Roles in the European Seabass (Dicentrarchus labrax) Innate Immune Response and Apoptosis Signaling—An In Vitro Study. Front. Immunol. 2021, 12, 660448. [Google Scholar] [CrossRef]
- Zhou, Y.; He, J.; Su, N.; Masagounder, K.; Xu, M.; Chen, L.; Liu, Q.; Ye, H.; Sun, Z.; Ye, C. Effects of DL-methionine and a methionine hydroxy analogue (MHA-Ca) on growth, amino acid profiles and the expression of genes related to taurine and protein synthesis in common carp (Cyprinus carpio). Aquaculture 2021, 532, 735962. [Google Scholar] [CrossRef]
- Guo, T.-Y.; Zhao, W.; He, J.-Y.; Liao, S.-Y.; Xie, J.-J.; Xie, S.-W.; Masagounder, K.; Liu, Y.-J.; Tian, L.-X.; Niu, J. Dietary dl-methionyl-dl-methionine supplementation increased growth performance, antioxidant ability, the content of essential amino acids and improved the diversity of intestinal microbiota in Nile tilapia (Oreochromis niloticus). Br. J. Nutr. 2020, 123, 72–83. [Google Scholar] [CrossRef]
- Lin, J.; Zeng, Q.; Zhang, C.; Song, K.; Lu, K.; Wang, L.; Rahimnejad, S. Effects of Bacillus subtilis supplementation in soybean meal-based diets on growth performance, diet digestibility and gut health in bullfrog Lithobates catesbeianus. Aquac. Rep. 2020, 16, 100273. [Google Scholar] [CrossRef]
- Tao, X.; He, J.; Lu, J.; Chen, Z.; Jin, M.; Jiao, L.; Masagounder, K.; Liu, W.; Zhou, Q. Effects of Bacillus subtilis DSM 32315 (Gutcare®) on the growth performance, antioxidant status, immune ability and intestinal function for juvenile Litopenaeus vannamei fed with high/low-fishmeal diets. Aquac. Rep. 2022, 26, 101282. [Google Scholar] [CrossRef]
- De Oliveira, M.J.K.; Sakomura, N.K.; Dorigam, J.C.D.P.; Doranalli, K.; Soares, L.; Viana, G. Bacillus amyloliquefaciens CECT 5940 alone or in combination with antibiotic growth promoters improves performance in broilers under enteric pathogen challenge. Poult. Sci. 2019, 98, 4391–4400. [Google Scholar] [CrossRef]
- Silva, T.F.A.; Petrillo, T.R.; Yunis-Aguinaga, J.; Marcusso, P.; Claudiano, G.D.S.; de Moraes, F.R.; de Moraes, J.R.E. Effects of the probiotic Bacillus amyloliquefaciens on growth performance, hematology and intestinal morphometry in cage-reared Nile tilapia. Lat. Am. J. Aquat. Res. 2015, 43, 963–971. [Google Scholar] [CrossRef]
- Bai, S.; Katya, K.; Yun, H. Additives in aquafeed. In Feed and Feeding Practices in Aquaculture; Woodhead Publishing: Sawston, UK, 2015; pp. 171–202. [Google Scholar] [CrossRef]
- Wang, L.; Ye, L.; Hua, Y.; Zhang, G.; Li, Y.; Zhang, J.; He, J.; Liu, M.; Shao, Q. Effects of dietarydl-methionyl-dl-methionine (Met-Met) on growth performance, body composition and haematological parameters of white shrimp (Litopenaeus vannamei) fed with plant protein–based diets. Aquac. Res. 2019, 50, 1718–1730. [Google Scholar] [CrossRef]
- Al-Deriny, S.H.; Dawood, M.A.; Zaid, A.A.A.; El-Tras, W.F.; Paray, B.A.; Van Doan, H.; Mohamed, R.A. The synergistic effects of Spirulina platensis and Bacillus amyloliquefaciens on the growth performance, intestinal histomorphology, and immune response of Nile tilapia (Oreochromis niloticus). Aquac. Rep. 2020, 17, 100390. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, Y.; Liu, M.; Li, J.; Lai, W.; Geng, S.; Yuan, T.; Liu, Y.; Di, Y.; Zhang, W.; et al. Effects of dietary Bacillus amyloliquefaciens CECT 5940 supplementation on growth performance, antioxidant status, immunity, and digestive enzyme activity of broilers fed corn-wheat-soybean meal diets. Poult. Sci. 2022, 101, 101585. [Google Scholar] [CrossRef]
- NRC (National Research Council). Nutrient Requirements of Fish and Shrimp; Natlional Academic Press: Washington, DC, USA, 2011. [Google Scholar] [CrossRef]
- Guo, J.; Davis, R.; Starkey, C.; Davis, D.A. Efficacy of various coated materials to prevent nutrient leaching for Pacific white shrimp Litopenaeus vannamei commercial diets. J. World Aquac. Soc. 2020, 52, 195–203. [Google Scholar] [CrossRef]
- International Office of Epizootics; Aquatic Animal Health Standards Commission. Manual of diagnostic tests for aquatic animals. Office international des épizooties. 2022. Available online: https://www.woah.org/en/what-we-do/standards/codes-and-manuals/aquatic-manual-online-access/ (accessed on 1 September 2022).
- Martinez-Porchas, M.; Ezquerra-Brauer, M.; Mendoza-Cano, F.; Higuera, J.E.C.; Vargas-Albores, F.; Martinez-Cordova, L.R. Effect of supplementing heterotrophic and photoautotrophic biofloc, on the production response, physiological condition and post-harvest quality of the whiteleg shrimp, Litopenaeus vannamei. Aquac. Rep. 2020, 16, 100257. [Google Scholar] [CrossRef]
- Peña-Rodríguez, A.; Elizondo-González, R.; Nieto-López, M.G.; Ricque-Marie, D.; Cruz-Suárez, L.E. Practical diets for the sustainable production of brown shrimp, Farfantepenaeus californiensis, juveniles in presence of the green macroalga Ulva clathrata as natural food. J. Appl. Phycol. 2017, 29, 413–421. [Google Scholar] [CrossRef]
- Piérri, V.; Valter-Severino, D.; Goulart-De-Oliveira, K.; Manoel-Do-Espírito-Santo, C.; Nascimento-Vieira, F.; Quadros-Seiffert, W. Cultivation of marine shrimp in biofloc technology (BFT) system under different water alkalinities. Braz. J. Biol. 2015, 75, 558–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Galaviz, J.R.; Casillas-Hernández, R.; Flores-Perez, M.B.; Lares-Villa, F.; Bórquez-López, R.A.; Gil-Núñez, J.C. Effect of genotype and protein source on performance of Pacific white shrimp (Litopenaeus vannamei). Ital. J. Anim. Sci. 2020, 19, 289–294. [Google Scholar] [CrossRef] [Green Version]
- Bell, T.A.; Lightner, D.V. A Handbook of Normal Penaeid Shrimp Histology; Food and Agriculture Organization of the United Nations: Rome, Italy, 1988. [Google Scholar]
- Casillas-Hernández, R.; Arévalo-Sainz, K.J.; Gonzalez-Galaviz, J.R.; Rodríguez-Jaramillo, M.D.C.; Bórquez-López, R.A.; Gil-Núñez, J.C.; Flores-Pérez, M.B.; Lares-Villa, F.; Ibarra-Gámez, J.C.; Barrios, R.M.M. The effects of high concentration of carbon dioxide on performance and tissue histology of shrimp Litopenaeus vannamei. Aquac. Res. 2021, 52, 3991–3996. [Google Scholar] [CrossRef]
- Xiao, Z.; Storms, R.; Tsang, A. A quantitative starch–iodine method for measuring alpha-amylase and glucoamylase activities. Anal. Biochem. 2006, 351, 146–148. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Zhang, S.-P.; Li, J.-F.; Wu, X.-C.; Zhong, W.-J.; Xian, J.-A.; Liao, S.-A.; Miao, Y.-T.; Wang, A.-L. Effects of different dietary lipid level on the growth, survival and immune-relating genes expression in Pacific white shrimp, Litopenaeus vannamei. Fish Shellfish. Immunol. 2013, 34, 1131–1138. [Google Scholar] [CrossRef]
- Anaya, L.R.; Casillas-Hernández, R.; Flores-Pérez, M.B.; Lares-Villa, F.; Lares-Jiménez, L.F.; Luna-Nevarez, P.; Gonzalez-Galaviz, J.R. Effect of genetic line, protein source, and protein level on growth, survival, and immune-related gene expression of Litopenaeus vannamei. J. World Aquac. Soc. 2020, 51, 1161–1174. [Google Scholar] [CrossRef]
- Casillas-Hernández, R.; Arévalo-Sainz, K.J.; Flores-Pérez, M.B.; Garcia-Clark, J.G.; Rodriguez-Anaya, L.Z.; Lares-Villa, F.; Bórquez-López, R.A.; Gil-Núñez, J.C.; Gonzalez-Galaviz, J.R. Transcriptional response of immune-related genes in Litopenaeus vannamei cultured in recirculating aquaculture systems with elevated CO2. Rev. Bras. de Zootec. 2021, 50, 20200197. [Google Scholar] [CrossRef]
- Guzmán-Villanueva, L.T.; Escobedo-Fregoso, C.; Barajas-Sandoval, D.R.; Gomez-Gil, B.; Peña-Rodríguez, A.; Martínez-Diaz, S.F.; Balcazar, J.L.; Quiroz-Guzmán, E. Assessment of microbial dynamics and antioxidant enzyme gene expression following probiotic administration in farmed Pacific white shrimp (Litopenaeus vannamei). Aquaculture 2020, 519, 734907. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Lahti, L.; Shetty, S. Microbiome R package; Release (3.16); Bioconductor: Boston, MA, USA, 2017. [Google Scholar] [CrossRef]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Cornejo-Granados, F.; Lopez-Zavala, A.A.; Gallardo-Becerra, L.; Mendoza-Vargas, A.; Sánchez, F.; Vichido, R.; Brieba, L.G.; Viana, M.T.; Sotelo-Mundo, R.R.; Ochoa-Leyva, A. Microbiome of Pacific Whiteleg shrimp reveals differential bacterial community composition between Wild, Aquacultured and AHPND/EMS outbreak conditions. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Façanha, F.N.; Oliveira-Neto, A.R.; Figueiredo-Silva, C.; Nunes, A.J. Effect of shrimp stocking density and graded levels of dietary methionine over the growth performance of Litopenaeus vannamei reared in a green-water system. Aquaculture 2016, 463, 16–21. [Google Scholar] [CrossRef]
- Nunes, A.J.P.; Sabry-Neto, H.; Masagounder, K. Crude protein in low-fish meal diets for juvenile Litopenaeus vannamei can be reduced through a well-balanced supplementation of essential amino acids. J. World Aquac. Soc. 2019, 50, 1093–1107. [Google Scholar] [CrossRef]
- Llario, F.; Falco, S.; Sebastiá-Frasquet, M.-T.; Escrivá, J.; Rodilla, M.; Poersch, L.H. The Role of Bacillus amyloliquefaciens on Litopenaeus vannamei During the Maturation of a Biofloc System. J. Mar. Sci. Eng. 2019, 7, 228. [Google Scholar] [CrossRef] [Green Version]
- Llario, F.; Romano, L.A.; Rodilla, M.; Sebastiá-Frasquet, M.T.; Poersch, L.H. Application of Bacillus amyloliquefaciens as probiotic for Litopenaeus vannamei (Boone, 1931) cultivated in a biofloc system. Iran. J. Fish. Sci. 2020, 19, 904–920. [Google Scholar]
- Gharib-Naseri, K.; Dorigam, J.C.D.P.; Doranalli, K.; Kheravii, S.; Swick, R.A.; Choct, M.; Wu, S.-B. Modulations of genes related to gut integrity, apoptosis, and immunity underlie the beneficial effects of Bacillus amyloliquefaciens CECT 5940 in broilers fed diets with different protein levels in a necrotic enteritis challenge model. J. Anim. Sci. Biotechnol. 2020, 11, 104. [Google Scholar] [CrossRef]
- Ajogwu, T.M.C.; Ekwealor, C.C.; Mbah, A.N.; Madukwe, E.J.; Ekwealor, I.A. Screening of Methionine-producing Bacillus Species from Nigerian Fermented Food Condiments and Effects of Some Cultural Parameters on Methionine Accumulation. Am. J. Food Nutr. 2020, 9, 16–22. [Google Scholar] [CrossRef]
- Yu, M.-C.; Li, Z.-J.; Lin, H.-Z.; Wen, G.-L.; Ma, S. Effects of dietary Bacillus and medicinal herbs on the growth, digestive enzyme activity, and serum biochemical parameters of the shrimp Litopenaeus vannamei. Aquac. Int. 2008, 16, 471–480. [Google Scholar] [CrossRef]
- Yılmaz, S.; Ergun, S.; Yigit, M.; Çelik, E. Effect of combination of dietary Bacillus subtilis and trans -cinnamic acid on innate immune responses and resistance of rainbow trout, Oncorhynchus mykiss to Yersinia ruckeri. Aquac. Res. 2020, 51, 441–454. [Google Scholar] [CrossRef]
- Zare, R.; Kenari, A.A.; Sadati, M.Y. Influence of dietary acetic acid, protexin (probiotic), and their combination on growth performance, intestinal microbiota, digestive enzymes, immunological parameters, and fatty acids composition in Siberian sturgeon (Acipenser baerii, Brandt, 1869). Aquac. Int. 2021, 29, 891–910. [Google Scholar] [CrossRef]
- Rodjan, P.; Soisuwan, K.; Thongprajukaew, K.; Theapparat, Y.; Khongthong, S.; Jeenkeawpieam, J.; Salaeharae, T. Effect of organic acids or probiotics alone or in combination on growth performance, nutrient digestibility, enzyme activities, intestinal morphology and gut microflora in broiler chickens. J. Anim. Physiol. Anim. Nutr. 2018, 102, e931–e940. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Chu, J.; Kirby, R.; Sheen, S.; Chien, A. The effects of replacing fish meal protein with a mixture of poultry by-product meal and fermented soybean meal on the growth performance and tissue nutritional composition of Asian seabass (Lates calcarifer). Aquac. Res. 2021, 52, 4105–4115. [Google Scholar] [CrossRef]
- Burri, L.; Berge, K.; Chumkam, S.; Jintasataporn, O. Effects of different feed inclusion levels of krill meal on growth and hepatopancreas morphology of Litopenaeus vannamei. Aquac. Res. 2020, 51, 4819–4823. [Google Scholar] [CrossRef]
- Murashita, K.; Matsunari, H.; Fukada, H.; Suzuki, N.; Furuita, H.; Oku, H.; Rønnestad, I.; Yoshinaga, H.; Yamamoto, T. Effect of a plant-based low-fishmeal diet on digestive physiology in yellowtail Seriola quinqueradiata. Aquaculture 2019, 506, 168–180. [Google Scholar] [CrossRef]
- Rahimnejad, S.; Yuan, X.; Wang, L.; Lu, K.; Song, K.; Zhang, C. Chitooligosaccharide supplementation in low-fish meal diets for Pacific white shrimp (Litopenaeus vannamei): Effects on growth, innate immunity, gut histology, and immune-related genes expression. Fish Shellfish. Immunol. 2018, 80, 405–415. [Google Scholar] [CrossRef]
- Peña-Rodríguez, A.; Pelletier-Morreeuw, Z.; García-Luján, J.; Rodríguez-Jaramillo, M.D.C.; Guzmán-Villanueva, L.; Escobedo-Fregoso, C.; Tovar-Ramírez, D.; Reyes, A.G. Evaluation ofAgave lechuguillaby-product crude extract as a feed additive for juvenile shrimpLitopenaeus vannamei. Aquac. Res. 2019, 51, 1336–1345. [Google Scholar] [CrossRef]
- Ray, G.W.; Liang, D.; Yang, Q.; Tan, B.; Dong, X.; Chi, S.; Liu, H.; Zhang, S.; Rimei, L. Effects of replacing fishmeal with dietary soybean protein concentrate (SPC) on growth, serum biochemical indices, and antioxidative functions for juvenile shrimp Litopenaeus vannamei. Aquaculture 2020, 516, 734630. [Google Scholar] [CrossRef]
- Ambasankar, K.; Dayal, J.S.; Vasagam, K.K.; Sivaramakrishnan, T.; Sandeep, K.P.; Panigrahi, A.; Ananda Raja, R.; Burri, L.; Vijayan, K.K. Growth, fatty acid composition, immune-related gene expression, histology and haematology indices of Penaeus vannamei fed graded levels of Antarctic krill meal at two different fishmeal concentrations. Aquaculture 2022, 553, 738069. [Google Scholar] [CrossRef]
- Noor, Z.; Noor, M.; Khan, S.A.; Younas, W.; Ualiyeva, D.; Hassan, Z.; Yousafzai, A.M. Dietary supplementations of methionine improve growth performances, innate immunity, digestive enzymes, and antioxidant activities of rohu (Labeo rohita). Fish Physiol. Biochem. 2021, 47, 451–464. [Google Scholar] [CrossRef]
- Yun, H.; Shahkar, E.; Hamidoghli, A.; Lee, S.; Won, S.; Bai, S.C. Evaluation of dietary soybean meal as fish meal replacer for juvenile whiteleg shrimp, Litopenaeus vannamei reared in biofloc system. Int. Aquat. Res. 2017, 9, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Yang, H.-L.; Yan, Y.-Y.; Zhang, C.-X.; Ye, J.-D.; Lu, K.-L.; Hu, L.-H.; Zhang, J.-J.; Ruan, L.; Sun, Y.-Z. Effects of fructooligosaccharide on growth, immunity and intestinal microbiota of shrimp (Litopenaeus vannamei ) fed diets with fish meal partially replaced by soybean meal. Aquac. Nutr. 2019, 25, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Mamauag, R.E.P.; Gao, J.; Nguyen, B.T.; Ragaza, J.A.; Koshio, S.; Ishikawa, M.; Yokoyama, S. Supplementations of dl-Methionine and Methionine Dipeptide in Diets are Effective for the Development and Growth of Larvae and Juvenile Red Sea Bream, Pagrus major. J. World Aquac. Soc. 2012, 43, 362–374. [Google Scholar] [CrossRef]
- Wu, P.; Tang, L.; Jiang, W.; Hu, K.; Liu, Y.; Jiang, J.; Kuang, S.; Tang, W.; Zhang, Y.; Zhou, X.; et al. The relationship between dietary methionine and growth, digestion, absorption, and antioxidant status in intestinal and hepatopancreatic tissues of sub-adult grass carp (Ctenopharyngodon idella). J. Anim. Sci. Biotechnol. 2017, 8, 1–14. [Google Scholar] [CrossRef]
- Encarnação, P. Functional Feed Additives in Aquaculture Feeds. In Aquafeed Formulation; Academic Press: Cambridge, MA, USA, 2016; pp. 217–237. [Google Scholar] [CrossRef]
- Xie, S.; Wei, D.; Yin, P.; Zheng, L.; Guo, T.; Liu, Y.; Tian, L.; Niu, J. Dietary replacement of fish-meal impaired protein synthesis and immune response of juvenile Pacific white shrimp, Litopenaeus vannamei at low salinity. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2018, 228, 26–33. [Google Scholar] [CrossRef]
- Ayiku, S.; Shen, J.; Tan, B.-P.; Dong, X.-H.; Liu, H.-Y. Effects of reducing dietary fishmeal with yeast supplementations on Litopenaeus vannamei growth, immune response and disease resistance against Vibrio harveyi. Microbiol. Res. 2020, 239, 126554. [Google Scholar] [CrossRef]
- Xie, S.; Wei, D.; Liu, Y.; Tian, L.; Niu, J. Dietary fish oil levels modulated lipid metabolism, immune response, intestinal health and salinity stress resistance of juvenile Penaeus monodon fed a low fish-meal diet. Anim. Feed. Sci. Technol. 2022, 289, 115321. [Google Scholar] [CrossRef]
- Xie, S.; Niu, J.; Zhou, W.; Liu, Y.; Tian, L. Developing a low fishmeal diet for juvenile Pacific white shrimp, Litopenaeus vannamei, using the nutritional value of FM as the reference profile. Aquac. Nutr. 2018, 24, 1184–1197. [Google Scholar] [CrossRef]
- Coutinho, F.; Simões, R.; Monge-Ortiz, R.; Furuya, W.; Pousão-Ferreira, P.; Kaushik, S.; Oliva-Teles, A.; Peres, H. Effects of dietary methionine and taurine supplementation to low-fish meal diets on growth performance and oxidative status of European sea bass (Dicentrarchus labrax) juveniles. Aquaculture 2017, 479, 447–454. [Google Scholar] [CrossRef]
- Li, J.; Jiang, H.; Li, L.; Zhang, X.; Chen, J. The Effect of Disease and Season to Hepatopancreas and Intestinal Mycobiota of Litopenaeus vannamei. Front. Microbiol. 2019, 10, 889. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Wang, Y.; Dong, H.; Ding, X.; Liu, Q.; Li, H.; Zhang, J.; Xiong, D. Changes in the Intestine Microbial, Digestive, and Immune-Related Genes of Litopenaeus vannamei in Response to Dietary Probiotic Clostridium butyricum Supplementation. Front. Microbiol. 2018, 9, 2191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.; Wang, Y.; Ding, X.; Xiong, D.; Zhang, J. Response of intestine microbiota, digestion, and immunity in Pacific white shrimp Litopenaeus vannamei to dietary succinate. Aquaculture 2020, 517, 734762. [Google Scholar] [CrossRef]
- Pacheco-Sandoval, A.; Schramm, Y.; Heckel, G.; Brassea-Pérez, E.; Martinez-Porchas, M.; Lago-Lestón, A. The Pacific harbor seal gut microbiota in Mexico: Its relationship with diet and functional inferences. PLoS ONE 2019, 14, e0221770. [Google Scholar] [CrossRef]
- Garibay-Valdez, E.; Cicala, F.; Martinez-Porchas, M.; Gómez-Reyes, R.; Vargas-Albores, F.; Gollas-Galván, T.; Martínez-Córdova, L.R.; Calderón, K. Longitudinal variations in the gastrointestinal microbiome of the white shrimp, Litopenaeus vannamei. PeerJ 2021, 9, e11827. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, Y.; Tan, Y.; Kumkhong, S.; Gu, Y.; Yu, H.; Yang, Y. Effects of deoxynivalenol-contaminated diet on the composition and diversity of the intestinal microbial community and intestinal ultrastructure of juvenile largemouth bass (Micropterus salmoides). Aquaculture 2021, 538, 736544. [Google Scholar] [CrossRef]
- Cicala, F.; Lago-Lestón, A.; Gomez-Gil, B.; Gollas-Galván, T.; Chong-Robles, J.; Cortés-Jacinto, E.; Martínez-Porchas, M. Gut microbiota shifts in the giant tiger shrimp, Penaeus monodon, during the postlarvae, juvenile, and adult stages. Aquac. Int. 2020, 28, 1421–1433. [Google Scholar] [CrossRef]
- Tzuc, J.T.; Escalante, D.R.; Herrera, R.R.; Cortés, G.G.; Ortiz, M.L.A. Microbiota from Litopenaeus vannamei: Digestive tract microbial community of Pacific white shrimp (Litopenaeus vannamei). SpringerPlus 2014, 3, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Li, E.; Xu, C.; Wang, X.; Wang, S.; Zhao, Q.; Zhang, M.; Qin, J.G.; Chen, L. Gut Microbiota and its Modulation for Healthy Farming of Pacific White ShrimpLitopenaeus vannamei. Rev. Fish. Sci. Aquac. 2018, 26, 381–399. [Google Scholar] [CrossRef]
- Garibay-Valdez, E.; Martínez-Porchas, M.; Calderón, K.; Vargas-Albores, F.; Gollas-Galván, T.; Martínez-Córdova, L. Taxonomic and functional changes in the microbiota of the white shrimp (Litopenaeus vannamei) associated with postlarval ontogenetic development. Aquaculture 2020, 518, 734842. [Google Scholar] [CrossRef]
- Chan, J.; Geng, D.; Pan, B.; Zhang, Q.; Xu, Q. Metagenomic Insights Into the Structure and Function of Intestinal Microbiota of the Hadal Amphipods. Front. Microbiol. 2021, 12, 668989. [Google Scholar] [CrossRef]
- Rajeev, R.; Adithya, K.; Kiran, G.S.; Selvin, J. Healthy microbiome: A key to successful and sustainable shrimp aquaculture. Rev. Aquac. 2021, 13, 238–258. [Google Scholar] [CrossRef]
- Escamilla-Montes, R.; Barraza, A.; Luna-González, A.; Angulo, C.; Fierro-Coronado, J.A.; Diarte-Plata, G.; Flores-Miranda, M.D.C. Effect of dietary inulin in the gut microbiota of whiteleg shrimp Penaeus vannamei. Lat. Am. J. Aquat. Res. 2021, 49, 418–430. [Google Scholar] [CrossRef]
- Duan, Y.; Wang, Y.; Liu, Q.; Xiong, D.; Zhang, J. Transcriptomic and microbiota response on Litopenaeus vannamei intestine subjected to acute sulfide exposure. Fish Shellfish. Immunol. 2019, 88, 335–343. [Google Scholar] [CrossRef]
- Duan, Y.; Wang, Y.; Huang, J.; Li, H.; Dong, H.; Zhang, J. Toxic effects of cadmium and lead exposure on intestinal histology, oxidative stress response, and microbial community of Pacific white shrimp Litopenaeus vannamei. Mar. Pollut. Bull. 2021, 167, 112220. [Google Scholar] [CrossRef]
- Zheng, X.; Duan, Y.; Dong, H.; Zhang, J. The effect of Lactobacillus plantarum administration on the intestinal microbiota of whiteleg shrimp Penaeus vannamei. Aquaculture 2020, 526, 735331. [Google Scholar] [CrossRef]
- Yu, Q.; Xie, J.; Huang, M.; Chen, C.; Qian, D.; Qin, J.G.; Chen, L.; Jia, Y.; Li, E. Growth and health responses to a long-term pH stress in Pacific white shrimp Litopenaeus vannamei. Aquac. Rep. 2020, 16, 100280. [Google Scholar] [CrossRef]
- Nouioui, I.; Carro, L.; García-López, M.; Meier-Kolthoff, J.P.; Woyke, T.; Kyrpides, N.C.; Pukall, R.; Klenk, H.-P.; Goodfellow, M.; Göker, M. Genome-Based Taxonomic Classification of the Phylum Actinobacteria. Front. Microbiol. 2018, 9, 2007. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Ma, S.; Liu, Y.; Pi, Y.; Zhao, Y. Intestinal microbial diversity and functional analysis of Urechis unicinctus under two different habitats: Pond poly-cultured with Penaeus japonicus and coastal zone. Aquac. Environ. Interactions 2021, 13, 211–224. [Google Scholar] [CrossRef]
- Tran, B.T.; Kim, K.-Y.; Heo, J.S.; Park, S.-J.; Park, H.K.; Choi, Y.H. Determination of the Pacific oyster Magallana gigas (Crassostrea gigas) diet composition in two aquaculture farms by fecal DNA metabarcoding. Aquaculture 2022, 552, 738042. [Google Scholar] [CrossRef]
- Li, S.; Wang, B.; Liu, L.; Song, Y.; Lv, C.; Zhu, X.; Luo, Y.; Cheng, C.H.K.; Chen, H.; Yang, X.; et al. Enhanced Growth Performance Physiological and Biochemical Indexes of Trachinotus ovatus Fed with Marine Microalgae Aurantiochytrium sp. Rich in n-3 Polyunsaturated Fatty Acids. Front. Mar. Sci. 2021, 7, 609837. [Google Scholar] [CrossRef]
- Hamamoto, Y.; Honda, D. Nutritional intake of Aplanochytrium (Labyrinthulea, Stramenopiles) from living diatoms revealed by culture experiments suggesting the new prey–predator interactions in the grazing food web of the marine ecosystem. PLoS ONE 2019, 14, e0208941. [Google Scholar] [CrossRef] [PubMed]
- Pauli, N.-C.; Metfies, K.; Pakhomov, E.A.; Neuhaus, S.; Graeve, M.; Wenta, P.; Flintrop, C.M.; Badewien, T.H.; Iversen, M.H.; Meyer, B. Selective feeding in Southern Ocean key grazers—Diet composition of krill and salps. Commun. Biol. 2021, 4, 1061. [Google Scholar] [CrossRef] [PubMed]
- Shaari, A.L.; Surif, M.; Latiff, F.A.; Omar, W.M.W.; Ahmad, M.N. Monitoring of Water Quality and Microalgae Species Composition of Penaeus monodon Ponds in Pulau Pinang, Malaysia. Trop. Life Sci. Res. 2011, 22, 51–69. [Google Scholar] [PubMed]
- Lukwambe, B.; Nicholaus, R.; Zhang, D.; Yang, W.; Zhu, J.; Zheng, Z. Successional changes of microalgae community in response to commercial probiotics in the intensive shrimp (Litopenaeus vannamei Boone) culture systems. Aquaculture 2019, 511, 734257. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/kg Dry Weight) | CD | D1 | D2 | D3 | D4 | D5 |
|---|---|---|---|---|---|---|
| Fishmeal 1 | 200.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| Soybean meal 2 | 277.1 | 319.3 | 316.1 | 316.8 | 319.7 | 316.2 |
| Poultry by-product meal 3 | 40.0 | 80.0 | 80.0 | 80.0 | 80.0 | 80.0 |
| Wheat flour 4 | 426.9 | 434.5 | 438.1 | 436.2 | 433.1 | 437 |
| Soy lecithin 5 | 35.0 | 35.0 | 35.0 | 35.0 | 35.0 | 35.0 |
| Fish oil 6 | 15.0 | 15.0 | 15.0 | 15.0 | 15.0 | 15.0 |
| Pellet binder 7 | 3.0 | 3.0 | 3.0 | 3.0 | 3.0 | 3.0 |
| Antioxidants (Calcium propionate) 8 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Minerals 9 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Vitamins 10 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Mono-dicalcium phosphate 11 | 0.0 | 2.7 | 2.7 | 2.7 | 2.7 | 2.7 |
| DL-Methionine 12 | 0.0 | 1.3 | 0.0 | 0.0 | 1.3 | 0.0 |
| Met-Met 13 | 0.0 | 0.0 | 0.6 | 1.9 | 0.0 | 0.6 |
| Biolys 77 14 | 0.0 | 4.9 | 5.0 | 5.0 | 4.9 | 5.0 |
| L-Threonine 15 | 0.0 | 1.2 | 1.3 | 1.3 | 1.2 | 1.3 |
| L-Tryptophan 16 | 0.0 | 0.1 | 0.2 | 0.2 | 0.1 | 0.2 |
| B. amyloliquefaciens CECT 5940 17 | 0.0 | 0.0 | 0.0 | 0.0 | 1.0 | 1.0 |
| Total (g) | 1000.0 | 1000.0 | 1000.0 | 1000.0 | 1000.0 | 1000.0 |
| Nutritional composition (%) | ||||||
| Dry matter | 90.4 | 90.4 | 90.4 | 90.4 | 90.4 | 90.4 |
| Crude protein | 35.0 | 32.9 | 32.8 | 32.9 | 32.9 | 32.8 |
| Crude lipid | 8.53 | 8.51 | 8.51 | 8.51 | 8.51 | 8.51 |
| Ash | 5.16 | 4.86 | 4.86 | 4.86 | 4.86 | 4.86 |
| Methionine | 0.7 | 0.7 | 0.7 | 0.9 | 0.7 | 0.7 |
| Lysine | 2.1 | 2.1 | 2.1 | 2.1 | 2.1 | 2.1 |
| Phosphorus | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 |
| Gross energy (kcal/kg) | 4.54 | 4.50 | 4.50 | 4.50 | 4.50 | 4.50 |
| Water stability | 0.95 ± 0.002 ab | 0.95 ± 0.006 ab | 0.94 ± 0.005 a | 0.95 ± 0.005 ab | 0.96 ± 0.003 b | 0.96 ± 0.003 b |
| Gene | Forward/Reverse Sequence | Amplicon Length (bp) | Efficiency (%) | Correlation Coefficient (R2) | GenBank Accession Number |
|---|---|---|---|---|---|
| β-actin | 5′-CCACGAGACCACCTACAAC-3′ 5′-AGCGAGGGCAGTGATTTC-3′ | 142 | 91 | 0.99 | AF300705 |
| hemocyanin (Hc) | 5′-GTCTTAGTGGTTCTTGGGCTTGTC-3′ 5′-GGTCTCCGTCCTGAATGTCTCC-3′ | 124 | 98 | 0.98 | X82502 |
| prophenoloxidase (proPO) | 5′-CGGTGACAAAGTTCCTCTTC-3′ 5′-GCAGGTCGCCGTAGTAAG-3′ | 122 | 99 | 0.99 | AY723296 |
| lipopolysaccharide- and β-glucan-binding protein (LGBP) | 5′-CCATGTCCGGCGGTGGAA-3′ 5′-GTCATCGCCCTTCCAGTTG-3′ | 122 | 110 | 0.99 | EU102286 |
| cytosolic manganese superoxide dismutase (cytMnSOD) | 5′-TGTTGCACAAGCCATTGACGA-3′ 5′-CCAGCCAGAGCCTTTCACTCC-3′ | 94 | 90 | 0.98 | DQ005531 |
| heat shock protein 60 (HSP60) | 5′-ATTGTCCGCAAGGCTATC-3′ 5′-ATCTCCAGACGCTTCCAT-3′ | 102 | 103 | 0.99 | FJ710169 |
| Diets | Initial Weight (g) | Final Weight (g) | Weekly Weight Gain (g/week) | Specific Growth Rate (%/day) | Survival Rate (%) | Final Biomass (g) | Feed Intake (g) | Feed Conversion Rate |
|---|---|---|---|---|---|---|---|---|
| CD | 0.28 ± 0.01 a | 5.00 ± 0.10 ab | 0.59 ± 0.01 ab | 5.17 ± 0.08 | 77.50 ± 5.95 | 77.64 ± 6.78 ab | 145.19 ± 11.06 ab | 1.87 ± 0.02 |
| D1 | 0.31 ± 0.02 a | 4.60 ± 0.38 a | 0.54 ± 0.05 a | 4.81 ± 0.28 | 73.75 ± 6.57 | 67.95 ± 8.42 a | 131.08 ± 17.28 a | 1.94 ± 0.16 |
| D2 | 0.29 ± 0.02 a | 5.34 ± 0.26 ab | 0.63 ± 0.03 ab | 5.21 ± 0.11 | 86.25 ± 7.74 | 93.21 ± 12.10 ab | 160.06 ± 14.88 ab | 1.75 ± 0.09 |
| D3 | 0.32 ± 0.02 a | 5.84 ± 0.25 b | 0.69 ± 0.03 b | 5.20 ± 0.18 | 95.00 ± 3.54 | 111.11 ± 7.02 b | 185.63 ± 7.15 ab | 1.68 ± 0.06 |
| D4 | 0.28 ± 0.02 a | 5.28 ± 0.10 ab | 0.63 ± 0.01 ab | 5.27 ± 0.12 | 83.75 ± 6.57 | 88.26 ± 6.64 ab | 163.87 ± 12.81 ab | 1.86 ± 0.06 |
| D5 | 0.32 ± 0.01 a | 5.98 ± 0.21 b | 0.71 ± 0.03 b | 5.22 ± 0.13 | 91.25 ± 3.75 | 108.94 ± 4.39 b | 190.83 ± 10.46 b | 1.75 ± 0.04 |
| p-value | 0.3196 | 0.0063 | 0.0109 | 0.4152 | 0.1507 | 0.0069 | 0.0285 | 0.3419 |
| Digestive Enzyme Activity [U/mg] | Dietary Treatment | ||||||
|---|---|---|---|---|---|---|---|
| CD | D1 | D2 | D3 | D4 | D5 | p Value | |
| Hepatopancreas | |||||||
| Amylase | 75.09 ± 3.16 a | 61.78 ± 1.87 a | 94.64 ± 9.12 a | 369.48 ± 4.73 b | 165.30 ± 70.30 a | 154.90 ± 28.38 a | 0.0010 |
| Protease | 0.14 ± 0.01 a | 0.16 ± 0.01 a | 0.42 ± 0.10 ab | 1.05 ± 0.21 b | 0.41 ± 0.22 ab | 0.33 ± 0.13 a | 0.0073 |
| Lipase | 1.32 ± 0.12 a | 0.80 ± 0.10 a | 1.05 ± 0.07 a | 5.53 ± 0.38 b | 1.86 ± 0.67 a | 2.70 ± 1.00 a | 0.0003 |
| Intestine | |||||||
| Amylase | 249.49 ± 8.91 | 202.36 ± 36.41 | 279.44 ± 48.10 | 291.38 ± 83.74 | 385.98 ± 104.15 | 405.49 ± 9.7 | 0.2044 |
| Protease | 16.80 ± 0.26 | 12.64 ± 3.14 | 18.18 ± 5.09 | 18.84 ± 2.06 | 22.62 ± 6.49 | 30.59 ± 2.1 | 0.0815 |
| Lipase | 1.48 ± 0.11 | 1.11 ± 0.25 | 1.61 ± 0.28 | 1.86 ± 0.40 | 2.39 ± 0.9 | 2.28 ± 0.08 | 0.3943 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casillas-Hernández, R.; Gonzalez-Galaviz, J.R.; Rodriguez-Anaya, L.Z.; Gil-Núñez, J.C.; Rodríguez-Jaramillo, M.d.C. Dietary Use of Methionine Sources and Bacillus amyloliquefaciens CECT 5940 Influences Growth Performance, Hepatopancreatic Histology, Digestion, Immunity, and Digestive Microbiota of Litopenaeus vannamei Fed Reduced Fishmeal Diets. Animals 2023, 13, 43. https://doi.org/10.3390/ani13010043
Casillas-Hernández R, Gonzalez-Galaviz JR, Rodriguez-Anaya LZ, Gil-Núñez JC, Rodríguez-Jaramillo MdC. Dietary Use of Methionine Sources and Bacillus amyloliquefaciens CECT 5940 Influences Growth Performance, Hepatopancreatic Histology, Digestion, Immunity, and Digestive Microbiota of Litopenaeus vannamei Fed Reduced Fishmeal Diets. Animals. 2023; 13(1):43. https://doi.org/10.3390/ani13010043
Chicago/Turabian StyleCasillas-Hernández, Ramón, Jose Reyes Gonzalez-Galaviz, Libia Zulema Rodriguez-Anaya, Juan Carlos Gil-Núñez, and María del Carmen Rodríguez-Jaramillo. 2023. "Dietary Use of Methionine Sources and Bacillus amyloliquefaciens CECT 5940 Influences Growth Performance, Hepatopancreatic Histology, Digestion, Immunity, and Digestive Microbiota of Litopenaeus vannamei Fed Reduced Fishmeal Diets" Animals 13, no. 1: 43. https://doi.org/10.3390/ani13010043