
A Brief Review of Meiotic Chromosomes in Early Spermatogenesis and Oogenesis and Mitotic Chromosomes in the Viviparous Lizard Zootoca vivipara (Squamata: Lacertidae) with Multiple Sex Chromosomes
Abstract
:Simple Summary
Abstract
1. Introduction
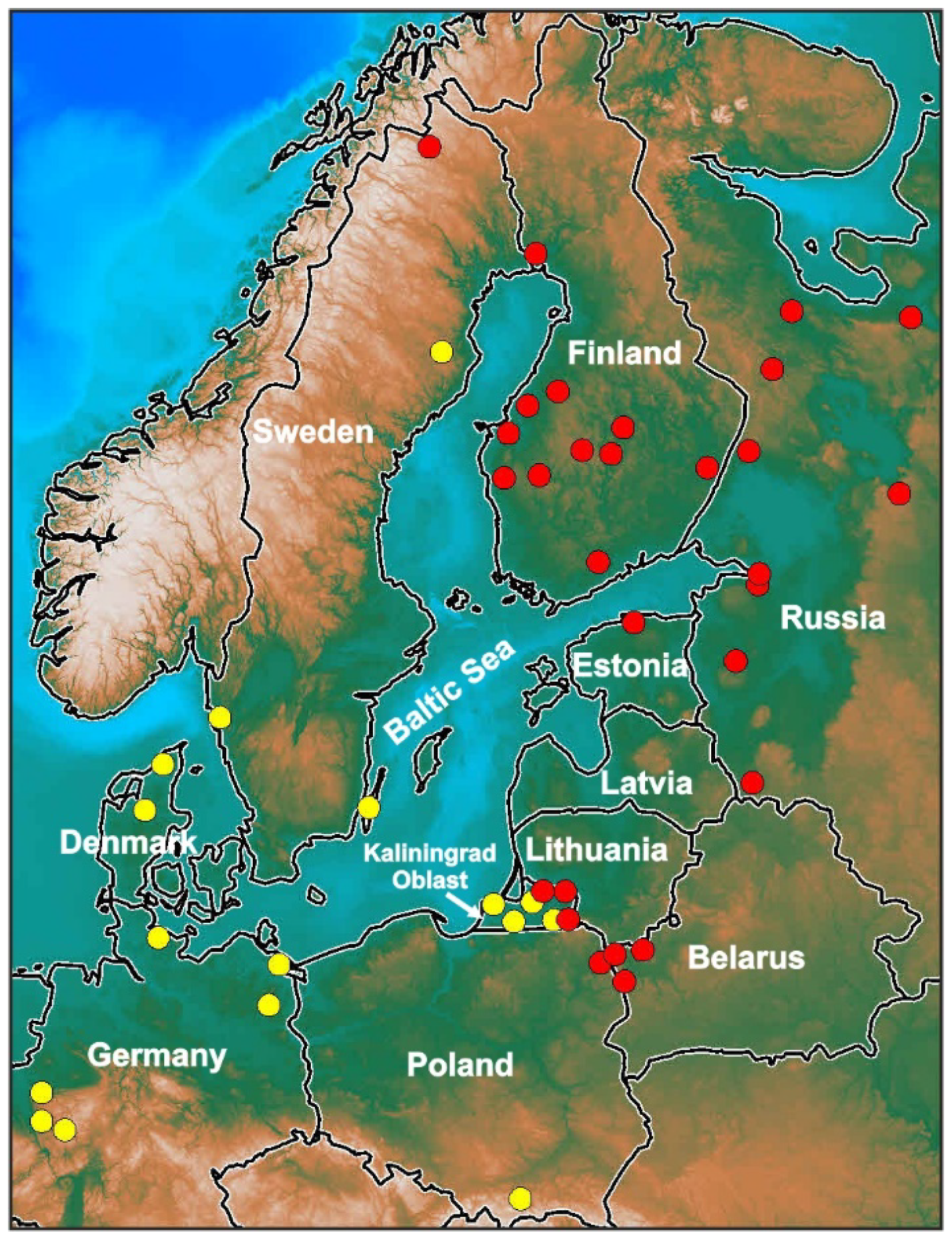
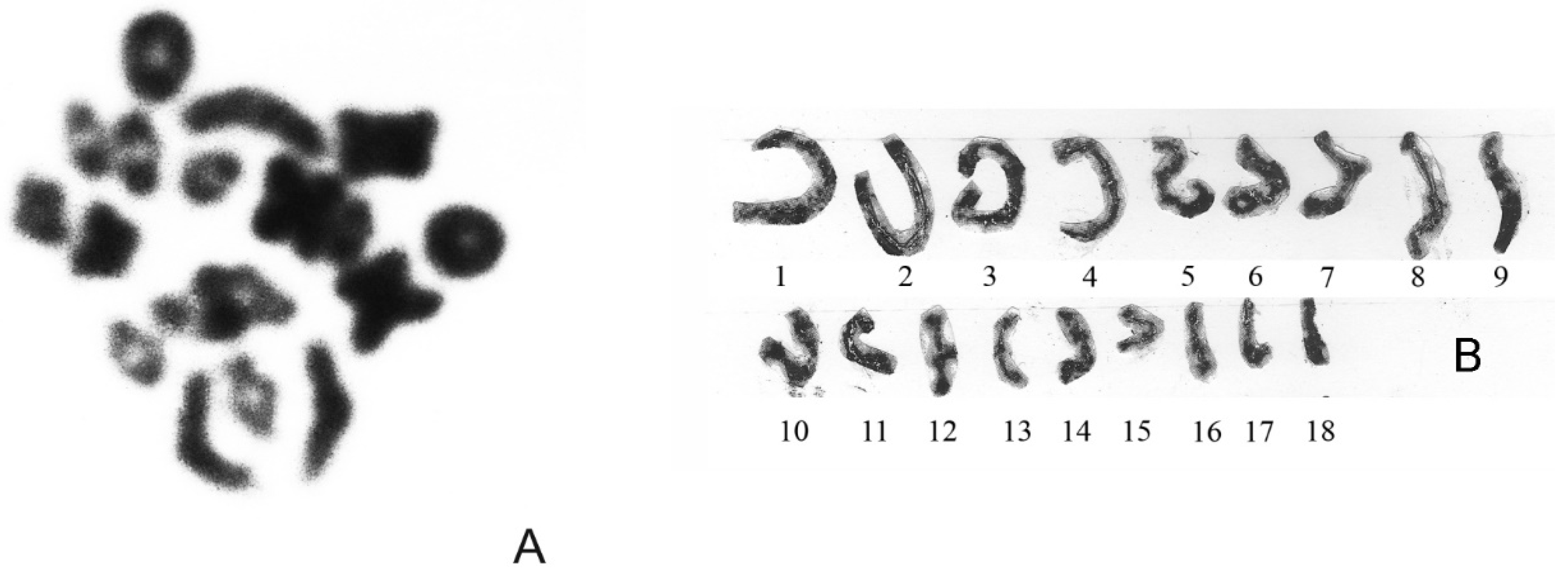
2. Characteristics of Karyotype
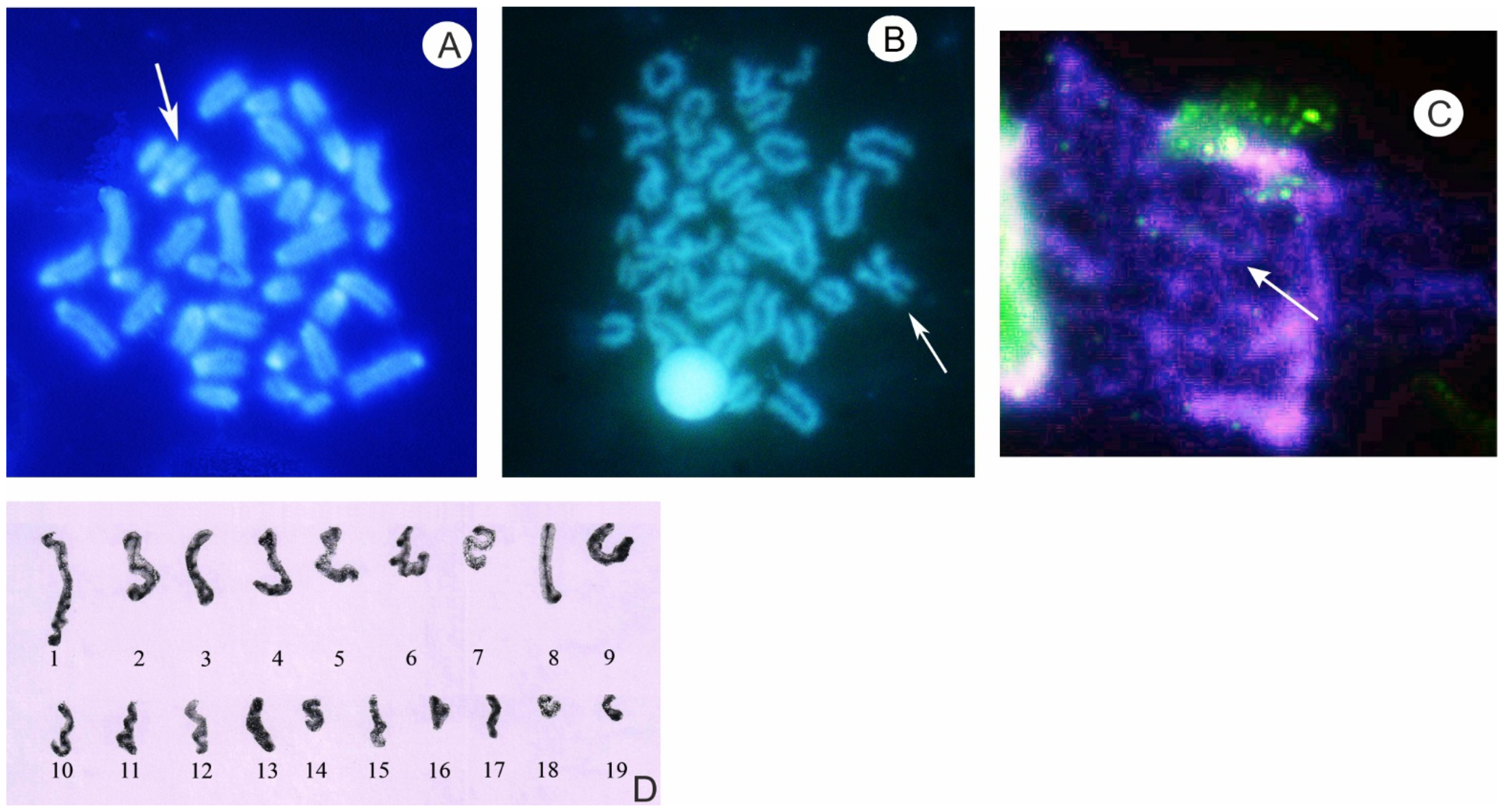
3. Characteristics of Meiosis in Spermatogenesis and Early Oogenesis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mezzasalma, M.; Guarino, F.; Odierna, G. Lizards as model organisms of sex Chromosome evolution: What we really know from a systematic distribution of available data? Genes 2021, 12, 1341. [Google Scholar] [CrossRef] [PubMed]
- Pokorná, M.; Almanová, M.; Kratochvil, L. Multiple sex chromosomes in the light of female meiotic drive in amniote vertebrates. Cytogenet. Genome Res. 2014, 22, 35–44. [Google Scholar] [CrossRef]
- Ezaz, T.; Sarre, S.; Meally, D.; Graves, J.M.M.; Georges, A. Sex chromosome evolution in lizards: Independent origins and rapid transitions. Cytogenet. Genome Res. 2009, 127, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Kitano, J. The contribution of female meiotic drive to the evolution of neo-sex chromosomes. Evolution 2012, 66, 3198–3208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kretschmer, R.; Ferguson-Smith, M.A.; De Oliveira, E.H.C. Karyotype evolution in birds: From conventional staining to chromosome painting. Genes 2018, 9, 181. [Google Scholar] [CrossRef] [Green Version]
- Pala, I.; Naurin, S.; Stervander, M.; Hasselquist, D.; Bensch, S.; Hansson, B. Evidence of a neo-sex chromosome in birds. Heredity 2011, 108, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Rovatsos, M.; Vukic, J.; Mrugala, A.; Suwala, C.; Lymberakis, P.; Kratochvil, L. Little evidence for switches to environmental sex determination and turnover of sex chromosomes in lacertid lizards. Sci. Rep. 2019, 9, 7832. [Google Scholar] [CrossRef] [Green Version]
- Rovatsos, M.; Almatová, M.; Augstenová, B.; Mazzoleni, S.; Velenský, P.; Kratochvil, L. ZZ/ZW sex determination with multiple neo-sex chromosomes in common in madagascan chameleons of the genus Furcifer (Reptilia: Chamaeleonidae). Genes 2019, 10, 1020. [Google Scholar] [CrossRef] [Green Version]
- Olmo, E.; Odierna, G.; Capriglione, T.; Cardone, A. DNA and chromosome evolution in lacertid lizards. In Book Cytogenetics of Amphibians and Reptiles, ALS; Olmo, E., Ed.; Birkhäuser Verlag: Basel, Switzerland; Boston, MA, USA; Berlin, Germany, 1990; pp. 181–204. [Google Scholar]
- Kupriyanova, L.; Rudi, E. Comparative karyological analysis of Lacerta vivipara (Lacertidae, Sauria) populations. Zool. J. 1990, 69, 93–101. (In Russian) [Google Scholar]
- Odierna, G.; Aprea, G.; Capriglione, T.; Arribas, O.; Kupriyanova, L.; Olmo, E. Progressive differentiation of the W sex chromosome between viviparous ndoviparous populations of Zootoca vivipara (Reptilia, Lacertidae). Ital. J. Zool. 1998, 65, 295–302. [Google Scholar] [CrossRef]
- Kupriyanova, L.; Odierna, G.; Caproglione, T.; Olmo, E.; Aprea, G. Chromosomal changes and form-formation, subspeciation in the wideranged Europasian species Zootoca vivipara (evolution, biogeography). Herpetologia Petropolitana. In Proceedings of the 12th Ordinary General Meeting of the Societas Europaea Herpetologica, St. Petersburg, Russia, 12–16 August 2003; Ananjeva, N., Tsinenko, O., Eds.; Folium: St. Petersburg, Russia, 2003; Volume 12, pp. 47–52. [Google Scholar]
- Kupriyanova, L.; Böhme, W. A review of the cryptic diversity of the viviparous lizard, Zootoca vivipara (Lichtenstein, 1823) (Squamata: Lacertidae) in Central Europe and its postglacial re-colonization out of the Carpathian basin: Chromosomal and molecular data. In Book Europe: Environmental, Political and Social Issues; Thygesen, E., Ed.; Nova Science Publisher: New York, NY, USA, 2020; Chapter 2; pp. 25–43. [Google Scholar]
- Surget-Groba, Y.; Heulin, B.; Guillaume, C.-P.; Puky, M.; Semenov, D.; Orlova, V.; Kupriyanova, L.; Ghira, I.; Smajda, B. Multiple origins of viviparity, or reversal from viviparity to oviparity and the evolution of parity. Biol. J. Linn. Soc. 2006, 87, 1–11. [Google Scholar] [CrossRef]
- Velekei, B.; Lakatos, F.; Biro, P.; Acs, E.; Puky, M. The genetic structure of Zootoca vivipara (Lichtenstein, 1823) populations did not support the existence of a north-south corridor of the VB haplogroup in eastern Hungary. North-West. J. Zool. 2014, 10, 187–189. [Google Scholar]
- Kupriyanova, L.; Kuksin, A.; Odierna, G. Karyotype, chromosome structure, reproductive modalities of three souuuuthern Eurasia populations of the common lacertid lizards, Zootoca vivipara (Jacquin, 1787). Acta Herpetol. 2008, 3, 99–106. [Google Scholar]
- Kupriyanova, L.; Melashchenko, O. The common Eurasian lizard Zootoca vivipara (Jacqouin, 1787) from Russia: Sex chromosomes, subspeciation, and colonization. Russ. J. Herpetol. 2011, 18, 99–104. [Google Scholar]
- Kupriyanova, L.; Kirschey, T.; Böhme, W. Distribution of the common or viviparous lizard, Zootoca vivipara (Lichtenstein, 1823) (Squamata: Lacertidae) in Central Europe and re-colonization of the Baltic Sea basin: New karyological evidence. Russ. J. Herpetol. 2017, 24, 311–317. [Google Scholar] [CrossRef]
- King, M. The evolution of sex chromosomes in lizards. In Evolution and Reproduction; Australian Academy of Science: Canberra, Australia, 1977; pp. 55–60. [Google Scholar]
- King, M. Species Evolution. The Role of Chromosome Change. Cambridge University Press: Cambridge, UK, 1993; 336p. [Google Scholar]
- Payseur, B.; Presgraves, D.; Filatov, D. Sex chromosomes and speciation. Mol. Ecol. 2018, 27, 3745–3748. [Google Scholar] [CrossRef]
- Pennell, M.W.; Mank, J.E.; Peichel, C.L. Transition in sex determination and sex chromosomes across vertebrate species. Mol. Ecol. 2018, 27, 3950–3963. [Google Scholar] [CrossRef] [Green Version]
- Olmo, E. Rate of chromosome changes and speciation in reptiles. Genetica 2005, 125, 185–203. [Google Scholar] [CrossRef]
- Petraccioli, A.; Guarino, F.; Kupriyanova, L.; Mezzasalma, M.; Odierna, G.; Picariello, O.; Capriglione, T. Isolation and characterization of interspersed repeated sequences in the European common lizard, Zootoca vivipara, and their conservation in Squamata. Cytogenet. Genome Res. 2019, 157, 65–76. [Google Scholar] [CrossRef]
- Safronova, L.D.; Kupriyanova, L.A. Metaphase and meiotic chromosomes, Synaptonemal complexes (SC) of the lizard Zootoca vivipara. Russ. J. Genet. 2016, 52, 1184–1191, ISSN 1022-7954. [Google Scholar] [CrossRef]
- Kupriyanova, L.A.; Safronova, L.D.; Chekunova, A.I. Meiotic chromosomes, Synaptonemal complexes (SC) in a female viviparoys lizard Zootoca vivipara. Russ. J. Genet. 2019, 55, 774–778, ISSN 1022-7954. [Google Scholar] [CrossRef]
- Summer, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Guttembach, M. Evolutionary diversity of reverse (R) fluorescent chromosome bands in vertebrates. Chromosoma 1988, 97, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Dresser, M.; Moses, M. Synaptonemal complex karyotyping in spermatocytes of the Chinesehamster (Cricetulus gricus).iY. Light and electron microscopy of synapsis and nucleolar development by silver staining. Chromosoma 1980, 76, 1–22. [Google Scholar] [CrossRef]
- Kupriyanova, L.A.; Safronova, L.D. Results and perspectives of cyto- and genetic studying of “cryptic” group of the Lacertidae. Trudyzin 2020, 324, 100–107. [Google Scholar] [CrossRef]
- Dinkel, B.J.; O’Laughlin-Phillips, E.A.; Fechheimer, N.S.; Jaap, R.G. Gametic products transmitted by chickens heterozygous for chromosomal rearrangements. Cytogenet. Cell. Genet. 1979, 23, 124–136. [Google Scholar] [CrossRef]
- De Villena, F.P.-M.; Sapienza, C. Female meiosis drives karyotypic evolution in mammals. Genetics 2001, 159, 1179–1189. [Google Scholar] [CrossRef]
- Kupriyanova, L.; Safronova, L.; Chekunova, A.; Osipov, F. The study of oocytes in the early oogenesis of the viviparous lizard (Zootoca vivipara) in prophase of its meiosis. Bull. Mosc. Soc. Nat. 2021, 126, 3–11. [Google Scholar]
- Kupriyanova, L.; Safronova, L.; Kirschey, T.; Böhme, W. Meiotic chromosomes, Synaptonemal complexex (SC) in the cryptic forms of the euroasian complex Zootoca vivipara with multiple sex chromosomes. In Book Abstract: The 12st European Congress of herpetology, Belgrade, Serbia 5–9 September 2022; Crnobrnja-Isailović, J., Vukov, T., Vučić, T., Tomović, L., Eds.; Institute for Biological Research “Siniša Stanković”, National Institute of Republicnof Serbia, University of Belgrade: Belgrade, Serbia, 2022; Volume 82, p. 261. [Google Scholar]
- Capriglione, T.; Olmo, E.; Odierna, G.; Kupriyanova, L. Mechanisms of differentiation in the sex chromosomes of some Lacertidae. Amphibia-Reptilia 1994, 15, 1–8. [Google Scholar]
- Chalopin, D.; Naville, M.; Plard, F.; Galiana, D.; Volff, J.N. Comparative analysis of transposable elements highlights mobilome diversity and evolution in vertebrates. Genome Biol. Evol. 2015, 7, 567–580. [Google Scholar] [CrossRef]
- Serrato-Capuchina, A.; Matute, D. The role of transposable elements in speciation. Genes 2018, 9, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, S.M.I.; Sarre, S.; Gleeson, D.; Georges, A.; Ezaz, T. Did lizards follow unique pathways in sex chromosome evolution? Genes 2018, 9, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakloushinskaya, I. Chromosomal rearrangements, reorganization of genome, and Speciation. Zool. J. 2016, 95, 376–393. [Google Scholar] [CrossRef]
- Axelsson, E.; Alrechtsen, A.; van A., P.; Li, L.; Megens, H.J.; Vereijken, A.L.J.; Crooijmans, R.P.M.A.; Groenen, M.A.M.; Ellegren, H.; Wierslev, E.; et al. Segregation distortion in chicken and the evolutionary consequences of female meiotic drive in birds. Heredity 2010, 105, 290–298. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| № | 2n ♂/♀ | System of Sex Chromosomes ♂/♀ | Morphology of w/W Sex Chromosomes | Mode of Reproduction, O/V (Ovi-/Viviparous) | Localities | Species, Subspecies, Chromosomal Forms |
|---|---|---|---|---|---|---|
| The first group of karyotype | ||||||
| 1. | 36A/36A | ZZ/Zw | M | O | Central, southwestern Europe | Z. vivipara, now Z. carniolica |
| 2. | 36A/36A | ZZ/Zw | M | V | Central Europe | Z. vivipara, now Z. vivipara Hungarian form |
| The second group of karyotype | ||||||
| 3. | 36A/35 (34A + 1 A/ST) | Z1Z1Z2Z2/Z1Z2W | A, ST | O | Western Europe | Z. vivipara, now Z. v. louislantzi Pyrenean form |
| 4. | 36A/35 (34 A + 1 A/ST) | Z1Z1Z2Z2/Z1Z2W | A/ST | V | Central Europe | Z. vivipara, now Z. vivipara Austrian form? |
| 5. | 36A/35 (34A + 1 SV) | Z1Z1Z2Z2/Z1Z2W | SV/ST | V | Central Europe | Z. vivipara, now Z. vivipara Romanian form |
| 6. | 36A/35 (34A + 1 A/ST) | Z1Z1Z2Z2/Z1Z2W | A/ST | V | Eastern Europe, eastern Baltic region, Asia | Z. vivipara, now Z. vivipara Eastern (Russian) form |
| 7. | 36A/35 (34A + 1 SV) | Z1Z1Z2Z2/Z1Z2W | SV | V | Western, Central Europe, Baltic region | Z. vivipara, now Z. vivipara Western form |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kupriyanova, L.; Safronova, L. A Brief Review of Meiotic Chromosomes in Early Spermatogenesis and Oogenesis and Mitotic Chromosomes in the Viviparous Lizard Zootoca vivipara (Squamata: Lacertidae) with Multiple Sex Chromosomes. Animals 2023, 13, 19. https://doi.org/10.3390/ani13010019
Kupriyanova L, Safronova L. A Brief Review of Meiotic Chromosomes in Early Spermatogenesis and Oogenesis and Mitotic Chromosomes in the Viviparous Lizard Zootoca vivipara (Squamata: Lacertidae) with Multiple Sex Chromosomes. Animals. 2023; 13(1):19. https://doi.org/10.3390/ani13010019
Chicago/Turabian StyleKupriyanova, Larissa, and Larissa Safronova. 2023. "A Brief Review of Meiotic Chromosomes in Early Spermatogenesis and Oogenesis and Mitotic Chromosomes in the Viviparous Lizard Zootoca vivipara (Squamata: Lacertidae) with Multiple Sex Chromosomes" Animals 13, no. 1: 19. https://doi.org/10.3390/ani13010019