1. Introduction
Ascites syndrome (AS) is also called broiler pulmonary hypertension syndrome. It is a common metabolic disorder, with main clinical symptoms including right heart hypertrophy and thinning, lung and liver congestion, edema, and hematoma [
1]. Numerous studies have shown that hypoxia is the main cause of AS, and pulmonary hypertension is an important link in the pathogenesis of AS. Fast-growing broilers have a high metabolic rate, but the cardiopulmonary function of broilers is insufficient to support this and cannot provide enough oxygen to meet the metabolic demand of fast growth, resulting in relative hypoxia in the body. Hypoxia stimulates the formation of compensatory mechanisms in the cardiopulmonary system, including increased pulmonary vascular resistance [
2], oxygen free radical production [
3], and metabolic reprogramming [
4], leading to pulmonary hypertension and further leading to the occurrence of AS. Although the pathological progression of AS has been studied for many years, the molecular mechanism of its occurrence and development has not been fully elucidated. Davoodi and Ehsani (2019) identified 24 genes that play a role in AS based on published studies, including
MPPK2,
AT1,
RhoGTPase,
MC4R,
CDH6,
NOS3,
HIF-
1A,
OSBL6,
CCDC141,
BMPR2,
LEPR,
AGTR1,
UTS2D,
5HT2b,
SST,
CHRD,
TFRC,
CDH13,
ACVRL1,
Arnt,
ACE,
ACVRL1,
MEF2C, and
HTR1A [
5]. A recent study found that
ALDH7A1,
IRG1,
GGT5,
IGSF1,
DHX58,
USP36,
TREML2,
SPAG1,
CD34, and
PLEKHA7 are closely related to the pathogenesis of pulmonary artery remodeling in the progression of AS [
6].
Both respiratory and circulatory systems are important for broiler development. The lack of any circulation between the lung, heart, and vascular system will lead to the above pathological cascade reaction, leading to the occurrence of AS in broilers [
2]. Therefore, the lung is the main target organ of AS. It has been shown that in the lungs of rapidly growing broilers, all available blood vessels appear congested, demonstrating a very limited ability of the broiler lung to utilize key compensatory mechanisms. In addition, the lung plays an important role in many other processes, including the metabolism of endogenous and exogenous compounds, the regulation of water-fluid metabolism, and the production of a wide variety of signaling molecules (as a para-endocrine organ), including growth factors and cytokines [
7,
8]. The lung comprises various cell types (fibroblasts, airway smooth muscle cells, blood vessels, nerve cells, alveolar macrophages, mast cells, and dendritic cells) and the extracellular matrix. These cells constantly interact with extracellular components to achieve dynamic regulation of various functions. Any disruption of these interactions can lead to diseases (including asthma, chronic obstructive pulmonary disease (COPD), acute lung injury (ALI), pulmonary hypertension, and cancer) [
7,
9]. To date, several studies have been conducted on broilers with ascites, covering the liver [
10], heart [
11,
12], and vasculature [
13]. However, the transcriptome of lung tissue in broilers with AS has not been studied.
We used high-throughput sequencing (RNA-seq) to comprehensively study the lung changes of broilers with AS at the transcriptome level and identified important pathways and significantly differentially expressed genes in lung tissue during the development of AS. This study aims to reveal the differential expression of lung genes and pathways in broilers, which is of great significance for studying the pathogenesis and prevention of AS.
4. Discussion
The pathogenesis of AS in broilers may be due to insufficient circulation between the heart, lung, and vascular systems to meet the metabolic needs of broilers. To some extent, defects in one system can prevent the other parts from being fully compensated, thus triggering a pathological cascade that eventually leads to ascites [
16]. The lung is an important target organ of AS [
17]. Hasanpur et al. (2019) found that lung weight, as an indicator of respiratory capacity, was the most important factor affecting the sensitivity of broilers to AS [
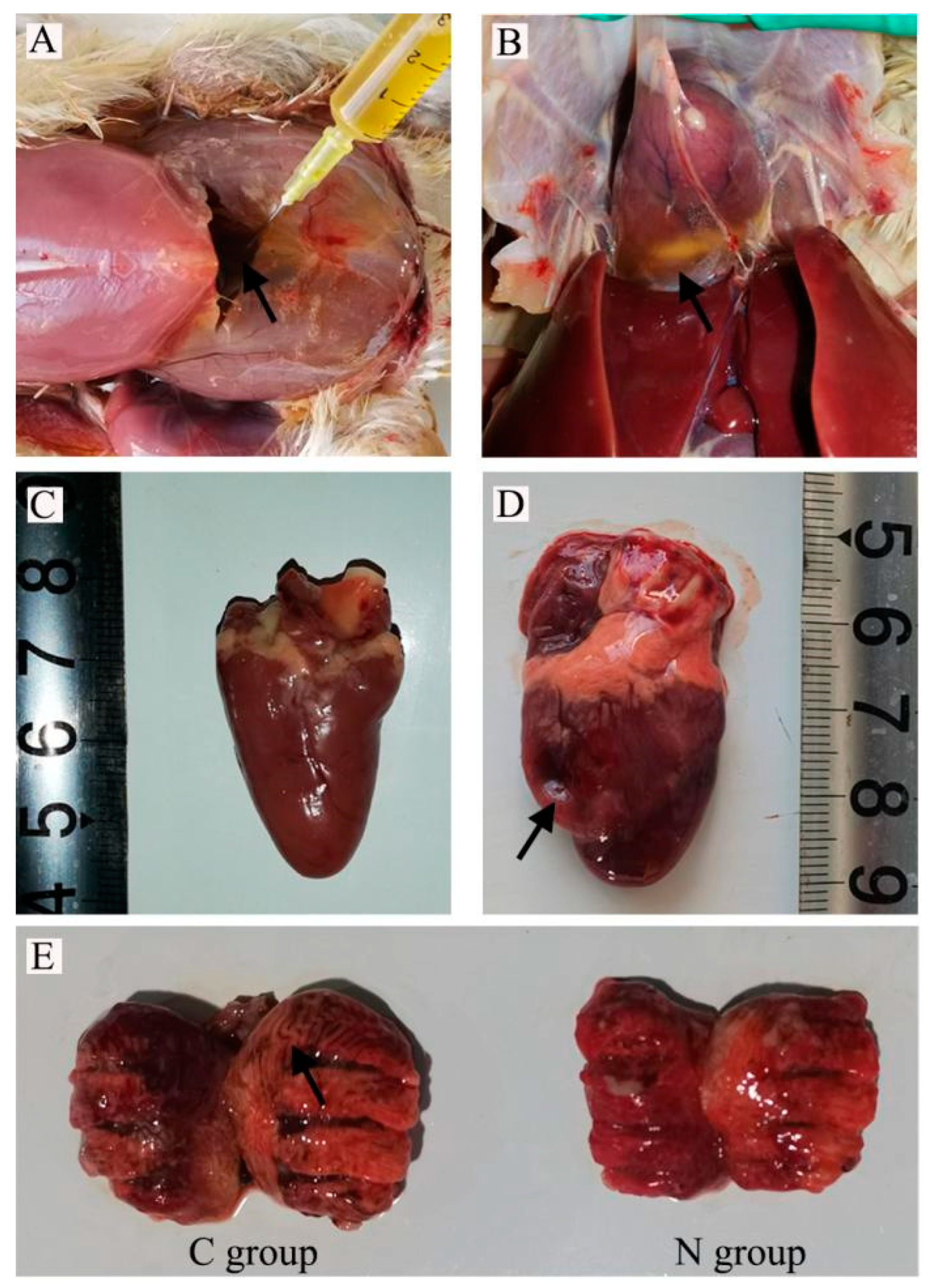
11]. It is consistent with the results of this study. Therefore, decreased respiratory capacity is the most likely factor to induce AS. In the current study, the average body weight of broilers in group C decreased, AHI increased, and AHI values more than 0.29. Compared with group N, the blood routine indexes increased, the levels of HCT and HB increased, and there was yellow fluid accumulation in the abdominal cavity and pericardium. Thus, the AS broiler model group was successfully established. In the current study, AHI (RV/TV) > 0.29, HCT, and HGB levels were increased compared with normal chickens, and there was yellow fluid accumulation in the abdominal cavity and pericardium. Hence, the AS broiler model group was successfully constructed. The lung tissue of broilers with ascites was congested, edematous, mottled, and slightly hard in texture, with grayish-white streaks in the interstitium (
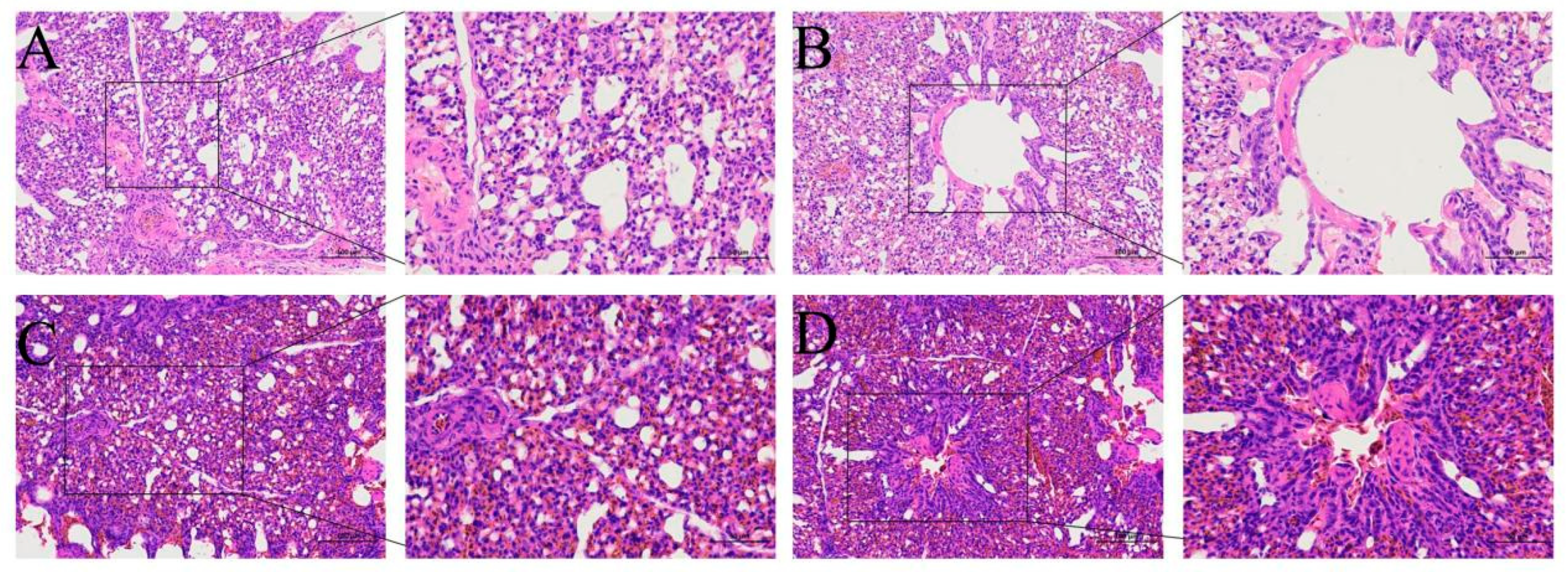
Figure 1a). The pathological changes in the lung were confirmed by the tissue sections. There was a tendency for the pulmonary vascular walls to thicken; the lung chamber was narrow, and there was respiratory capillary atrophy. Furthermore, there were pulmonary congestion, edema, and signs of inflammation in the lungs.
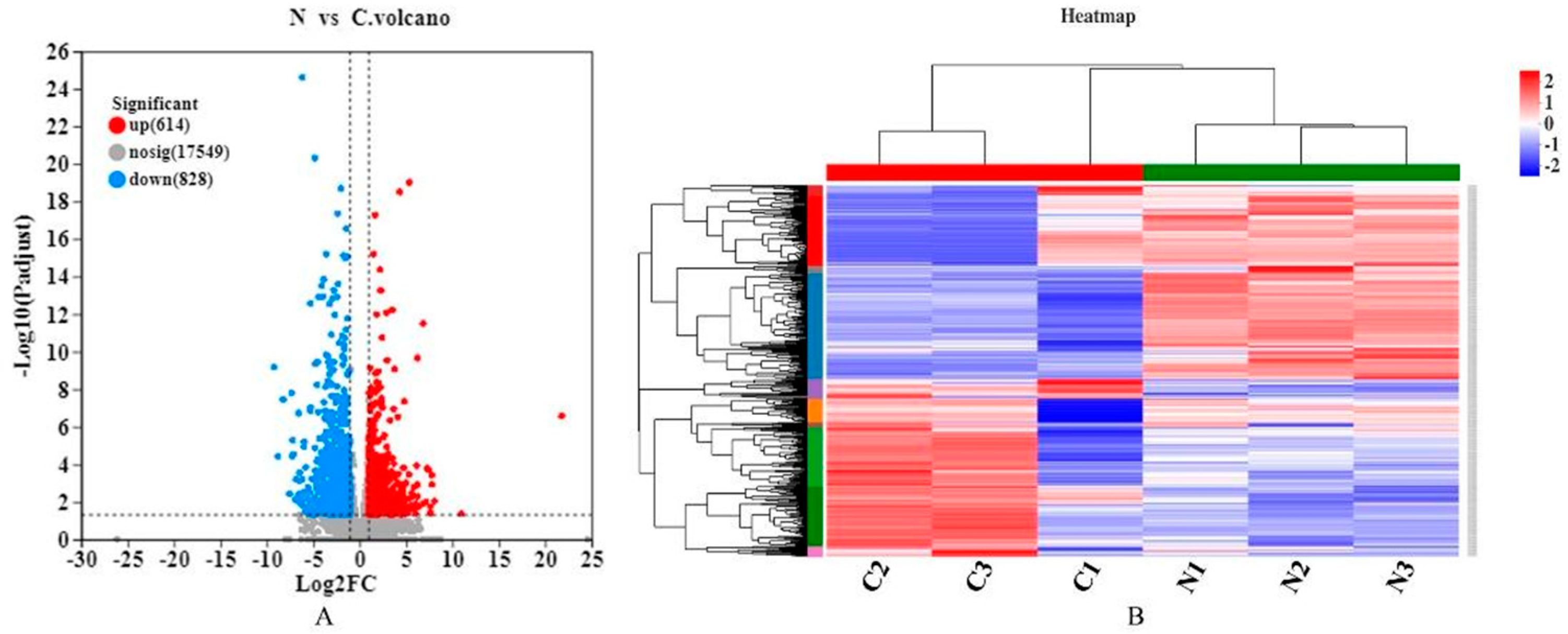
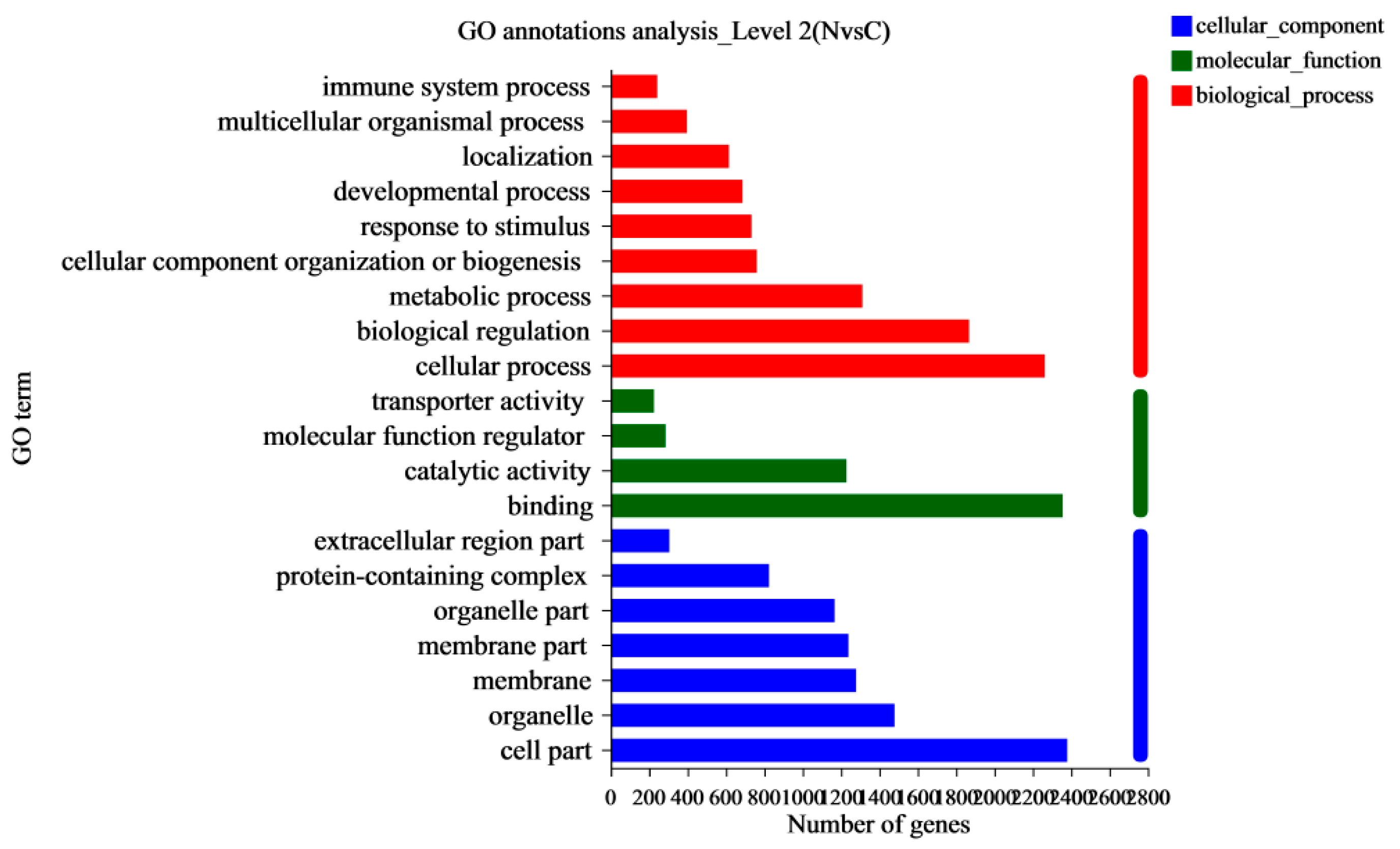
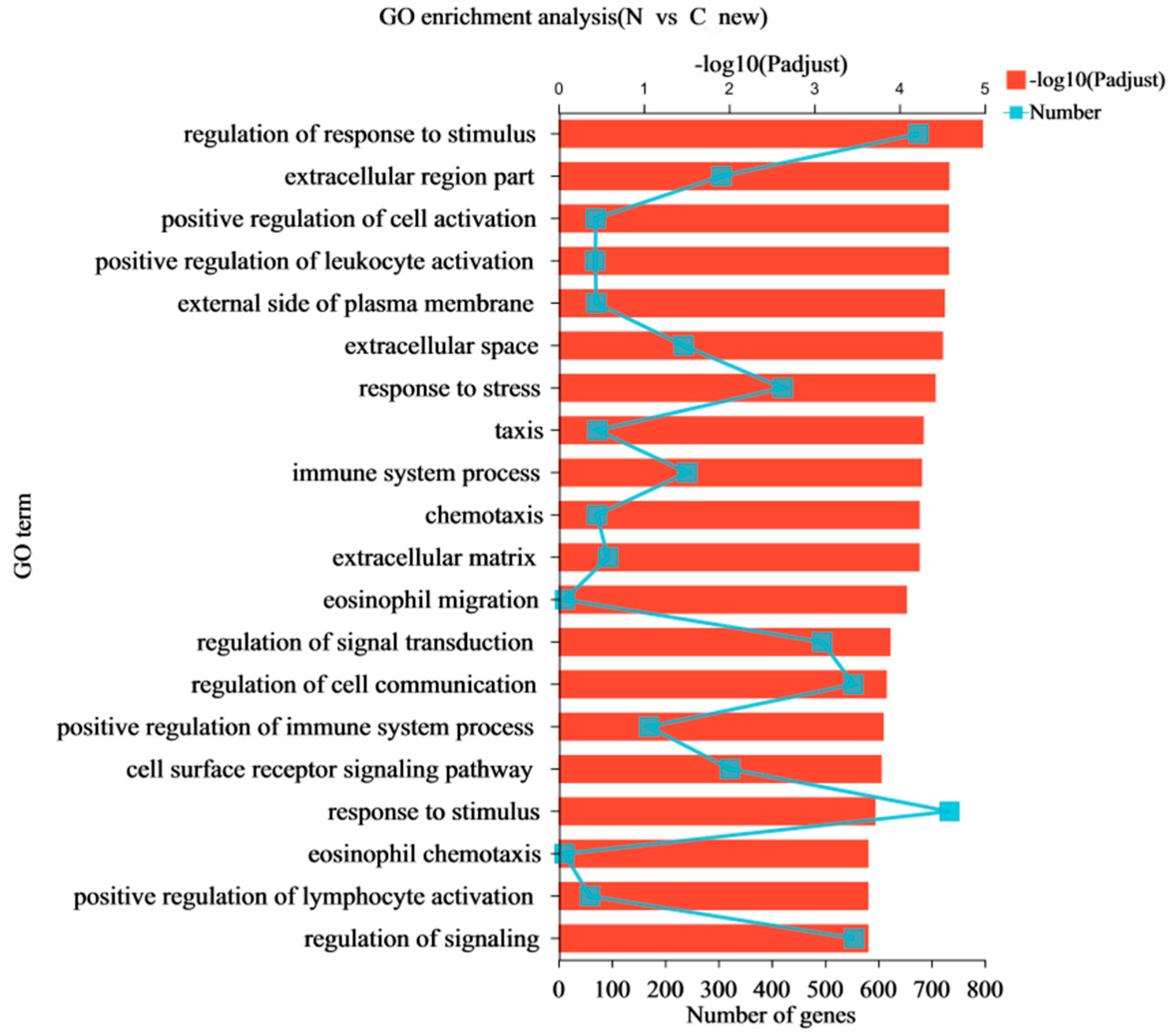
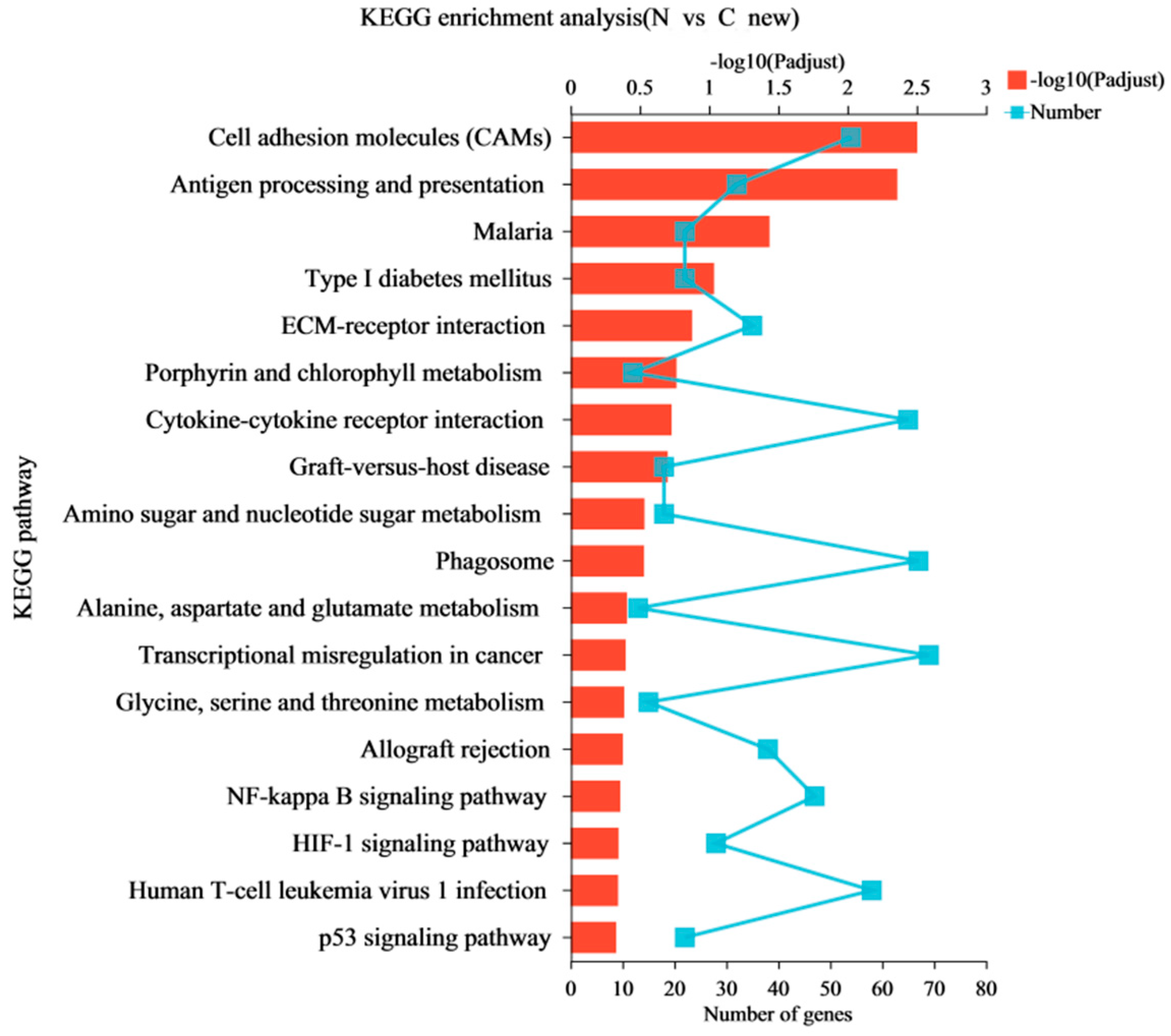
In this study, RNA-Seq was performed on the lung tissues of the normal group (group N) and the AS model group (group C) to identify the differential expression of possible candidate genes and pathways in the lung tissues and to elucidate the pathological mechanism of AS in broilers. DEGs and different regulatory pathways that may be involved in the pathogenesis of AS were found in the GO enrichment and KEGG enrichment pathways. We found that “Cell adhesion molecules (CAMs)” and the HIF-1, NF-κB, and p53 signaling pathways were enriched in AS. It has been shown that endothelial cells overexpress adhesion molecules in response to stimuli, such as turbulent flow, dyslipidemia, or increased luminal pressure. The Refmir test has proven that increased blood pressure is associated with increased serum CAMs concentration and decreased circulating leukocyte ligands freely accessible by CAMs [
18]. In the current study, the CAMs pathway was most significantly enriched, possibly in response to endothelial dysfunction or vascular injury. In addition, sequencing results in this study showed that the HIF-1 signaling pathway was very important in the occurrence and development of AS. In lung tissue, HIF-1 signaling orchestrates the body’s physiological response to oxygen. By participating in nitric oxide (NO) and the corresponding metabolic changes, the HIF signaling pathways increase pulmonary hypertension (pulmonary artery hypertension, PAH) vascular lesions [
19]. NF-κB and p53 signaling pathways interact with each other to affect hypoxic PAH, thus playing a role in the occurrence and development of AS. Compared to wild-type mice, p53-knockout mice exhibit more severe PAH in response to chronic hypoxia [
20]. Studies have found that TNF-activated NF-κB and p53 signaling pathways regulate the abnormal function of platelet mesenchymal stem cells, thereby improving hypoxic PAH [
21]. In addition, studies have shown that persistent hypoxia can induce the accumulation of leukocytes and mesenchymal progenitor cells in pulmonary arteries, promoting the formation of a pulmonary artery-specific chronic inflammatory microenvironment [
22]. The inflammatory mechanism of chemokine, fractalkine, and its receptor, Cx3CR1, in the lungs of patients with pulmonary hypertension may play a role in the natural development of pulmonary hypertension, proving the possibility of an inflammatory component in pulmonary hypertension [
23].
The “HIF-1 signaling pathway” plays an important role in regulating the AS hypoxic stress response in broilers. The pathology of rapid development of AS is caused by hypoxia. Hypoxia-inducible factors (HIFs) are major regulators of angiogenesis, erythropoiesis, and metabolism regulation [
24], and their expression is substantially increased in the lungs and heart of broilers with AS [
25]. The HIF-1α subunit is a unique functional subunit of HIF-1, which is closely related to hypoxia. The ratio of lung volume to body weight of broilers is small, implying that the respiratory system cannot respond to the high oxygen demand of broilers, leading to hypoxia and increased oxidative stress [
26]. HIF-1 is a key transcriptional factor of hypoxia-induced genes, which can initiate a vast hypoxic stress regulatory network under hypoxia.
Several pathways related to metabolic processes were enriched in this study, including amino sugar and nucleotide-sugar metabolism, alanine-aspartate-glutamate, and glycine-serine-threonine metabolism. A growing number of studies have identified a pervasive feature of metabolic and bioenergetic alterations in human and animal models of pulmonary hypertension, as metabolic abnormalities have been found in the lungs and hearts of patients, in animal models of this disease, and in cells derived from the lungs of patients. Stenmark et al. proposed that metabolic reprogramming is involved in the pathogenesis of PAH [
4,
27]. Metabolic reprogramming refers to the rearrangement of disordered metabolic pathways in the body in response to changes in the internal and external environment, i.e., the reintegration of metabolic systems to provide energy for cells [
28]. Key features of PAH metabolism include shifts in glycolysis, increases in glutamine availability and single-carbon metabolism, and decreases in fatty acid oxidation. Increased reactive oxygen species (ROS) production and changes in the tricarboxylic acid cycle (TCA) intermediates may contribute to stabilizing hypoxia-inducible factors driving the observed metabolic changes. Arginine metabolism is related to NO production and the TCA cycle and plays a key role in the relationship between a vasoconstrictive phenotype and metabolic abnormalities in diseases [
29]. In addition, the metabolism of alanine, aspartate, and glutamate is related to arginine biosynthesis [
30]. The two pathways of arginine metabolism include the conversion of arginine to NO and the decomposition of arginine to urea and ornithine, which are regulated in the inflammatory response [
31].
Serine/glycine metabolism is very important in biological metabolism because it provides one carbon unit, synthesizes other amino acids, nucleotides, and lipids, and synthesizes reducing substances to make REDOX in the cell in a state of dynamic equilibrium [
32,
33]. In recent years, studies have shown that serine/glycine metabolism plays a crucial role in tumors, affecting the occurrence and development of tumors and the therapeutic effect [
34]. Metabolic remodeling can occur by promoting the proliferation of pulmonary artery smooth muscle cells (PASMCS), inducing pulmonary vascular remodeling, thus accelerating the development of PAH [
35]. Metabolites are interrelated and mutually restricted. The change in one substance may be associated with the change in many substances, thus promoting the pathological process of PAH from different aspects. To elucidate the metabolic mechanism of AS in broilers by analyzing abnormal metabolic changes provides more targets for the prevention and control of AS.
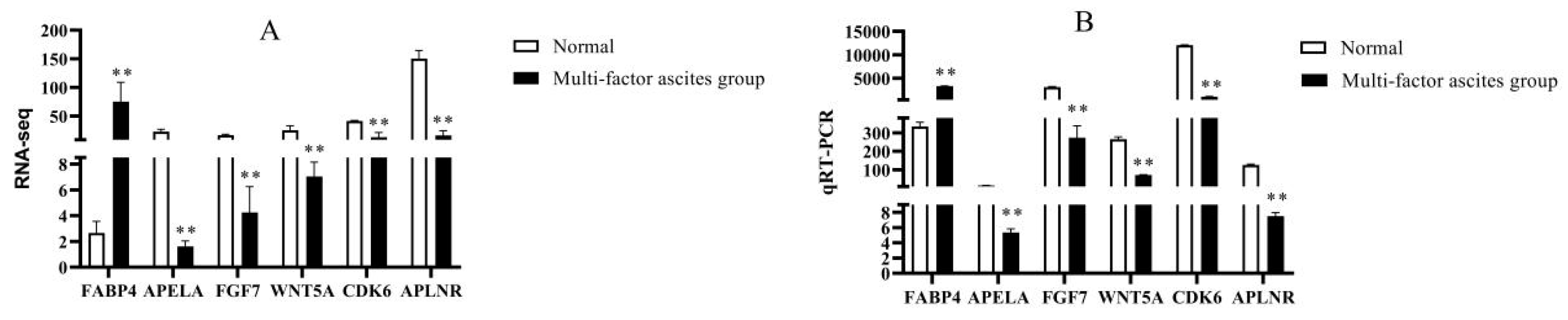
In this study, in addition to DEGs significantly enriched in GO terms and KEGG pathways, we also found other important DEGs,
FABP4,
APLNR,
APLN,
APELA,
EIF2AK4,
HMOX1,
THBS1,
TLR4,
BCL2,
FGF7,
WNT5A, and
CDK6. The highly up-regulated differential gene,
FABP4, also known as fatty acid binding protein 4, is mainly expressed in adipocytes and macrophages and is involved in regulating glucose and lipid metabolism related to inflammation and metabolic processes in target cells. FABP4 is associated with obesity, insulin resistance, diabetes mellitus, hypertension, cardiac dysfunction, atherosclerosis, and cardiovascular events [
36,
37]. FABP4 enhances hepatic glucose production in vivo and in vitro, reduces cardiomyocyte contraction in vitro, inhibits endothelial nitric oxide synthase (eNOS) expression/activation in vascular endothelial cells, increases vascular smooth muscle cell proliferation/migration and glucose-stimulated insulin secretion in pancreatic β-cells. FABP4 may also play a role in atherosclerosis through TLRs and other mechanisms [
36,
38]. Accumulating evidence for the pathogenic role of FABP4 in cardiovascular events suggests that FABP4 is a potential new target for preventing cardiovascular diseases. APLN is a novel peptide that is an endogenous ligand for the G protein-coupled receptor APLNR [
39]. APLN–APLNR plays a key role in cardiac development, angiogenesis, NO-dependent vasodilation, and muscle strength signaling. APLN and APLNR are regulated by hypoxia, and the APLN–APLNR axis system has a protective effect on cardiovascular diseases [
40]. APLN may inhibit the expression of fibroblast growth factor 2 (FGF2) and FGF receptor 1 (FGFR1) by regulating the expression of miR-424 and miR-503, thereby improving PAH [
41]. In addition, APLN can regulate myocardial metabolism and glucose metabolism. Apelin treatment can substantially reduce the increase in myocardial free fatty acid (FFA) and glycogen content. In the treatment of type 2 diabetic rats, Apelin substantially reduced gene expression and increased the expression of CD36CPT-1 and PPAR-α in the heart fatty acid transporter [
42]. APELA is also an endogenous ligand of APLNR, which is expressed in both lung and cardiac vascularization. Recent studies have shown that exogenous APELA and APLN have therapeutic significance for PAH in a monocrotaline rat model [
43]. Additional studies have shown that APELA triggers vascular relaxation in a NO-independent manner, despite functional similarities between APELA and APLN. Therefore, like APLN, the effect of APELA on the vascular system needs further study [
40]. EIF2AK4 expression leads to pulmonary vascular remodeling, pulmonary telangiectasia, a proliferation of vascular cells, and an increase in typical metabolites in oxidative stress, which are very important for the physiology and pathology of pulmonary hypertension [
44]. HMOX1 is an inducible enzyme that can respond to a variety of stressors, including hypoxia, hyperoxia, acidosis, shear stress, and reactive oxygen species [
45]. Additionally, it plays an important role in resolving pulmonary hypoxia and inflammation [
46]. THBS1 is highly expressed in the plasma and pulmonary vessels of patients with pulmonary hypertension. It inhibits the production of NO through its cognate receptor, CD47 [
47], and also participates in the pathogenesis of hypoxia-induced PAH through TGF β signaling [
48]. TLR4 is expressed in platelets and mediates inflammation and the immune response [
49]. Studies have shown that it plays an important role in maintaining normal pulmonary vasculature and the hypoxia-induced development of PAH [
50,
51]. BCL-2 can reduce cell apoptosis and improve cell activity by releasing cytochrome from mitochondria. Therefore, it is a valuable marker for PAH [
52]. Wnts are secreted glycoproteins that control a variety of biological processes, especially cell proliferation, and trigger intracellular responses through various signaling pathways [
53]. Recent studies have shown that the recruitment of classical and nonclassical Wnt pathways promotes the proliferation, survival, and migration of pulmonary arterial endothelial cells [
54,
55]. CDKs can bind to cyclin D1 to form complex phosphorylated substrates, which are involved in promoting cell cycle progression [
56]. Studies have shown that JQ1+ reduces the expression of CDK2, CDK4, and CDK6 by increasing the expression of CDK inhibitor (CDKN) and CDKN2D to induce G1 cell cycle arrest. JQ1 + also inhibits the migration of serum stimulus pulmonary vascular endothelial cells (human pulmonary microvascular endothelial cell, HPMEC) [
57]. These enriched genes were found to play an important role in broilers with AS and may be a way to explore a new intracellular signaling pathway in the pathogenesis of PAH, which is worthy of further study.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}