

Nipah Virus Disease: Epidemiological, Clinical, Diagnostic and Legislative Aspects of This Unpredictable Emerging Zoonosis
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
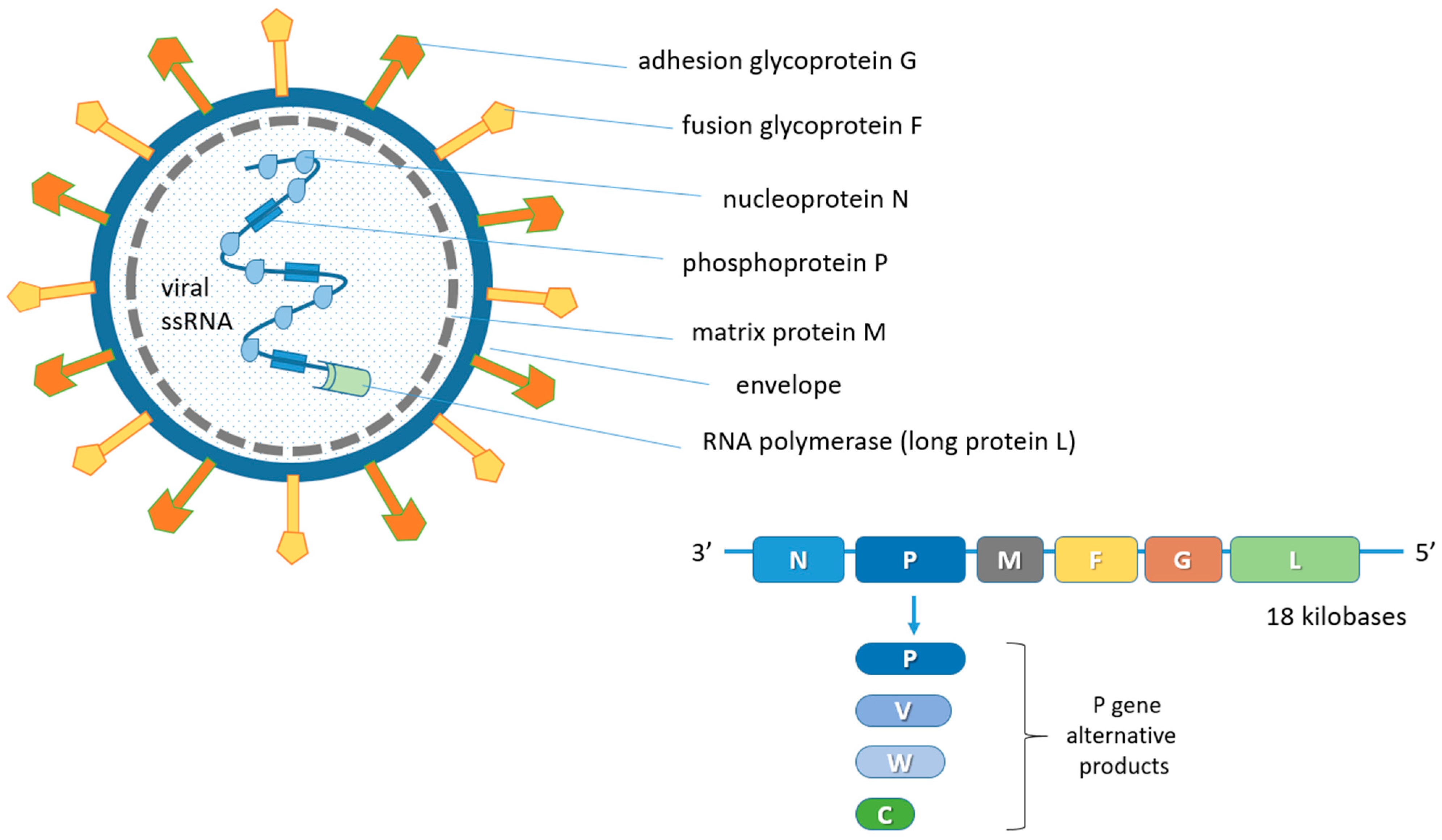
2. Biological Features of the Nipah Virus (NiV)
3. Host Range
4. Epidemiology
5. Biological Reservoir
6. Climate and Anthropogenic Influence on Pteropus Bats and NiV Spillover Events
7. Biological Cycle/Pathogenesis
8. Inflammatory and Immune Response to NiV Infection
9. Clinical Signs
9.1. Chiroptera
9.2. Pigs
9.3. Dogs and Cats
9.4. Humans
10. Differential Diagnosis
10.1. Swine
10.2. Humans
11. Clinical Pathology/Imaging
12. Diagnosis
- Enzyme-linked immunosorbent assay (ELISA): it is used to detect the NiV antigen and also to evaluate the antibody response. It is a simple and inexpensive method for screening suspicious samples [5]. Several techniques of this serological test are used: ELISA-capture uses monoclonal antibodies for the detection of the NiV and to differentiate it from the forms of HeV [203] or even from the one that uses the recombinant N protein of the NiV [204]. Indirect ELISAs for IgG and IgM have also been developed to test both porcine and human sera [2] and are also described to detect seroconversion in bats [205,206]. Another variant of the technique is a sandwich ELISA, which uses rabbit polyclonal antibodies against the NiV G protein [207]. Among the tests used for screening in suids, one based on the use of a recombinant protein N was developed in India (High Security Animal Disease Laboratory [HSADL], Bhopal, India) [5].
- Virus neutralization test (VNT): this test was developed shortly after the Malaysian outbreak and was considered the reference serological test. The conventional NiV VN test commonly uses Vero cells where the prevention of the cytopathic effect via the serum to be tested is considered as a positive neutralization. Furthermore, plate VN tests were developed [208].
- Molecular biology methods: the most sensitive and specific system is PCR. The viral N, M and P sequences are often the targets of RT-PCR and nested-PCR [6]. Furthermore, NiV-targeted PCR has important applications in phylogenetic studies [199,206]. RT-PCR (and its variants) represent the gold standard for the detection of the NiV from various biological samples. RT-PCR for the NiV was developed in 2004 and is based on the N gene sequence. It had high specificity as it was able to detect NiV RNA in blood samples from infected hamsters, where the HeV could not be detected [210]. A commercial kit, developed by Zoologix Inc. (Chatsworth, CA, USA), is able to detect the NiV in whole blood, plasma, CSF and infected tissues/secretions from different animal species [211]. These tests are vital for viral surveillance; for example, the nested RT-PCR duplex, used in Thailand on specimens of P. lyei and which distinguished the NiV variants circulating in bat populations [23,184,192]. Other reported RT-PCR techniques include the SYBR Green quantitative real-time PCR (qRT-PCR) using N-specific primers [212], and a novel one-step qRT-PCR that targets the inter-genic region between the F and G gene sequences for the quantitative detection of replicative NiV RNA, discriminating from the mRNA, which may be more accurate than the conventional qRT-PCR [213].
- Viral isolation: it is very useful in early cases and in new outbreaks where the NiV is suspected. Samples are brain, lung, kidney and/or spleen. Vero cells are a suitable substrate for NiV growth, and the cytopathic effect is usually observed after 3 days of culture in the form of characteristic syncytia and plaques in the cell monolayer [6]. The next step in virus identification includes immunostaining, seroneutralization (SN) and PCR of the culture supernatant. Electron microscopy and immune electro-microscopy are useful tools for identifying the structure of the NiV and detecting virus-antibody interactions, respectively [199,205].
- Immunohistochemistry (IHC): anti-NiV antibodies have been used to stain formalin-fixed tissues of the CNS, lung, spleen, lymph nodes, kidney and heart for detecting viral antigens. In tissue sections, it is possible to identify NiV-associated lesions such as flogosis, necrosis and vasculitis [206].
13. Pathology
13.1. NiV in Natural Host—Pteropus Bats
13.2. Pigs
13.3. Dogs and Cats
13.4. Horses
13.5. NiV in Animal Models
13.6. NiV in Non-Human Primate Models: African Green Monkeys (AGM) and Squirrel Monkeys (SM)
14. Prevention
14.1. Biosecurity
14.2. Vaccines Candidates and Available Vaccines
15. Legislation
15.1. Legislation in Europe
15.2. Legislation in India and Malesia
- a “suspected case” is defined by a person from a community affected by a NiV outbreak showing fever and an altered mental status or respiratory signs;
- a “probable case” is defined by a person who lived in the same area where a confirmed case was detected during an outbreak and died before diagnosis, or a person who came in contact with a confirmed case in the hospital (nosocomial infection) who died before diagnosis during an outbreak;
- a “confirmed case” is defined by a person who was a “suspected case” whose infection was confirmed by diagnostic tests (PCR for viral RNA or virus isolation from biological fluids) [3].
- the state government held a meeting of senior health officials to plan and implement response measures;
- a district central committee was formed and a district action plan for the NiV disease was published for all interested parties;
- a central multidisciplinary team from the NCDC was sent to the state of Kerala to provide technical support. Immediate public health measures were applied, including an active search for cases in families, hospitals, villages and areas with similar topography, especially in the Malappuram district, located in the south-east of the Kozhikode district;
- risk communication campaigns were conducted regarding the transmission of the NiV disease and prevention measures;
- national authorities issued an alert to the districts of Mysuru, Mangaluru, Chamarajanagar and Kodagu in the state of Karnataka, which border the state of Kerala.
- the isolation of potential disease transmitters;
- the blocking of pigs and meat trading (both locally, nationally and internationally);
- restrictions communicated through the media;
- the observation of restrictions strictly controlled by law enforcement and veterinary services;
- the movement of pigs outside the infected areas to state slaughterhouses being allowed as long as there is sanitary authorization from the veterinary services who will escort the animals during the commute, supervising them;
- the establishment of an infected area (2 km) and a protection area (10 km) around the outbreaks. The slaughter of pigs will be carried out with the involvement of the veterinary services, the department of transport, the army and other government agencies and non-governative organizations (NGO);
- the animals will be stunned and killed by bullet and then buried deep in designated areas both on the farm and outside, as appropriate;
- after the killing, cleaning and disinfection operations with chlorine will be carried out on the farm and the sites where the stamping-out is physically performed.
16. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yuen, K.Y.; Fraser, N.S.; Henning, J.; Halpin, K.; Gibson, J.S.; Betzien, L.; Stewart, A.J. Hendra virus: Epidemiology dynamics in relation to climate change, diagnostic tests and control measures. One Health 2021, 12, 100207. [Google Scholar] [CrossRef]
- Mohd Nor, M.N.; Gan, C.H.; Ong, B.L. Nipah virus infection of pigs in peninsular Malaysia. Rev. Sci. Tech. 2000, 19, 160–165. [Google Scholar] [CrossRef]
- Aditi; Shariff, M. Nipah virus infection: A review. Epidemiol. Infect. 2019, 147, e95. [Google Scholar] [CrossRef]
- Thanapongtharm, W.; Linard, C.; Wiriyarat, W.; Chinsorn, P.; Kanchanasaka, B.; Xiao, X.; Biradar, C.; Wallace, R.G.; Gilbert, M. Spatial characterization of colonies of the flying fox bat, a carrier of Nipah virus in Thailand. BMC Vet. Res. 2015, 11, 81. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Dhama, K.; Chakraborty, S.; Tiwari, R.; Natesan, S.; Khandia, R.; Munjal, A.; Vora, K.S.; Latheef, S.K.; Karthik, K.; et al. Nipah virus: Epidemiology, pathology, immunobiology and advances in diagnosis, vaccine designing and control strategies—A comprehensive review. Vet. Q. 2019, 39, 26–55. [Google Scholar] [CrossRef] [Green Version]
- Pillai, V.S.; Krishna, G.; Veettil, M.V. Nipah virus: Past outbreaks and future containment. Viruses 2020, 12, 465. [Google Scholar] [CrossRef] [Green Version]
- Satterfield, B.A.; Dawes, B.E.; Milligan, G.N. Status of vaccine research and development of vaccines for Nipah virus. Vaccine 2016, 34, 2971–2975. [Google Scholar] [CrossRef] [Green Version]
- Devnath, P.; Wajed, S.; Chandra Das, R.; Kar, S.; Islam, I.; Masud, H.M.A.A. The pathogenesis of Nipah virus: A review. Microb. Pathog. 2022, 170, 105693. [Google Scholar] [CrossRef]
- Pedrera, M.; Macchi, F.; McLean, R.K.; Franceschi, V.; Thakur, N.; Russo, L.; Medfai, L.; Todd, S.; Tchilian, E.Z.; Audonnet, J.C.; et al. Bovine herpesvirus-4-vectored delivery of Nipah virus glycoproteins enhances T cell immunogenicity in pigs. Vaccines 2020, 8, 115. [Google Scholar] [CrossRef] [Green Version]
- Mungall, B.A.; Middleton, D.; Crameri, G.; Bingham, J.; Halpin, K.; Russell, G.; Green, D.; McEachern, J.; Pritchard, L.I.; Eaton, B.T.; et al. Feline model of acute Nipah virus infection and protection with a soluble glycoprotein-based subunit vaccine. J. Virol. 2006, 80, 12293–12302. [Google Scholar] [CrossRef]
- Pallister, J.A.; Klein, R.; Arkinstall, R.; Haining, J.; Long, F.; White, J.R.; Payne, J.; Feng, Y.R.; Wang, L.F.; Broder, C.C.; et al. Vaccination of ferrets with a recombinant G glycoprotein subunit vaccine provides protection against Nipah virus disease for over 12 months. Virol. J. 2013, 10, 237. [Google Scholar] [CrossRef] [Green Version]
- Weingartl, H.M.; Berhane, Y.; Caswell, J.L.; Loosmore, S.; Audonnet, J.C.; Roth, J.A.; Czub, M. Recombinant Nipah virus vaccines protect pigs against challenge. J. Virol. 2006, 80, 7929–7938. [Google Scholar] [CrossRef] [Green Version]
- Yoneda, M.; Georges-Courbot, M.C.; Ikeda, F.; Ishii, M.; Nagata, N.; Jacquot, F.; Raoul, H.; Sato, H.; Kai, C. Recombinant measles virus vaccine expressing the Nipah virus glycoprotein protects against lethal Nipah virus challenge. PLoS ONE 2013, 8, e58414. [Google Scholar] [CrossRef]
- Orusa, T.; Orusa, R.; Viani, A.; Carella, E.; Borgogno Mondino, E. Geomatics and EO data to support wildlife diseases assessment at landscape level: A pilot experience to map infectious keratoconjunctivitis in Chamois and phenological trends in Aosta Valley (NW Italy). Remote Sens. 2020, 12, 3542. [Google Scholar] [CrossRef]
- Carella, E.; Orusa, T.; Viani, A.; Meloni, D.; Borgogno Mondino, E.; Orusa, R. An integrated, tentative remote-sensing approach based on NDVI entropy to model canine distemper virus in wildlife and to prompt science-based management policies. Animals 2022, 12, 1049. [Google Scholar] [CrossRef]
- Kumar, V.B.; Rooney, N.; Carr, A. Nipah virus from bats—Another potential pandemic? Risk mapping the impact of anthropogenic and climate change on the transmission of Nipah virus infection to humans. medRxiv 2022. [Google Scholar] [CrossRef]
- Welbergen, J.A.; Klose, S.M.; Markus, N.; Eby, P. Climate change and the effects of temperature extremes on Australian flying-foxes. Proc. Biol. Sci. 2008, 275, 419–425. [Google Scholar] [CrossRef] [Green Version]
- Ratnayake, H.U.; Kearney, M.R.; Govekar, P.; Karoly, D.; Welbergen, J.A. Forecasting wildlife die-offs from extreme heat events. Anim. Conserv. 2019, 22, 386–395. [Google Scholar] [CrossRef]
- Diengdoh, V.L.; Ondei, S.; Hunt, M.; Brook, B.W. Predicted impacts of climate change and extreme temperature events on the future distribution of fruit bat species in Australia. Glob. Ecol. Conserv. 2022, 37, e02181. [Google Scholar] [CrossRef]
- European Union (EU). Regulation (EU) 2016/429 of the European Parliament and of the Council of 9 March 2016 on Transmissible Animal Diseases and Amending and Repealing Certain Acts in the Area of Animal Health (‘Animal Health Law’). Off. J. 2016. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32016R0429&from=EN (accessed on 22 December 2022).
- Aziz, B.J.; Azri, B.A. Nipah virus infection—Malaysia experience. In Proceedings of the World Organization of Animal Health (OIE) Conference WILDLIFE ACTES 2011; Available online: https://www.woah.org/fileadmin/Home/eng/Conferences_Events/sites/WILDLIFE_ACTES_2011/Presentations/S1_3_AzriBinAdzhar.pdf (accessed on 22 December 2022).
- National Centre for Disease Control (NCDC) India. Available online: https://ncdc.gov.in/ (accessed on 22 December 2022).
- Chua, K.B.; Bellini, W.J.; Rota, P.A.; Harcourt, B.H.; Tamin, A.; Lam, S.K.; Ksiazek, T.G.; Rollin, P.E.; Zaki, S.R.; Goldsmith, C.S. Nipah virus: A recently emergent deadly paramyxovirus. Science 2000, 288, 1432–1435. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Shen, Q.T. Insights into paramyxovirus nucleocapsids from diverse assemblies. Viruses 2021, 13, 2479. [Google Scholar] [CrossRef]
- Middleton, D. Hendra virus. Vet. Clin. N. Am. Equine Pract. 2014, 30, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Middleton, D.; Pallister, J.; Klein, R.; Feng, Y.R.; Haining, J.; Arkinstall, R.; Frazer, L.; Huang, J.A.; Edwards, N.; Wareing, M.; et al. Hendra virus vaccine, a one health approach to protecting horse, human, and environmental health. Emerg. Infect. Dis. 2014, 20, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Enchèry, F.; Horvat, B. Understanding the interaction between Henipaviruses and their natural host, fruit bats: Paving the way toward control of highly lethal infection in humans. Int. Rev. Immunol. 2017, 36, 108–121. [Google Scholar] [CrossRef] [PubMed]
- Eaton, B.T.; Broder, C.C.; Middleton, D.; Wang, L.F. Hendra and Nipah viruses: Different and dangerous. Nat. Rev. Microbiol. 2006, 4, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Iehlé, C.; Razafitrimo, G.; Razainirina, J.; Andriaholinirina, N.; Goodman, S.M.; Faure, C.; Georges-Courbot, M.C.; Rousset, D.; Reynes, J.M. Henipavirus and Tioman virus antibodies in pteropodid bats, Madagascar. Emerg. Infect. Dis. 2007, 13, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Hayman, D.T.S.; Suu-Ire, R.; Breed, A.C.; McEachern, J.A.; Wang, L.; Wood, J.L.N.; Cunningham, A.A. Evidence of Henipavirus infection in West African fruit bats. PLoS ONE 2008, 3, e2739. [Google Scholar] [CrossRef] [Green Version]
- Drexler, J.F.; Corman, V.M.; Gloza-Rausch, F.; Seebens, A.; Annan, A.; Ipsen, A.; Kruppa, T.; Müller, M.A.; Kalko, E.K.V.; Adu-Sarkodie, Y.; et al. Henipavirus RNA in African bats. PLoS ONE 2009, 4, e6367. [Google Scholar] [CrossRef] [Green Version]
- Hayman, D.T.S.; Wang, L.F.; Barr, J.; Baker, K.S.; Suu-Ire, R.; Broder, C.C.; Cunningham, A.A.; Wood, J.L.N. Antibodies to Henipavirus or henipa-like viruses in domestic pigs in Ghana, West Africa. PLoS ONE 2011, 6, e25256. [Google Scholar] [CrossRef] [Green Version]
- Drexler, J.F.; Corman, V.M.; Müller, M.A.; Maganga, G.D.; Vallo, P.; Binger, T.; Gloza-Rausch, F.; Cottontail, V.M.; Rasche, A.; Yordanov, S.; et al. Bats host major mammalian paramyxoviruses. Nat. Commun. 2012, 3, 796. [Google Scholar] [CrossRef]
- Mbu’u, C.M.; Mbacham, W.F.; Gontao, P.; Sado Kamdem, S.L.; Nlôga, A.M.N.; Groschup, M.H.; Wade, A.; Fischer, K.; Balkema-Buschmann, A. Henipaviruses at the interface between bats, livestock and human population in Africa. Vector-Borne Zoonotic Dis. 2019, 19, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Pernet, O.; Schneider, B.S.; Beaty, S.M.; LeBreton, M.; Yun, T.E.; Park, A.; Zachariah, T.T.; Bowden, T.A.; Hitchens, P.; Ramirez, C.M.; et al. Evidence for Henipavirus spillover into human populations in Africa. Nat. Commun. 2014, 5, 5342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Dimitrov, A.S.; Bossart, K.N.; Crameri, G.; Bishop, K.A.; Choudhry, V.; Mungall, B.A.; Feng, Y.R.; Choudhary, A.; Zhang, M.Y.; et al. Potent neutralization of Hendra and Nipah viruses by human monoclonal antibodies. J. Virol. 2006, 80, 891–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellini, W.J.; Harcourt, B.H.; Bowden, N.; Rota, P.A. Nipah virus: An emergent paramyxovirus causing severe encephalitis in humans. J. Neurovirol. 2005, 11, 481–487. [Google Scholar] [CrossRef]
- Lamb, R.A.; Parks, G.D. Paramyxoviridae: The viruses and their replication. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott, Williams and Wilkins: Philadelphia, PA, USA, 2006; pp. 1449–1496. [Google Scholar]
- El Najjar, F.; Schmitt, A.P.; Dutch, R.E. Paramyxovirus glycoprotein incorporation, assembly and budding: A three way dance for infectious particle production. Viruses 2014, 6, 3019–3054. [Google Scholar] [CrossRef] [Green Version]
- Maisner, A.; Neufeld, J.; Weingartl, H. Organ- and endotheliotropism of Nipah virus infections in vivo and in vitro. Thromb. Haemost. 2009, 102, 1014–1023. [Google Scholar] [CrossRef]
- Aguilar, H.C.; Henderson, B.A.; Zamora, J.L.; Johnston, G.P. Paramyxovirus glycoproteins and the membrane fusion process. Curr. Clin. Microbiol. Rep. 2016, 3, 142–154. [Google Scholar] [CrossRef] [Green Version]
- Walpita, P.; Cong, Y.; Jahrling, P.B.; Rojas, O.; Postnikova, E.; Yu, S.; Johns, L.; Holbrook, M.R. A VLP-based vaccine provides complete protection against Nipah virus challenge following multiple-dose or single-dose vaccination schedules in a hamster model. NPJ Vaccines 2017, 2, 21. [Google Scholar] [CrossRef]
- Wang, L.; Harcourt, B.H.; Yu, M.; Tamin, A.; Rota, P.A.; Bellini, W.J.; Eaton, B.T. Molecular biology of Hendra and Nipah viruses. Microbes Infect. 2001, 3, 279–287. [Google Scholar] [CrossRef]
- Harcourt, B.H.; Tamin, A.; Ksiazek, T.G.; Rollin, P.E.; Anderson, L.J.; Bellini, W.J.; Rota, P.A. Molecular characterization of Nipah virus, a newly emergent paramyxovirus. Virology 2000, 271, 334–349. [Google Scholar] [CrossRef]
- Harcourt, B.H.; Lowe, L.; Tamin, A.; Liu, X.; Bankamp, B.; Bowden, N.; Rollin, P.E.; Comer, J.A.; Ksiazek, T.G.; Hossain, M.J.; et al. Genetic characterization of Nipah virus, Bangladesh, 2004. Emerg. Infect. Dis. 2005, 11, 1594–1597. [Google Scholar] [CrossRef]
- de Wit, E.; Munster, V.J. Nipah virus emergence, transmission, and pathogenesis. In Global Virology I—Identifying and Investigating Viral Diseases; Shapshak, P., Sinnott, J., Somboonwit, C., Kuhn, J., Eds.; Springer: New York, NY, USA, 2015; pp. 125–146. [Google Scholar]
- de Wit, E.; Munster, V.J. Animal models of disease shed light on Nipah virus pathogenesis and transmission. J. Pathol. 2015, 235, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Angeletti, S.; Presti, A.L.; Cella, E.; Ciccozzi, M. Molecular epidemiology and phylogeny of Nipah virus infection: A mini review. Asian Pac. J. Trop. Med. 2016, 9, 630–634. [Google Scholar] [CrossRef] [Green Version]
- AbuBakar, S.; Chang, L.Y.; Ali, A.R.; Sharifah, S.H.; Yusoff, K.; Zamrod, Z. Isolation and molecular identification of Nipah virus from pigs. Emerg. Infect. Dis. 2004, 10, 2228–2230. [Google Scholar] [CrossRef] [PubMed]
- Liew, Y.J.M.; Ibrahim, P.A.S.; Ong, H.M.; Chong, C.N.; Tan, C.T.; Schee, J.P.; Gómez Román, R.; Cherian, N.G.; Wong, W.F.; Chang, L.Y. The immunobiology of Nipah virus. Microorganisms 2022, 10, 1162. [Google Scholar] [CrossRef]
- Tan, C.T.; Chua, K.B. Nipah virus encephalitis. Curr. Infect. Dis. Rep. 2008, 10, 315–320. [Google Scholar] [CrossRef]
- Vidal, S.; Curran, J.; Kolakofsky, D. Editing of the Sendai virus P/C mRNA by G insertion occurs during mRNA synthesis via a virus-encoded activity. J. Virol. 1990, 64, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Steward, M.; Vipond, I.B.; Millar, N.S.; Emmerson, P.T. RNA editing in Newcastle disease virus. J. Gen. Virol. 1993, 74, 2539–2547. [Google Scholar] [CrossRef]
- Delenda, C.; Taylor, G.; Hausmann, S.; Garcin, D.; Kolakofsky, D. Sendai viruses with altered P, V, and W protein expression. Virology 1998, 242, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Hansson, E.; Langedijk, J.P.; Eaton, B.T.; Wang, L.F. The attachment protein of Hendra virus has high structural similarity but limited primary sequence homology compared with viruses in the genus Paramyxovirus. Virology 1998, 251, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Reynes, J.M.; Counor, D.; Ong, S.; Faure, C.; Seng, V.; Molia, S.; Walston, J.; Georges-Courbot, M.C.; Deubel, V.; Sarthou, J.L. Nipah virus in Lyle’s flying foxes, Cambodia. Emerg. Infect. Dis. 2005, 11, 1042–1047. [Google Scholar] [CrossRef] [PubMed]
- Chadha, M.S.; Comer, J.A.; Lowe, L.; Rota, P.A.; Rollin, P.E.; Bellini, W.J.; Ksiazek, T.G.; Mishra, A. Nipah virus-associated encephalitis outbreak, Siliguri, India. Emerg. Infect. Dis. 2006, 12, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Bonaparte, M.I.; Dimitrov, A.S.; Bossart, K.N.; Crameri, G.; Mungall, B.A.; Bishop, K.A.; Choudhry, V.; Dimitrov, D.S.; Wang, L.F.; Eaton, B.T. Ephrin-B2 ligand is a functional receptor for Hendra virus and Nipah virus. Proc. Natl. Acad. Sci. USA 2005, 102, 10652–10657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negrete, O.A.; Levroney, E.L.; Aguilar, H.C.; Bertolotti-Ciarlet, A.; Nazarian, R.; Tajyar, S.; Lee, B. EphrinB2 is the entry receptor for Nipah virus, an emergent deadly paramyxovirus. Nature 2005, 436, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Negrete, O.A.; Wolf, M.C.; Aguilar, H.C.; Enterlein, S.; Wang, W.; Mühlberger, E.; Su, S.V.; Bertolotti-Ciarlet, A.; Flick, R.; Lee, B. Two key residues in ephrinB3 are critical for its use as an alternative receptor for Nipah virus. PLoS Pathog. 2006, 2, e7. [Google Scholar] [CrossRef] [PubMed]
- Liebl, D.J.; Morris, C.J.; Henkemeyer, M.; Parada, L.F. mRNA expression of ephrins and Eph receptor tyrosine kinases in the neonatal and adult mouse central nervous system. J. Neurosci. Res. 2003, 71, 7–22. [Google Scholar] [CrossRef]
- Zimmer, M.; Palmer, A.; Köhler, J.; Klein, R. Ephb-ephrinB bi-directional endocytosis terminates adhesion allowing contact-mediated repulsion. Nat. Cell Biol. 2003, 5, 869–878. [Google Scholar] [CrossRef]
- Bossart, K.N.; Tachedjian, M.; McEachern, J.A.; Crameri, G.; Zhu, Z.; Dimitrov, D.S.; Broder, C.C.; Wang, L.F. Functional studies of host-specific ephrin-B ligands as Henipavirus receptors. Virology 2008, 372, 357–371. [Google Scholar] [CrossRef] [Green Version]
- Hassan, M.Z.; Sazzad, H.M.S.; Luby, S.P.; Sturm-Ramirez, K.; Bhuiyan, M.U.; Rahman, M.Z.; Islam, M.M.; Ströher, U.; Sultana, S.; Kafi, M.A.H.; et al. Nipah virus contamination of hospital surfaces during outbreaks, Bangladesh, 2013–2014. Emerg. Infect. Dis. 2018, 24, 15–21. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Geographic Distribution of Henipavirus Outbreaks and Fruit Bats of Pteropodidae Family. Reproduced According to WHO Permission ID: 390902 (25 October 2022). Available online: https://www.who.int/health-topics/nipah-virus-infection#tab=tab_1 (accessed on 22 December 2022).
- Nikolay, B.; Salje, H.; Hossain, M.J.; Khan, A.K.M.D.; Sazzad, H.M.S.; Rahman, M.; Daszak, P.; Ströher, U.; Pulliam, J.R.C.; Kilpatrick, A.M.; et al. Transmission of Nipah virus—14 Years of investigations in Bangladesh. N. Engl. J. Med. 2019, 380, 1804–1814. [Google Scholar] [CrossRef] [PubMed]
- Epstein, J.H.; Rahman, S.A.; Zambriski, A.; Halpin, K.; Meehan, G.; Jamaluddin, A.A.; Hassan, S.S.; Field, H.E.; Hyatt, A.D.; Daszak, P. Henipavirus Ecology Research Group. Feral cats and risk for Nipah virus transmission, Emerg. Infect. Dis. 2006, 12, 1178–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Organization for Animal Health (OIE). Nipah Virus Infection (Wild Animals). Available online: https://www.woah.org/en/disease/nipah-virus-infection-wild-animals/ (accessed on 22 December 2022).
- World Organization for Animal Health (OIE). Henipaviruses (Nipah viruses) (Infection with). Available online: https://www.woah.org/app/uploads/2022/02/henipaviruses-nipah-viruses-infection-with.pdf (accessed on 22 December 2022).
- Eshaghi, M.; Tan, W.S.; Mohodin, T.B.; Yusoff, K. Nipah virus glycoprotein: Production in baculovirus and application in diagnosis. Virus Res. 2004, 106, 71–76. [Google Scholar] [CrossRef]
- Weingartl, H.M. Hendra and Nipah viruses: Pathogenesis, animal models and recent breakthroughs in vaccination. Vaccine 2015, 5, 59–74. [Google Scholar] [CrossRef] [Green Version]
- Ching, P.K.G.; de los Reyes, V.C.; Sucaldito, M.N.; Tayag, E.; Columna-Vingno, A.B.; Malbas, F.F., Jr.; Bolo, G.C., Jr.; Sejvar, J.J.; Eagles, D.; Playford, G.; et al. Outbreak of Henipavirus infection, Philippines, 2014. Emerg. Infect. Dis. 2015, 21, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Looi, L.M.; Chua, K.B. Lessons from the Nipah virus outbreak in Malaysia. Malays. J. Pathol. 2007, 29, 63–67. [Google Scholar] [PubMed]
- Paton, N.I.; Leo, Y.S.; Zaki, S.R.; Auchus, A.P.; Lee, K.E.; Ling, A.E.; Chew, S.K.; Ang, B.; Rollin, P.E.; Umapathi, T.; et al. Outbreak of Nipah-virus infection among abattoir workers in Singapore. Lancet 1999, 354, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Parashar, U.D.; Sunn, L.M.; Ong, F.; Mounts, A.W.; Arif, M.T.; Ksiazek, T.G.; Kamaluddin, M.A.; Mustafa, A.N.; Kaur, H.; Ding, L.M.; et al. Case-control study of risk factors for human infection with a new zoonotic Paramyxovirus, Nipah virus, during a 1998–1999 outbreak of severe encephalitis in Malaysia. J. Infect. Dis. 2000, 181, 1755–1759. [Google Scholar] [CrossRef]
- Institute of Epidemiology, Disease Control and Research (IEDCR). Available online: http://www.iedcr.org/ (accessed on 22 December 2022).
- Yadav, P.D.; Raut, C.G.; Shete, A.M.; Mishra, A.C.; Towner, J.S.; Nichol, S.T.; Mourya, D.T. Detection of Nipah virus RNA in fruit Bat (Pteropus giganteus) from India. Am. J. Trop. Med. Hyg. 2012, 87, 576–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, S.; Khan, S.U.; Crameri, G.; Epstein, J.H.; Broder, C.C.; Islam, A.; Peel, A.J.; Barr, J.; Daszak, P.; Wang, L.F.; et al. Serological evidence of Henipavirus exposure in cattle, goats, and pigs in Bangladesh. PLoS Negl. Trop. Dis. 2014, 8, e3302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luby, S.P.; Rahman, M.; Hossain, M.J.; Blum, L.S.; Husain, M.M.; Gurley, E.; Khan, R.; Ahmed, B.N.; Rahman, S.; Nahar, N.; et al. Foodborne transmission of Nipah virus, Bangladesh. Emerg. Infect. Dis. 2006, 12, 1888–1894. [Google Scholar] [CrossRef]
- Khan, M.S.U.; Hossain, J.; Gurley, E.S.; Nahar, N.; Sultana, R.; Luby, S.P. Use of infrared camera to understand bats’ access to date palm Sap: Implications for preventing Nipah virus transmission. EcoHealth 2010, 7, 517–525. [Google Scholar] [CrossRef]
- Gurley, E.S.; Montgomery, J.M.; Hossain, M.J.; Bell, M.; Azad, A.K.; Islam, M.R.; Molla, M.A.; Carroll, D.S.; Ksiazek, T.G.; Rota, P.A.; et al. Person-to-person transmission of Nipah virus in a Bangladeshi community. Emerg. Infect. Dis. 2007, 13, 1031–1037. [Google Scholar] [CrossRef]
- Homaira, N.; Rahman, M.; Hossain, M.J.; Epstein, J.H.; Sultana, R.; Khan, M.S.; Podder, G.; Nahar, K.; Ahmed, B.; Gurley, E.S.; et al. Nipah virus outbreak with person-to-person transmission in a district of Bangladesh, 2007. Epidemiol. Infect. 2010, 138, 1630–1636. [Google Scholar] [CrossRef] [Green Version]
- Rimi, N.A.; Sultana, R.; Luby, S.P.; Islam, M.S.; Uddin, M.; Hossain, M.J.; Zaman, R.U.; Nahar, N.; Gurley, E.S. Infrastructure and contamination of the physical environment in three Bangladeshi hospitals: Putting infection control into context. PLoS ONE 2014, 9, e89085. [Google Scholar] [CrossRef]
- Luby, S.P.; Hossain, M.J.; Gurley, E.S.; Ahmed, B.N.; Banu, S.; Khan, S.U.; Homaira, N.; Rota, P.A.; Rollin, P.E.; Comer, J.A.; et al. Recurrent zoonotic transmission of Nipah virus into humans, Bangladesh, 2001–2007. Emerg. Infect. Dis. 2009, 15, 1229–1235. [Google Scholar] [CrossRef]
- Luby, S.P.; Gurley, E.S.; Hossain, M.J. Transmission of human infection with Nipah virus. Clin. Infect. Dis. 2009, 49, 1743–1748. [Google Scholar] [CrossRef] [Green Version]
- Ang, B.S.P.; Lim, T.C.C.; Wang, L. Nipah Virus Infection. J. Clin. Microbiol. 2018, 56, e01875-17. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Gupta, N.; Kodan, P.; Mittal, A.; Ray, Y.; Nischal, N.; Soneja, M.; Biswas, A.; Wig, N. Nipah virus disease: A rare and intractable disease. Intractable Rare Dis. Res. 2019, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Sejvar, J.J.; Hossain, J.; Saha, S.K.; Gurley, E.S.; Banu, S.; Hamadani, J.D.; Faiz, M.A.; Siddiqui, F.M.; Mohammad, Q.D.; Mollah, A.H.; et al. Long-term neurological and functional outcome in Nipah virus infection. Ann. Neurol. 2007, 62, 235–242. [Google Scholar] [CrossRef]
- Goh, K.J.; Tan, C.T.; Chew, N.K.; Tan, P.S.; Kamarulzaman, A.; Sarji, S.A.; Wong, K.T.; Abdullah, B.J.; Chua, K.B.; Lam, S.K. Clinical features of Nipah virus encephalitis among pig farmers in Malaysia. N. Engl. J. Med. 2000, 342, 1229–1235. [Google Scholar] [CrossRef]
- Arankalle, V.A.; Bandyopadhyay, B.T.; Ramdasi, A.Y.; Jadi, R.; Patil, D.R.; Rahman, M.; Majumdar, M.; Banerjee, P.S.; Hati, A.K.; Goswami, R.P.; et al. Genomic characterization of Nipah virus, West Bengal, India. Emerg. Infect. Dis. 2011, 17, 907–909. [Google Scholar] [CrossRef]
- Singhai, M.; Jain, R.; Jain, S.; Bala, M.; Singh, S.; Goyal, R. Nipah virus disease: Recent perspective and one health approach. Ann. Glob. Health 2021, 87, 102. [Google Scholar] [CrossRef]
- Alam, A.M. Nipah virus, an emerging zoonotic disease causing fatal encephalitis. Clin. Med. 2022, 22, 348–352. [Google Scholar] [CrossRef]
- Thakur, N.; Bailey, D. Advances in diagnostics, vaccines and therapeutics for Nipah virus. Microbes Infect. 2019, 21, 278–286. [Google Scholar] [CrossRef]
- Sazzad, H.M.S.; Hossain, M.J.; Gurley, E.S.; Ameen, K.M.H.; Parveen, S.; Islam, M.S.; Faruque, L.I.; Podder, G.; Banu, S.S.; Lo, M.K.; et al. Nipah virus infection outbreak with nosocomial and corpse-to-human transmission, Bangladesh. Emerg. Infect. Dis. 2013, 19, 210–217. [Google Scholar] [CrossRef]
- Clayton, B.A. Nipah virus: Transmission of a zoonotic paramyxovirus. Curr. Opin. Virol. 2017, 22, 97–104. [Google Scholar] [CrossRef]
- Clayton, B.A.; Middleton, D.; Bergfeld, J.; Haining, J.; Arkinstall, R.; Wang, L.; Marsh, G.A. Transmission routes for Nipah virus from Malaysia and Bangladesh. Emerg. Infect. Dis. 2012, 18, 1983–1993. [Google Scholar] [CrossRef]
- Sayed, A.; Bottu, A.; Qaisar, M.; Mane, M.P.; Acharya, Y. Nipah virus: A narrative review of viral characteristics and epidemiological determinants. Public Health 2019, 173, 97–104. [Google Scholar] [CrossRef]
- Openshaw, J.J.; Hegde, S.; Sazzad, H.M.S.; Khan, S.U.; Hossain, M.J.; Epstein, J.H.; Daszak, P.; Gurley, E.S.; Luby, S.P. Bat hunting and bat-human interactions in Bangladeshi villages: Implications for zoonotic disease transmission and bat conservation. Transbound. Emerg. Dis. 2017, 64, 1287–1293. [Google Scholar] [CrossRef]
- Ray, A.; Mittal, A. Nipah virus infection: Gaps in evidence and its public health importance. Public Health 2020, 181, 202–203. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, G.; Abdulmajeed, J.; Santhosha, D.; Aswathyraj, S.; Robin, S.; Jayaram, A.; Radhakrishnan, C.; Sajeeth, K.K.G.; Sakeena, K.; Jayasree, V.; et al. Persistence of Nipah virus RNA in semen of survivor. Clin. Infect. Dis. 2019, 69, 377–378. [Google Scholar] [CrossRef] [PubMed]
- Yob, J.M.; Field, H.; Rashdi, A.M.; Morrissy, C.; van der Heide, B.; Rota, P.; bin Adzhar, A.; White, J.; Daniels, P.; Jamaluddin, A.; et al. Nipah virus infection in bats (order Chiroptera) in Peninsular Malaysia. Emerg. Infect. Dis. 2001, 7, 439–441. [Google Scholar] [CrossRef] [PubMed]
- Chua, K.B.; Koh, C.L.; Hooi, P.S.; Wee, K.F.; Khong, J.H.; Chua, B.H.; Chan, Y.P.; Lim, M.E.; Lam, S.K. Isolation of Nipah virus from Malaysian flying-foxes. Microbes Infect. 2002, 4, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Hsu, V.P.; Hossain, M.J.; Parashar, U.D.; Ali, M.M.; Ksiazek, T.G.; Kuzmin, I.; Niezgoda, M.; Rupprecht, C.; Bresee, J.; Breiman, R.F. Nipah virus encephalitis reemergence, Bangladesh. Emerg. Infect. Dis. 2004, 10, 2082–2087. [Google Scholar] [CrossRef]
- Epstein, J.H.; Anthony, S.J.; Islam, A.; Kilpatrick, A.M.; Ali Khan, S.; Balkey, M.D.; Ross, N.; Smith, I.; Zambrana-Torrelio, C.; Tao, Y.; et al. Nipah virus dynamics in bats and implications for spillover to humans. Proc. Natl. Acad. Sci. USA 2020, 117, 29190–29201. [Google Scholar] [CrossRef]
- Geller-Grimm, F. Pteropus giganteus. Zoological Garden Berlin, Germany (Personal Work, 2002). This File (Photograph) is Licensed under the Creative Commons Attribution-Share Alike 2.5 Generic License. Available online: https://creativecommons.org/licenses/by-sa/2.5/deed.en (accessed on 22 December 2022).
- Nowak, R.M. Walker’s Mammals of the World, 6th ed.; Johns Hopkins University Press: Baltimore, MD, USA, 1999; pp. 264–271. [Google Scholar]
- Sendow, I.; Field, H.E.; Adjid, A.; Ratnawati, A.; Breed, A.C.; Darminto Morrissy, C.; Daniels, P. Screening for Nipah virus infection in West Kalimantan province, Indonesia. Zoonoses Public Health 2010, 57, 499–503. [Google Scholar] [CrossRef]
- Simmons, N.B. Genus Pteropus. In Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; Wilson, D.E., Reeder, D.M., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2005; pp. 334–346. [Google Scholar]
- Marshall, A.G. Old World phytophagous bats (Megachiroptera) and their food plants: A survey. Zool. J. Linn. Soc. 1985, 83, 351–369. [Google Scholar] [CrossRef]
- Aziz, S.A.; Clements, G.R.; Peng, L.Y.; Campos-Arceiz, A.; McConkey, K.R.; Forget, P.M.; Gan, H.M. Elucidating the diet of the island flying fox (Pteropus hypomelanus) in Peninsular Malaysia through Illumina Next-Generation Sequencing. PeerJ 2017, 5, e3176. [Google Scholar] [CrossRef] [Green Version]
- Clulow, S.; Blundell, A.T. Deliberate insectivory by the fruit bat Pteropus poliocephalus by aerial hunting. Acta Chiropt. 2011, 13, 201–205. [Google Scholar] [CrossRef]
- New South Wales Wildlife Council Inc. (NWC)—Nature Conservation Council (NCC) of NSW. Appendix 1. Flying-Foxes in NSW–Brief Summary. In Proceedings of the Annual Conference 2010, Surry Hills, Australia, 30–31 October 2010; Available online: https://assets.nationbuilder.com/natureorg/legacy_url/2532/2010-annual-conference-minutes.pdf?1630462707 (accessed on 22 December 2022).
- Aziz, S.A.; Olival, K.J.; Bumrungsri, S.; Richards, G.C.; Racey, P.A. The conflict between pteropodid bats and fruit growers: Species, legislation and mitigation. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C., Kingston, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Cappelle, J.; Hoem, T.; Hul, V.; Furey, N.; Nguon, K.; Prigent, S.; Dupon, L.; Ken, S.; Neung, C.; Hok, V.; et al. Nipah virus circulation at human-bat interfaces, Cambodia. Bull. World Health Organ. 2020, 98, 539–547. [Google Scholar] [CrossRef]
- Luis, A.D.; Hayman, D.T.S.; O’Shea, T.J.; Cryan, P.M.; Gilbert, A.T.; Pulliam, J.R.C.; Mills, J.N.; Timonin, M.E.; Willis, C.K.; Cunningham, A.A.; et al. A comparison of bats and rodents as reservoirs of zoonotic viruses: Are bats special? Proc. Biol. Sci. 2013, 280, 20122753. [Google Scholar] [CrossRef] [Green Version]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important reservoir hosts of emerging viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef] [Green Version]
- Hengjan, Y.; Pramono, D.; Takemae, H.; Kobayashi, R.; Iida, K.; Ando, T.; Kasmono, S.; Basri, C.; Fitriana, Y.S.; Arifin, E.M.Z.; et al. Daytime behavior of Pteropus vampyrus in a natural habitat: The driver of viral transmission. J. Vet. Med. Sci. 2017, 79, 1125–1133. [Google Scholar] [CrossRef] [Green Version]
- Yong, M.Y.; Lee, S.C.; Ngui, R.; Lim, Y.A.; Phipps, M.E.; Chang, L.Y. Seroprevalence of Nipah virus infection in peninsular Malaysia. J. Infect. Dis. 2020, 221, S370–S374. [Google Scholar] [CrossRef]
- Bates, P.J.J.; Harrison, D. Bats of the Indian Subcontinent; Harrison Zoological Museum: Sevenoaks, UK, 1997. [Google Scholar]
- Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) (2022). The CITES Appendices. Available online: https://cites.org/eng/app/index.php (accessed on 22 December 2022).
- Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES), United Nations Environment Programme (UNEP) (2022). Appendices I, II and III. Available online: https://cites.org/sites/default/files/eng/app/2022/E-Appendices-2022-06-22.pdf (accessed on 22 December 2022).
- Anyamba, A.; Chretien, J.P.; Britch, S.C.; Soebiyanto, R.P.; Small, J.L.; Jepsen, R.; Forshey, B.M.; Sanchez, J.L.; Smith, R.D.; Harris, R.; et al. Global disease outbreaks associated with the 2015-2016 El Niño event. Sci. Rep. 2019, 9, 1930. [Google Scholar] [CrossRef] [Green Version]
- Orusa, T.; Borgogno Mondino, E. Exploring short-term climate change effects on rangelands and broad-leaved forests by free satellite data in Aosta Valley (Northwest Italy). Climate 2021, 9, 47. [Google Scholar] [CrossRef]
- Carlson, C.J.; Albery, G.F.; Merow, C.; Trisos, C.H.; Zipfel, C.M.; Eskew, E.A.; Olival, K.J.; Ross, N.; Bansal, S. Climate change increases cross-species viral transmission risk. Nature 2022, 607, 555–562. [Google Scholar] [CrossRef]
- Translated by Volken, E.; Brönnimann, S. 1884. Köppen, W. “Die Wärmezonen der Erde, nach der Dauer der heissen, gemässigten und kalten Zeit und nach der Wirkung der Wärme auf die organische Welt betrachtet” [The thermal zones of the earth according to the duration of hot, moderate and cold periods and to the impact of heat on the organic world)]. Meteorol. Z. 2011, 20, 351–360. [Google Scholar] [CrossRef]
- Mickleburgh, S.P.; Hutson, A.M.; Racey, P.A. Old World Fruit Bats—An Action Plan for Their Conservation; International Union for Conservation of Nature and Natural Resources (IUCN): Gland, Switzerland, 1992; ISBN 2-8317-0055-8. Available online: https://portals.iucn.org/library/efiles/documents/1992-034.pdf (accessed on 22 December 2022).
- Palmer, C.; Price, O.F.; Bach, C. Foraging ecology of the black flying fox (Pteropus alecto) in the seasonal tropics of the Northern Territory, Australia. Wildl. Res. 2000, 27, 169–178. [Google Scholar] [CrossRef]
- Courts, S. Dietary strategies of old world fruit bats (Megachiroptera, Pteropodidae): How do they obtain sufficient protein? Mammal Rev. 1998, 28, 185–193. [Google Scholar] [CrossRef]
- Giles, J.R.; Eby, P.; Parry, H.; Peel, A.J.; Plowright, R.K.; Westcott, D.A.; McCallum, H. Environmental drivers of spatiotemporal foraging intensity in fruit bats and implications for Hendra virus ecology. Sci. Rep. 2018, 8, 9555. [Google Scholar] [CrossRef] [PubMed]
- Edson, D.; Field, H.; McMichael, L.; Jordan, D.; Kung, N.; Mayer, D.; Smith, C. Flying-fox roost disturbance and Hendra virus spillover risk. PLoS ONE 2015, 10, e0125881. [Google Scholar] [CrossRef] [PubMed]
- McMichael, L.; Edson, D.; Smith, C.; Mayer, D.; Smith, I.; Kopp, S.; Meers, J.; Field, H. Physiological stress and Hendra virus in flying-foxes (Pteropus spp.), Australia. PLoS ONE 2017, 12, e0182171. [Google Scholar] [CrossRef] [Green Version]
- Edson, D.; Peel, A.J.; Huth, L.; Mayer, D.G.; Vidgen, M.E.; McMichael, L.; Broos, A.; Melville, D.; Kristoffersen, J.; de Jong, C.; et al. Time of year, age class and body condition predict Hendra virus infection in Australian black flying foxes (Pteropus alecto). Epidemiol. Infec. 2019, 147, e240. [Google Scholar] [CrossRef] [Green Version]
- Plowright, R.K.; Field, H.E.; Smith, C.; Divljan, A.; Palmer, C.; Tabor, G.; Daszak, P.; Foley, J.E. Reproduction and nutritional stress are risk factors for Hendra virus infection in little red flying foxes (Pteropus scapulatus). Proc. Biol. Sci. 2008, 275, 861–869. [Google Scholar] [CrossRef] [Green Version]
- McFarlane, R.; Becker, N.; Field, H. Investigation of the climatic and environmental context of Hendra virus spillover events 1994–2010. PLoS ONE 2011, 6, e28374. [Google Scholar] [CrossRef] [Green Version]
- Páez, D.J.; Giles, J.; McCallum, H.; Field, H.; Jordan, D.; Peel, A.J.; Plowright, R.K. Conditions affecting the timing and magnitude of Hendra virus shedding across pteropodid bat populations in Australia. Epidemiol. Infec. 2017, 145, 3143–3153. [Google Scholar] [CrossRef] [Green Version]
- Martin, G.; Yanez-Arenas, C.; Chen, C.; Plowright, R.K.; Webb, R.J.; Skerratt, L.F. Climate change could increase the geographic extent of Hendra virus spillover risk. EcoHealth 2018, 15, 509–525. [Google Scholar] [CrossRef] [Green Version]
- Martin, G.; Yanez-Arenas, C.; Plowright, R.K.; Chen, C.; Roberts, B.; Skerratt, L.F. Hendra virus spillover is a bimodal system driven by climatic factors. EcoHealth 2018, 15, 526–542. [Google Scholar] [CrossRef]
- Timmermann, A.; Oberhuber, J.; Bacher, A.; Esch, M.; Latif, M.; Roeckner, E. Increased El Niño frequency in a climate model forced by future greenhouse warming. Nature 1999, 398, 694–697. [Google Scholar] [CrossRef]
- Mpelasoka, F.; Hennessy, K.; Jones, R.; Bates, B. Comparison of suitable drought indices for climate change impacts assessment over Australia towards resource management. Int. J. Climatol. 2008, 28, 1283–1292. [Google Scholar] [CrossRef]
- Nahar, N.; Asaduzzaman, M.; Mandal, U.K.; Rimi, N.A.; Gurley, E.S.; Rahman, M.; Garcia, F.; Zimicki, S.; Sultana, R.; Luby, S.P. Hunting bats for human consumption in Bangladesh. EcoHealth 2020, 17, 139–151. [Google Scholar] [CrossRef]
- Latinne, A.; Saputro, S.; Kalengkongan, J.; Kowel, C.L.; Gaghiwu, L.; Ransaleleh, T.A.; Nangoy, M.J.; Wahyuni, I.; Kusumaningrum, T.; Safari, D.; et al. Characterizing and quantifying the wildlife trade network in Sulawesi, Indonesia. Glob. Ecol. Conserv. 2020, 21, e00887. [Google Scholar] [CrossRef]
- Wong, K.T.; Shieh, W.J.; Kumar, S.; Norain, K.; Abdullah, W.; Guarner, J.; Goldsmith, C.S.; Chua, K.B.; Lam, S.K.; Tan, C.T.; et al. Nipah virus infection: Pathology and pathogenesis of an emerging paramyxoviral zoonosis. Am. J. Pathol. 2002, 161, 2153–2167. [Google Scholar] [CrossRef]
- Rockx, B.; Bossart, K.N.; Feldmann, F.; Geisbert, J.B.; Hickey, A.C.; Brining, D.; Callison, J.; Safronetz, D.; Marzi, A.; Kercher, L.; et al. A novel model of lethal Hendra virus infection in African green monkeys and the effectiveness of ribavirin treatment. J. Virol. 2010, 84, 9831–9839. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, C.; Pohl, C.; Szecsi, J.; Trajkovic-Bodennec, S.; Devergnas, S.; Raoul, H.; Cosset, F.L.; Gerlier, D.; Wild, T.F.; Horvat, B. Nipah virus uses leukocytes for efficient dissemination within a host. J. Virol. 2011, 85, 7863–7871. [Google Scholar] [CrossRef] [Green Version]
- Halpin, K.; Hyatt, A.D.; Fogarty, R.; Middleton, D.; Bingham, J.; Epstein, J.H.; Rahman, S.A.; Hughes, T.; Smith, C.; Field, H.E.; et al. Pteropid bats are confirmed as the reservoir hosts of Henipaviruses: A comprehensive experimental study of virus transmission. Am. J. Trop. Med. Hyg. 2011, 85, 946–951. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, H.C.; Aspericueta, V.; Robinson, L.R.; Aanensen, K.E.; Lee, B. A quantitative and kinetic fusion protein-triggering assay can discern distinct steps in the Nipah virus membrane fusion cascade. J. Virol. 2010, 84, 8033–8041. [Google Scholar] [CrossRef] [Green Version]
- Stone, J.A.; Vemulapati, B.M.; Bradel-Tretheway, B.; Aguilar, H.C. Multiple strategies reveal a bidentate interaction between the Nipah virus attachment and fusion glycoproteins. J. Virol. 2016, 90, 10762–10773. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, C.; Dhondt, K.P.; Châlons, M.; Mély, S.; Raoul, H.; Negre, D.; Cosset, F.L.; Gerlier, D.; Vivès, R.R.; Horvat, B. Heparan sulfate-dependent enhancement of Henipavirus infection. MBio 2015, 6, e02427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weingartl, H.; Czub, S.; Copps, J.; Berhane, Y.; Middleton, D.; Marszal, P.; Gren, J.; Smith, G.; Ganske, S.; Manning, L.; et al. Invasion of the central nervous system in a porcine host by Nipah virus. J. Virol. 2005, 79, 7528–7534. [Google Scholar] [CrossRef] [PubMed]
- Park, M.S.; Shaw, M.L.; Muñoz-Jordan, J.; Cros, J.F.; Nakaya, T.; Bouvier, N.; Palese, P.; García-Sastre, A.; Basler, C.F. Newcastle disease virus (NDV)-based assay demonstrates interferon-antagonist activity for the NDV V protein and the Nipah virus V, W, and C proteins. J. Virol. 2003, 77, 1501–1511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virtue, E.R.; Marsh, G.A.; Baker, M.L.; Wang, L.F. Interferon production and signaling pathways are antagonized during Henipavirus infection of fruit bat cell lines. PLoS ONE 2011, 6, e22488. [Google Scholar] [CrossRef] [Green Version]
- Gutsche, I.; Desfosses, A.; Effantin, G.; Ling, W.L.; Haupt, M.; Ruigrok, R.W.H.; Sachse, C.; Schoehn, G. Near-atomic cryo-EM structure of the helical measles virus nucleocapsid. Science 2015, 348, 704–707. [Google Scholar] [CrossRef]
- Alayyoubi, M.; Leser, G.P.; Kors, C.A.; Lamb, R.A. Structure of the paramyxovirus parainfluenza virus 5 nucleoprotein–RNA complex. Proc. Natl. Acad. Sci. USA 2015, 112, E1792–E1799. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Shan, H.; Zhu, Y.; Hu, S.; Xue, L.; Chen, Y.; Ding, W.; Niu, T.; Gu, J.; Ouyang, S.; et al. Self-capping of nucleoprotein filaments protects the Newcastle disease virus genome. eLife 2019, 8, e45057. [Google Scholar] [CrossRef]
- Tan, W.S.; Ong, S.T.; Eshaghi, M.; Foo, S.S.; Yusoff, K. Solubility, immunogenicity and physical properties of the nucleocapsid protein of Nipah virus produced in Escherichia coli. J. Med. Virol. 2004, 73, 105–112. [Google Scholar] [CrossRef]
- Joseph, N.M.S.; Tey, B.T.; Tan, C.S.; Shafee, N.; Tan, W.S. Production of long helical capsid of Nipah virus by Pichia pastoris. Process Biochem. 2011, 46, 1871–1874. [Google Scholar] [CrossRef]
- Eshaghi, M.; Tan, W.S.; Ong, S.T.; Yusoff, K. Purification and characterization of Nipah virus nucleocapsid protein produced in insect cells. J. Clin. Microbiol. 2005, 43, 3172–3177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chua, K.B.; Lam, S.K.; Goh, K.J.; Hooi, P.S.; Ksiazek, T.G.; Kamarulzaman, A.; Olson, J.; Tan, C.T. The presence of Nipah virus in respiratory secretions and urine of patients during an outbreak of Nipah virus encephalitis in Malaysia. J. Infect. 2001, 42, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Escaffre, O.; Borisevich, V.; Rockx, B. Pathogenesis of Hendra and Nipah virus infection in humans. J. Infect. Dev. Ctries. 2013, 7, 308–311. [Google Scholar] [CrossRef] [PubMed]
- Munster, V.J.; Prescott, J.B.; Bushmaker, T.; Long, D.; Rosenke, R.; Thomas, T.; Scott, D.; Fischer, E.R.; Feldmann, H.; de Wit, E. Rapid Nipah virus entry into the central nervous system of hamsters via the olfactory route. Sci. Rep. 2012, 2, 736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rockx, B.; Brining, D.; Kramer, J.; Callison, J.; Ebihara, H.; Mansfield, K.; Feldmann, H. Clinical outcome of Henipavirus infection in hamsters is determined by the route and dose of infection. J. Virol. 2011, 85, 7658–7671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, S.; Volchkova, V.; Basler, C.F.; Palese, P.; Volchkov, V.E.; Shaw, M.L. Nipah virus edits its P gene at high frequency to express the V and W proteins. J. Virol. 2009, 83, 3982–3987. [Google Scholar] [CrossRef] [Green Version]
- Andrejeva, J.; Childs, K.S.; Young, D.F.; Carlos, T.S.; Stock, N.; Goodbourn, S.; Randall, R.E. The V proteins of paramyxoviruses bind the IFN-inducible RNA helicase, mda-5, and inhibit its activation of the IFN-beta promoter. Proc. Natl. Acad. Sci. USA 2004, 101, 17264–17269. [Google Scholar] [CrossRef] [Green Version]
- Shaw, M.L.; Cardenas, W.B.; Zamarin, D.; Palese, P.; Basler, C.F. Nuclear localization of the Nipah virus W protein allows for inhibition of both virus- and toll-like receptor 3-triggered signaling pathways. J. Virol. 2005, 79, 6078–6088. [Google Scholar] [CrossRef] [Green Version]
- Pelissier, R.; Iampietro, M.; Horvat, B. Recent advances in the understanding of Nipah virus immunopathogenesis and anti-viral approaches. F1000Research 2019, 8, 1763. [Google Scholar] [CrossRef]
- Yoneda, M.; Guillaume, V.; Sato, H.; Fujita, K.; Georges-Courbot, M.C.; Ikeda, F.; Omi, M.; Muto-Terao, Y.; Wild, T.F.; Kai, C. The nonstructural proteins of Nipah virus play a key role in pathogenicity in experimentally infected animals. PLoS ONE 2010, 5, e12709. [Google Scholar] [CrossRef] [Green Version]
- Middleton, D.J.; Morrissy, C.J.; van der Heide, B.M.; Russell, G.M.; Braun, M.A.; Westbury, H.A.; Halpin, K.; Daniels, P.W. Experimental Nipah virus infection in pteropid bats (Pteropus poliocephalus). J. Comp. Pathol. 2007, 136, 266–272. [Google Scholar] [CrossRef]
- Halpin, K.; Bankamp, B.; Harcourt, B.H.; Bellini, W.J.; Rota, P.A. Nipah virus conforms to the rule of six in a minigenome replication assay. J. Gen. Virol. 2004, 85, 701–707. [Google Scholar] [CrossRef]
- Gauldie, J.; Richards, C.; Harnish, D.; Lansdorp, P.; Baumann, H. Interferon beta 2/B-cell stimulatory factor type 2 shares identity with monocyte-derived hepatocyte-stimulating factor and regulates the major acute phase protein response in liver cells. Proc. Natl. Acad. Sci. USA 1987, 84, 7251–7255. [Google Scholar] [CrossRef]
- Mahalingam, S.; Farber, J.M.; Karupiah, G. The interferon-inducible chemokines MuMig and Crg-2 exhibit antiviral activity in vivo. J. Virol. 1999, 73, 1479–1491. [Google Scholar] [CrossRef] [Green Version]
- Lei, J.; Yin, X.; Shang, H.; Jiang, Y. IP-10 is highly involved in HIV infection. Cytokine 2019, 115, 97–103. [Google Scholar] [CrossRef]
- Lo, M.K.; Miller, D.; Aljofan, M.; Mungall, B.A.; Rollin, P.E.; Bellini, W.J.; Rota, P.A. Characterization of the antiviral and inflammatory responses against Nipah virus in endothelial cells and neurons. Virology 2010, 404, 78–88. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, C.; Guillaume, V.; Sabine, A.; Ong, K.C.; Wong, K.T.; Legras-Lachuer, C.; Horvat, B. Lethal Nipah virus infection induces rapid overexpression of CXCL10. PLoS ONE 2012, 7, e32157. [Google Scholar] [CrossRef] [Green Version]
- Georges-Courbot, M.C.; Contamin, H.; Faure, C.; Loth, P.; Baize, S.; Leyssen, P.; Neyts, J.; Deubel, V. Poly(I)-poly(C12U) but not ribavirin prevents death in a hamster model of Nipah virus infection. Antimicrob. Agents Chemother. 2006, 50, 1768–1772. [Google Scholar] [CrossRef] [Green Version]
- Ramasundram, V.; Tan, C.T.; Chua, K.B.; Chong, H.T.; Goh, K.J.; Chew, N.K.; Tan, K.S.; Thayaparan, T.; Kunjapan, S.R.; Petharunam, V.; et al. Kinetics of IgM and IgG seroconversion in Nipah virus infection. Neurol. J. Southeast Asia 2000, 5, 6. [Google Scholar]
- Hooper, P.; Zaki, S.; Daniels, P.; Middleton, D. Comparative pathology of the diseases caused by Hendra and Nipah viruses. Microbes Infect. 2001, 3, 315–322. [Google Scholar] [CrossRef]
- Wong, S.C.; Ooi, M.H.; Wong, M.N.; Tio, P.H.; Solomon, T.; Cardosa, M.J. Late presentation of Nipah virus encephalitis and kinetics of the humoral immune response. J. Neurol. Neurosurg. Psychiatry 2001, 71, 552–554. [Google Scholar] [CrossRef]
- Middleton, D.J.; Westbury, H.A.; Morrissy, C.J.; van der Heide, B.M.; Russell, G.M.; Braun, M.A.; Hyatt, A.D. Experimental Nipah virus infection in pigs and cats. J. Comp. Pathol. 2002, 126, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.T.; Grosjean, I.; Brisson, C.; Blanquier, B.; Fevre-Montange, M.; Bernard, A.; Loth, P.; Georges-Courbot, M.C.; Chevallier, M.; Akaoka, H.; et al. A golden hamster model for human acute Nipah virus infection. Am. J. Pathol. 2003, 163, 2127–2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossart, K.N.; Zhu, Z.; Middleton, D.; Klippel, J.; Crameri, G.; Bingham, J.; McEachern, J.A.; Green, D.; Hancock, T.J.; Chan, Y.P.; et al. A neutralizing human monoclonal antibody protects against lethal disease in a new ferret model of acute Nipah virus infection. PLoS Pathog. 2009, 5, e1000642. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Daddario-DiCaprio, K.M.; Hickey, A.C.; Smith, M.A.; Chan, Y.P.; Wang, L.F.; Mattapallil, J.J.; Geisbert, J.B.; Bossart, K.N.; Broder, C.C. Development of an acute and highly pathogenic nonhuman primate model of Nipah virus infection. PLoS ONE 2010, 5, e10690. [Google Scholar] [CrossRef]
- Dhondt, K.P.; Mathieu, C.; Chalons, M.; Reynaud, J.M.; Vallve, A.; Raoul, H.; Horvat, B. Type I interferon signaling protects mice from lethal Henipavirus infection. J. Infect. Dis. 2013, 207, 142–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalodimou, G.; Veit, S.; Jany, S.; Kalinke, U.; Broder, C.C.; Sutter, G.; Volz, A. A soluble version of Nipah virus glycoprotein G delivered by vaccinia virus MVA activates specific CD8 and CD4 T cells in mice. Viruses 2019, 12, 26. [Google Scholar] [CrossRef] [Green Version]
- Guillaume, V.; Contamin, H.; Loth, P.; Georges-Courbot, M.C.; Lefeuvre, A.; Marianneau, P.; Chua, K.B.; Lam, S.K.; Buckland, R.; Deubel, V.; et al. Nipah virus: Vaccination and passive protection studies in a hamster model. J. Virol. 2004, 78, 834–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossart, K.N.; Geisbert, T.W.; Feldmann, H.; Zhu, Z.; Feldmann, F.; Geisbert, J.B.; Yan, L.; Feng, Y.R.; Brining, D.; Scott, D.; et al. A neutralizing human monoclonal antibody protects African green monkeys from Hendra virus challenge. Sci. Transl. Med. 2011, 3, 105ra103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arunkumar, G.; Devadiga, S.; McElroy, A.K.; Prabhu, S.; Sheik, S.; Abdulmajeed, J.; Robin, S.; Sushama, A.; Jayaram, A.; Nittur, S.; et al. Adaptive immune responses in humans during Nipah virus acute and convalescent phases of infection. Clin. Infect. Dis. 2019, 69, 1752–1756. [Google Scholar] [CrossRef] [PubMed]
- Stachowiak, B.; Weingartl, H.M. Nipah virus infects specific subsets of porcine peripheral blood mononuclear cells. PLoS ONE 2012, 7, e30855. [Google Scholar] [CrossRef]
- World Organization for Animal Health (OIE). Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2022 (OIE Terrestrial Manual 2022). Available online: https://www.woah.org/en/what-we-do/standards/codes-and-manuals/terrestrial-manual-online-access/ (accessed on 22 December 2022).
- Rahman, S.A.; Hassan, L.; Sharifah, S.H.; Lazarus, K.; Zaini, C.M.; Epstein, J.H.; Shamsyul Naim, N.; Field, H.E.; Arshad, S.S.; Abdul Aziz, J.; et al. Evidence for Nipah virus recrudescence and serological patterns of captive Pteropus vampyrus. Epidemiol. Infect. 2011, 139, 1570–1579, Erratum in: Epidemiol. Infect. 2011, 139, 1580. [Google Scholar] [CrossRef] [Green Version]
- Rahman, S.A.; Hassan, L.; Epstein, J.H.; Zaini, C.M.; Azizi, M.Y.; Sharifah, S.H.; Field, H.E.; Tom, H.; Justin, W.; Naim, S.N.; et al. Risk factors for Nipah virus infections among pteropid bats, Peninsular Malaysia. Emerg. Infect. Dis. 2013, 1, 51–60. [Google Scholar] [CrossRef]
- Wacharapluesadee, S.; Lumlertdacha, B.; Boongird, K.; Wanghongsa, S.; Chanhome, L.; Rollin, P.; Stockton, P.; Rupprecht, C.E.; Ksiazek, T.G.; Hemachudha, T. Bat Nipah virus, Thailand. Emerg. Infect. Dis. 2005, 11, 1949–1951. [Google Scholar] [CrossRef]
- Wacharapluesadee, S.; Hemachudha, T. Duplex nested RT-PCR for detection of Nipah virus RNA from urine specimens of bats. J. Virol. Methods 2007, 141, 97–101. [Google Scholar] [CrossRef]
- Aziz, A.J.; Mahendran, R.; Daniels, P.; Shahiruddin, S.; Narasiman, M.; Azizah, D.; Johara, M.Y. The status, public response and challenges in overcoming emerging and exotic diseases—Nipah virus disease experience. In Proceedings of the National Congress on Animal Health and Production: Environmental Care in Animal Production 1999, Alor Gajah (Malacca), Malaysia, 3–5 September 1999; Available online: https://www.researchgate.net/profile/Muniandy-Narasiman-2/publication/280224022_THE_STATUS_PUBLIC_RESPONSE_AND_CHALLENGES_IN_OVERCOMING_EMERGING_AND_EXOTIC_DISEASES_-_NIPAH_VIRUS_DISEASE_EXPERIENCE/links/55adff0408aee079921e4cbe/THE-STATUS-PUBLIC-RESPONSE-AND-CHALLENGES-IN-OVERCOMING-EMERGING-AND-EXOTIC-DISEASES-NIPAH-VIRUS-DISEASE-EXPERIENCE.pdf (accessed on 22 December 2022).
- Berhane, Y.; Weingartl, H.M.; Lopez, J.; Neufeld, J.; Czub, S.; Embury-Hyatt, C.; Goolia, M.; Copps, J.; Czub, M. Bacterial infections in pigs experimentally infected with Nipah virus. Transbound. Emerg. Dis. 2008, 55, 165–174. [Google Scholar] [CrossRef]
- Abdullah, S.; Chang, L.Y.; Rahmat, K.; Goh, K.J.; Tan, C.T. Late-onset Nipah virus encephalitis 11 years after the initial outbreak: A case report. Neurol. Asia 2012, 17, 71–74. [Google Scholar]
- Ng, B.Y.; Lim, C.C.; Yeoh, A.; Lee, W.L. Neuropsychiatric sequelae of Nipah virus encephalitis. J. Neuropsychiatry Clin. Neurosci. 2004, 16, 500–504. [Google Scholar] [CrossRef]
- Tan, C.T.; Goh, K.J.; Wong, K.T.; Sarji, S.A.; Chua, K.B.; Chew, N.K.; Murugasu, P.; Loh, Y.L.; Chong, H.T.; Tan, K.S.; et al. Relapsed and late onset Nipah encephalitis. Ann. Neurol. 2002, 51, 703–708. [Google Scholar] [CrossRef]
- McLean, R.K.; Graham, S.P. Vaccine development for Nipah virus infection in pigs. Front. Vet. Sci. 2019, 6, 16. [Google Scholar] [CrossRef] [Green Version]
- Daniels, P.; Ksiazek, T.; Eaton, B.T. Laboratory diagnosis of Nipah and Hendra virus infections. Microbes Infect. 2001, 3, 289–295. [Google Scholar] [CrossRef]
- Chong, H.T.; Kunjappan, S.R.; Thayaparan, T.; Tong, J.; Petharunam, V.; Jusoh, M.R.; Tan, C.T. Nipah encephalitis outbreak in Malaysia, clinical features in patients from Seremban. Neurol. J. Southeast Asia 2000, 5, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Sarji, S.A.; Abdullah, B.J.; Goh, K.J.; Tan, C.T.; Wong, K.T. Magnetic resonance imaging features of Nipah encephalitis. Am. J. Roentgenol. 2000, 175, 437–442. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.S.; Ahmad Sarji, S.; Tan, C.T.; Abdullah, B.J.J.; Chong, H.T.; Thayaparan, T.; Koh, C.N. Patients with asymptomatic Nipah virus infection may have abnormal cerebral MR imaging. Neurol. J. Southeast Asia 2000, 5, 69–73. [Google Scholar]
- Chiang, C.F.; Lo, M.K.; Rota, P.A.; Spiropoulou, C.F.; Rollin, P.E. Use of monoclonal antibodies against Hendra and Nipah viruses in an antigen capture-ELISA. Virol. J. 2010, 7, 115. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Khairullah, N.S.; Inoue, S.; Balasubramaniam, V.; Berendam, S.J.; Teh, L.K.; Ibrahim, N.S.W.; Abdul Rahman, S.; Hassan, S.S.; Hasebe, F.; et al. Serodiagnosis using recombinant Nipah virus nucleocapsid protein expressed in Escherichia coli. J. Clin. Microbiol. 2006, 44, 3134–3138. [Google Scholar] [CrossRef]
- Kulkarni, D.D.; Tosh, C.; Venkatesh, G.; Senthil Kumar, D. Nipah virus infection: Current scenario. Indian J. Virol. 2013, 24, 398–408. [Google Scholar] [CrossRef] [Green Version]
- Ong, K.C.; Wong, K.T. Henipavirus encephalitis: Recent developments and advances. Brain Pathol. 2015, 25, 605–613. [Google Scholar] [CrossRef]
- Kaku, Y.; Noguchi, A.; Marsh, G.A.; Barr, J.A.; Okutani, A.; Hotta, K.; Bazartseren, B.; Broder, C.C.; Yamada, A.; Inoue, S.; et al. Antigens capture ELISA system for Henipaviruses using polyclonal antibodies obtained by DNA immunization. Arch. Virol. 2012, 157, 1605–1609. [Google Scholar] [CrossRef]
- Wang, L.F.; Daniels, P. Diagnosis of Henipavirus infection: Current capabilities and future directions. Curr. Top. Microbiol. Immunol. 2012, 359, 179–196. [Google Scholar] [CrossRef]
- Kaku, Y.; Noguchi, A.; Marsh, G.A.; McEachern, J.A.; Okutani, A.; Hotta, K.; Bazartseren, B.; Fukushi, S.; Broder, C.C.; Yamada, A.; et al. A neutralization test for specific detection of Nipah virus antibodies using pseudotyped vesicular stomatitis virus expressing green fluorescent protein. J. Virol. Methods 2009, 160, 7–13. [Google Scholar] [CrossRef]
- Guillaume, V.; Lefeuvre, A.; Faure, C.; Marianneau, P.; Buckland, R.; Lam, S.K.; Wild, T.F.; Deubel, V. Specific detection of Nipah virus using real-time RT-PCR (TaqMan). J. Virol. Methods 2004, 120, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Mazzola, L.T.; Kelly-Cirino, C. Diagnostics for Nipah virus: A zoonotic pathogen endemic to Southeast Asia. BMJ Global Health 2019, 4, e001118. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.Y.; Ali, A.R.; Hassan, S.S.; AbuBakar, S. Quantitative estimation of Nipah virus replication kinetics in vitro. Virol. J. 2006, 3, 47. [Google Scholar] [CrossRef] [Green Version]
- Jensen, K.S.; Adams, R.; Bennett, R.S.; Bernbaum, J.; Jahrling, P.B.; Holbrook, M.R. Development of a novel real-time polymerase chain reaction assay for the quantitative detection of Nipah virus replicative viral RNA. PLoS ONE 2018, 13, e0199534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkland, P.D.; Stephano, H.; Weingartl, H.M. Paramyxoviruses. In Diseases of Swine, 11th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 594–610. [Google Scholar] [CrossRef]
- Sauerhering, L.; Elvert, M.; Behner, L.; Matrosovich, T.; Erbar, S.; Matrosovich, M.; Maisner, A. Species-specific and individual differences in Nipah virus replication in porcine and human airway epithelial cells. J. Gen. Virol. 2016, 97, 1511–1519. [Google Scholar] [CrossRef] [PubMed]
- Westbury, H.A.; Hooper, P.T.; Brouwer, S.L.; Selleck, P.W. Susceptibility of cats to equine morbillivirus. Aust. Vet. J. 1996, 74, 132–134. [Google Scholar] [CrossRef] [PubMed]
- Mungall, B.A.; Middleton, D.; Crameri, G.; Halpin, K.; Bingham, J.; Eaton, B.T.; Broder, C.C. Vertical transmission and fetal replication of Nipah virus in an experimentally infected cat. J. Infect. Dis. 2007, 196, 812–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, M.M.; Hooper, P.T.; Selleck, P.W.; Westbury, H.A.; Slocombe, R.F. A guinea-pig model of Hendra virus encephalitis. J. Comp. Pathol. 2001, 124, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Tanimura, N.; Imada, T.; Kashiwazaki, Y.; Sharifah, S.H. Distribution of viral antigens and development of lesions in chicken embryos inoculated with nipah virus. J. Comp. Pathol. 2006, 135, 74–82. [Google Scholar] [CrossRef]
- Mire, C.E.; Satterfield, B.A.; Geisbert, J.B.; Agans, K.N.; Borisevich, V.; Yan, L.; Chan, Y.P.; Cross, R.W.; Fenton, K.A.; Broder, C.C.; et al. Pathogenic differences between Nipah virus Bangladesh and Malaysia strains in primates: Implications for antibody therapy. Sci. Rep. 2016, 6, 30916. [Google Scholar] [CrossRef] [Green Version]
- Marianneau, P.; Guillaume, V.; Wong, T.; Badmanathan, M.; Looi, R.Y.; Murri, S.; Loth, P.; Tordo, N.; Wild, F.; Horvat, B.; et al. Experimental infection of squirrel monkeys with Nipah virus. Emerg. Infect. Dis. 2010, 16, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.K.; Chua, K.B. Nipah virus encephalitis outbreak in Malaysia. Clin. Infect. Dis. 2002, 34, S48–S51. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization (FAO) of the United Nations, Regional Office for Asia and the Pacific. Manual on the Diagnosis of Nipah Virus Infection in Animals; RAP Publication No. 2002/01; Food and Agriculture Organization of the United Nations, Regional Office for Asia and the Pacific: Bangkok, Thailand, 2002. [Google Scholar]
- Playford, E.G.; Munro, T.; Mahler, S.M.; Elliott, S.; Gerometta, M.; Hoger, K.L.; Jones, M.L.; Griffin, P.; Lynch, K.D.; Carroll, H. Safety, tolerability, pharmacokinetics, and immunogenicity of a human monoclonal antibody targeting the G glycoprotein of Henipaviruses in healthy adults: A first-in-human, randomised, controlled, phase 1 study. Lancet Infect. Dis. 2020, 20, 445–454. [Google Scholar] [CrossRef]
- Pallister, J.; Middleton, D.; Wang, L.F.; Klein, R.; Haining, J.; Robinson, R.; Yamada, M.; White, J.; Payne, J.; Feng, Y.R.; et al. A recombinant Hendra virus G glycoprotein-based subunit vaccine protects ferrets from lethal Hendra virus challenge. Vaccine 2011, 29, 5623–5630. [Google Scholar] [CrossRef]
- Mire, C.E.; Geisbert, J.B.; Agans, K.N.; Feng, Y.R.; Fenton, K.A.; Bossart, K.N.; Yan, L.; Chan, Y.P.; Broder, C.C.; Geisbert, T.W. A recombinant Hendra virus G glycoprotein subunit vaccine protects nonhuman primates against Hendra virus challenge. J. Virol. 2014, 88, 4624–4631. [Google Scholar] [CrossRef] [Green Version]
- Pickering, B.S.; Hardham, J.M.; Smith, G.; Weingartl, E.T.; Dominowski, P.J.; Foss, D.L.; Mwangi, D.; Broder, C.C.; Roth, J.A.; Weingartl, H.M. Protection against Henipaviruses in swine requires both cell-mediated and humoral immune response. Vaccine 2016, 34, 4777–4786. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Bobb, K.; Borisevich, V.; Geisbert, J.B.; Agans, K.N.; Cross, R.W.; Prasad, A.N.; Fenton, K.A.; Yu, H.; Fouts, T.R.; et al. A single dose investigational subunit vaccine for human use against Nipah virus and Hendra virus. NPJ Vaccines 2021, 6, 23. [Google Scholar] [CrossRef]
- Wang, Z.; Amaya, M.; Addetia, A.; Dang, H.V.; Reggiano, G.; Yan, L.; Hickey, A.C.; DiMaio, F.; Broder, C.C.; Veesler, D. Architecture and antigenicity of the Nipah virus attachment glycoprotein. Science 2022, 375, 1373–1378. [Google Scholar] [CrossRef]
- Predictor of Natural Disordered Regions (PONDR® VLXT). Available online: http://pondr.com/ (accessed on 22 December 2022).
- Goh, G.K.; Dunker, A.K.; Foster, J.A.; Uversky, V.N. Nipah shell disorder, modes of infection, and virulence. Microb Pathog. 2020, 141, 103976. [Google Scholar] [CrossRef]
- Goh, G.K.; Dunker, A.K.; Foster, J.A.; Uversky, V.N. A novel strategy for the development of vaccines for SARS-CoV-2 (COVID-19) and other viruses using AI and viral shell disorder. J. Proteome Res. 2020, 19, 4355–4363. [Google Scholar] [CrossRef]
- European Union (EU). Commission Delegated Regulation (EU) 2018/1629 of 25 July 2018 Amending the List of Diseases Set Out in Annex II to Regulation (EU) 2016/429 of the European Parliament and of the Council on Transmissible Animal Diseases and Amending and Repealing Certain Acts in the Area of Animal Health (‘Animal Health Law’). Off. J. 2018. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32018R1629&from=EN (accessed on 22 December 2022).
- European Union (EU). Commission Implementing Regulation (EU) 2018/1882 of 3 December 2018 on the Application of Certain Disease Prevention and Control Rules to Categories of Listed Diseases and Establishing a List of Species and Groups of Species Posing a Considerable Risk for the Spread of Those Listed Diseases. Off. J. 2018. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32018R1882&qid=1669988175289&from=EN (accessed on 22 December 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Initial Source of Infection | Intermediate Host | Source of Infection for Humans | Transmission | |
|---|---|---|---|---|---|
| Malaysia Singapore | NiV-M | Bat-bitten fruit | Pig | Pig excretions Pig secretions Aerosol Raw meat/products Direct contact Aerosol Fomites | Pig-to-human (main route) Human-to-human |
| Bangladesh | NiV-B | Bat saliva-contaminated and excreta-contaminated palm sap | – | Raw palm sap Direct contact Aerosol Fomites | Bat-to-human Human-to-human (especially nosocomial) |
| India | NiV-B | Bat-bitten fruit | – | Fruit Direct contact Aerosol Fomites | Bat-to-human Human-to-human (especially nosocomial) |
| Philippines | NiV-M | Bat-bitten fruit (most likely) | Horse | Direct contact Horse secretions Raw meat Direct contact Aerosol Fomites | Horse-to-human Human-to-human (also nosocomial) |
| Inflammation and Innate Immunity | References |
|---|---|
| [50,165,172] |
| [50,165] |
| [50,165,172] |
| [50,165] |
| [50,165] |
| [50,159,172] |
| Adaptive Immunity | References |
|---|---|
| Antibody and B cell responses | |
| Humans: | |
| [50,175,186] |
| [50,165,186] |
| Experimentally-infected pigs: | |
| [50] |
| Experimentally-infected AGM: | |
| [50,144] |
| [50] |
| [50] |
| T cell responses | |
| Humans: | |
| [50,165,186] |
| [159,187] |
| Experimentally-infected pigs: | |
| [50,165,187] |
| Experimentally-infected AGM: | |
| [50,165] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruno, L.; Nappo, M.A.; Ferrari, L.; Di Lecce, R.; Guarnieri, C.; Cantoni, A.M.; Corradi, A. Nipah Virus Disease: Epidemiological, Clinical, Diagnostic and Legislative Aspects of This Unpredictable Emerging Zoonosis. Animals 2023, 13, 159. https://doi.org/10.3390/ani13010159
Bruno L, Nappo MA, Ferrari L, Di Lecce R, Guarnieri C, Cantoni AM, Corradi A. Nipah Virus Disease: Epidemiological, Clinical, Diagnostic and Legislative Aspects of This Unpredictable Emerging Zoonosis. Animals. 2023; 13(1):159. https://doi.org/10.3390/ani13010159
Chicago/Turabian StyleBruno, Luigi, Maria Anna Nappo, Luca Ferrari, Rosanna Di Lecce, Chiara Guarnieri, Anna Maria Cantoni, and Attilio Corradi. 2023. "Nipah Virus Disease: Epidemiological, Clinical, Diagnostic and Legislative Aspects of This Unpredictable Emerging Zoonosis" Animals 13, no. 1: 159. https://doi.org/10.3390/ani13010159