
A Cautionary Tale of Sexing by Methylation: Hybrid Bisulfite-Conversion Sequencing of Immunoprecipitated Methylated DNA in Chrysemys picta Turtles with Temperature-Dependent Sex Determination Reveals Contrasting Patterns of Somatic and Gonadal Methylation, but No Unobtrusive Sex Diagnostic

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Collection and DNA Extraction
2.2. Global Methylation Analysis via ELISA
2.3. MeDIP-BS-Seq Library Construction and Sequencing
2.4. Methylome Assembly and Analysis
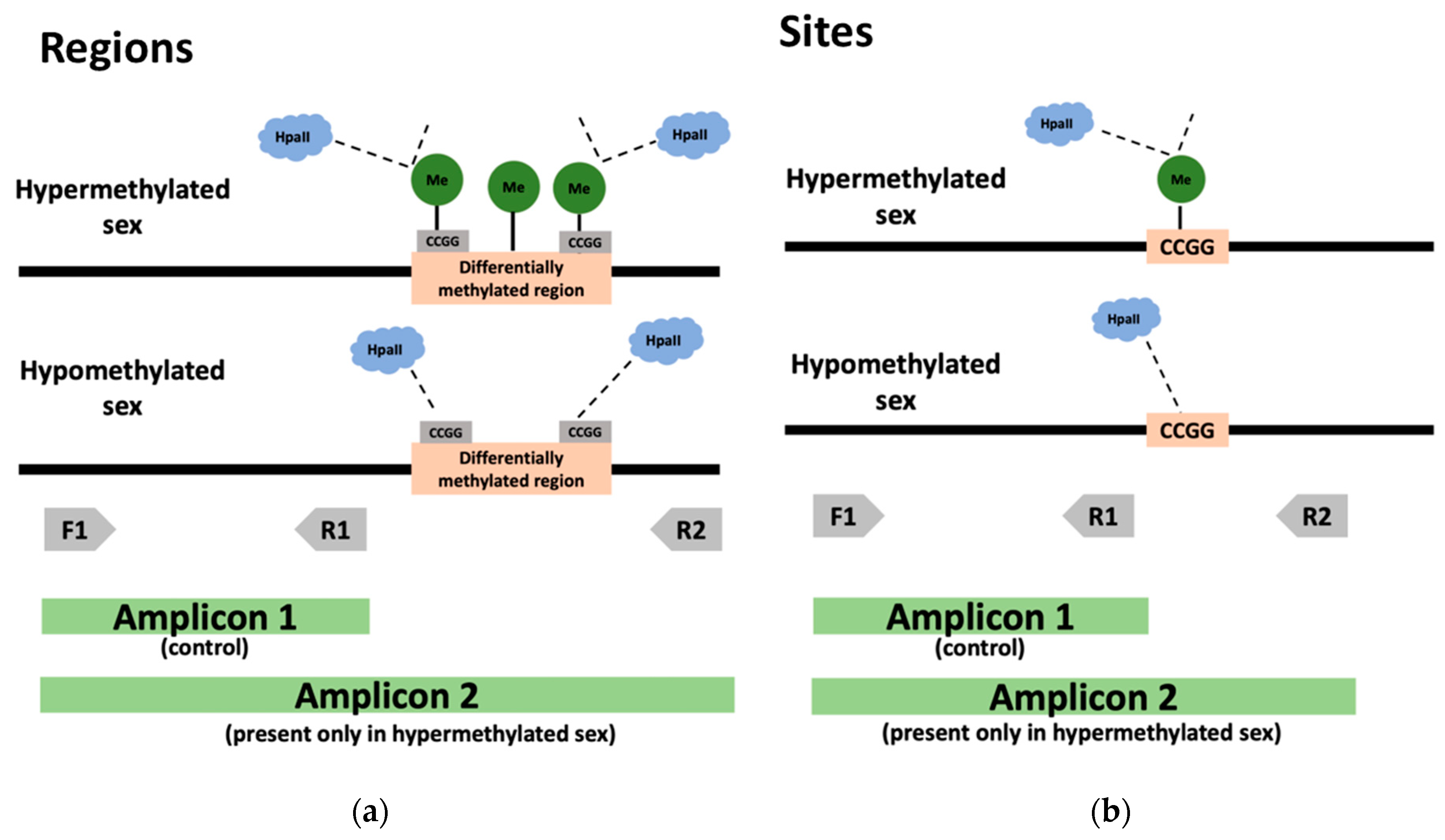
2.5. DNA Digestion and Methylome Validation by PCR
3. Results
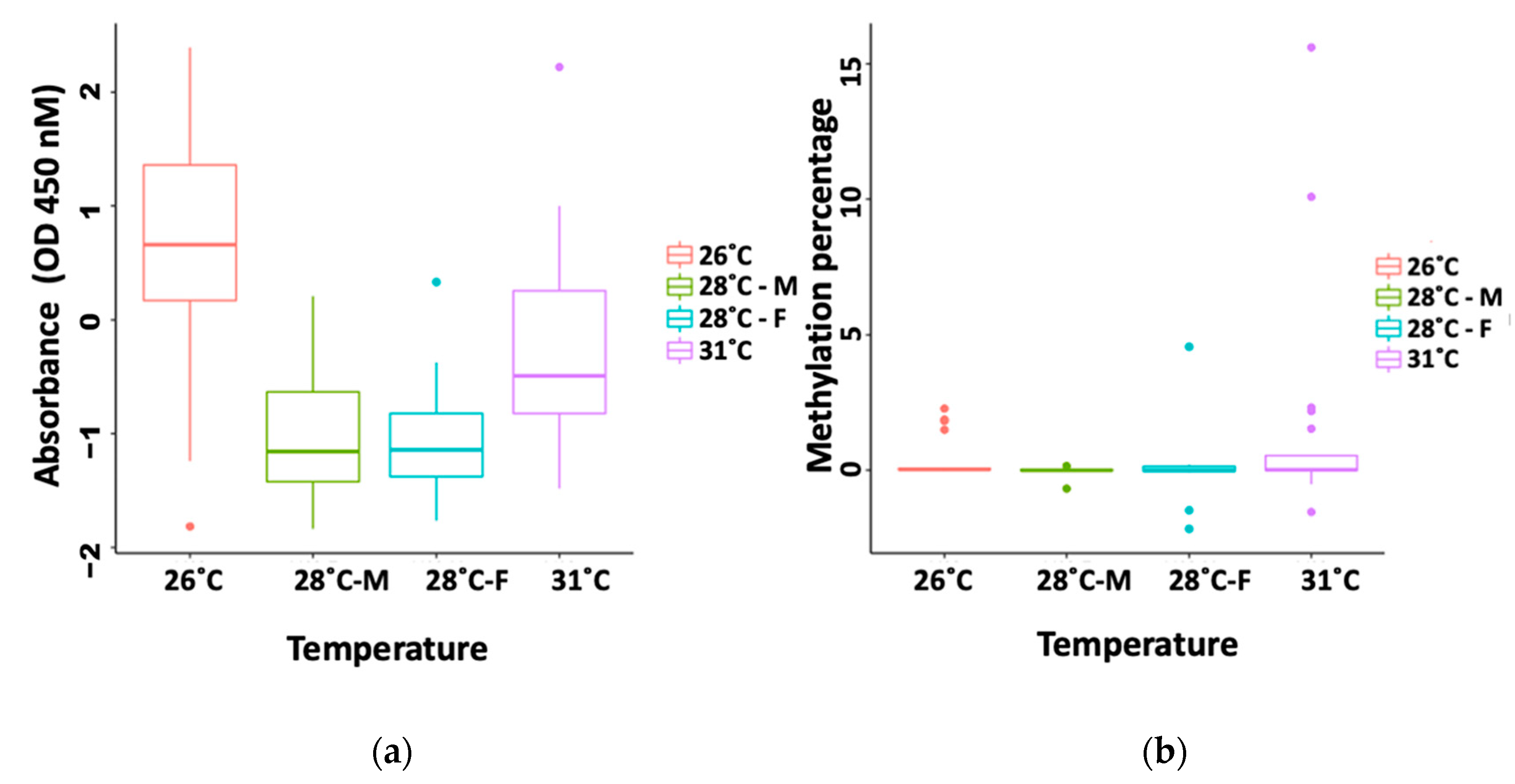
3.1. Global Methylation by ELISA
3.2. MeDIP-BS-Seq Methylome Assembly and Analysis
3.3. Methylation-Sensitive PCR
4. Discussion
4.1. A Novel Complexity-Reduction and Site-by-Site Approach for DNA Methylation Analysis
4.2. Inter-Individual Variation in DNA Methylation Exists in Both Males and Females
4.3. Contrasting Patterns of DNA Methylation between Somatic and Gonadal Tissues
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vandegehuchte, M.B.; Janssen, C.R. Epigenetics and its implications for ecotoxicology. Ecotoxicology 2011, 20, 607–624. [Google Scholar] [CrossRef] [PubMed]
- Mizoguchi, B.A.; Valenzuela, N. Ecotoxicological Perspectives of Sex Determination. Sex. Dev. 2016, 10, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Senner, C.E. The role of DNA methylation in mammalian development. Reprod. Biomed. Online 2011, 22, 529–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piferrer, F. Epigenetics of sex determination and gonadogenesis. Dev. Dyn. 2013, 242, 360–370. [Google Scholar] [CrossRef]
- Cedar, H. DNA methylation and gene activity. Cell 1988, 53, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, S.; Literman, R.; Mizoguchi, B.; Valenzuela, N. MeDIP-seq and nCpG analyses illuminate sexually dimorphic methylation of gonadal development genes with high historic methylation in turtle hatchlings with temperature-dependent sex determination. Epigenetics Chromatin 2017, 10, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elango, N.; Hunt, B.G.; Goodisman, M.A.D.; Yi, S.V. DNA methylation is widespread and associated with differential gene expression in castes of the honeybee, Apis mellifera. Proc. Natl. Acad. Sci. USA 2009, 106, 11206–11211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, Y.; Buemio, A.; Chu, R.; Vafaee, M.; Crews, D. Epigenetic Control of Gonadal Aromatase (cyp19a1) in Temperature-Dependent Sex Determination of Red-Eared Slider Turtles. PLoS ONE 2013, 8, e63599. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Hannigan, B.; Crews, D. Temperature Shift Alters DNA Methylation and Histone Modification Patterns in Gonadal Aromatase (cyp19a1) Gene in Species with Temperature-Dependent Sex Determination. PLoS ONE 2016, 11, e0167362. [Google Scholar] [CrossRef] [Green Version]
- Venegas, D.; Marmolejo-Valencia, A.; Valdes-Quezada, C.; Govenzensky, T.; Recillas-Targa, F.; Merchant-Larios, H. Dimorphic DNA methylation during temperature-dependent sex determination in the sea turtle Lepidochelys olivacea. Gen. Comp. Endocrinol. 2016, 236, 35–41. [Google Scholar] [CrossRef]
- Ge, C.; Ye, J.; Zhang, H.; Zhang, Y.; Sun, W.; Sang, Y.; Capel, B.; Qian, G. Dmrt1 induces the male pathway in a turtle with temperature-dependent sex determination. Development 2017, 144, 2222–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrott, B.B.; Kohno, S.; Cloy-McCoy, J.A.; Guillette, L.J. Differential Incubation Temperatures Result in Dimorphic DNA Methylation Patterning of the SOX9 and Aromatase Promoters in Gonads of Alligator (Alligator mississippiensis) Embryos1. Biol. Reprod. 2014, 90, 2. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.-X.; Wang, Y.-Y.; Zhao, Y.; Wang, H.; Li, N.; Ji, X.S. Global DNA Methylation Changes in Nile Tilapia Gonads during High Temperature-Induced Masculinization. PLoS ONE 2016, 11, e0158483. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Martín, L.; Viñas, J.; Ribas, L.; Díaz, N.; Gutiérrez, A.; Di Croce, L.; Piferrer, F. DNA Methylation of the Gonadal Aromatase (cyp19a) Promoter Is Involved in Temperature-Dependent Sex Ratio Shifts in the European Sea Bass. PLOS Genet. 2011, 7, e1002447. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, N.; Adams, D.C.; Janzen, F.J. Pattern Does Not Equal Process: Exactly When Is Sex Environmentally Determined? Am. Nat. 2003, 161, 676–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, M. Sexual Selection, 1st ed.; Princeton University Press: Princeton, NJ, USA, 1994. [Google Scholar]
- Valenzuela, N.; Lance, V. Temperature-Dependent Sex Determination in Vertebrates, 1st ed.; Smithsonian Books: Washington, DC, USA, 2004. [Google Scholar]
- Mrosovsky, N. Sex ratio bias in hatchling sea turtles from artificially incubated eggs. Biol. Conserv. 1982, 23, 309–314. [Google Scholar] [CrossRef]
- Valenzuela, N.; Literman, R.; Neuwald, J.L.; Mizoguchi, B.; Iverson, J.B.; Riley, J.L.; Litzgus, J.D. Extreme thermal fluctuations from climate change unexpectedly accelerate demographic collapse of vertebrates with temperature-dependent sex determination. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holleley, C.E.; O’Meally, D.; Sarre, S.D.; Graves, J.A.M.; Ezaz, T.; Matsubara, K.; Azad, B.; Zhang, X.; Georges, A. Sex reversal triggers the rapid transition from genetic to temperature-dependent sex. Nature 2015, 523, 79–82. [Google Scholar] [CrossRef]
- Jensen, M.P.; Allen, C.D.; Eguchi, T.; Bell, I.P.; LaCasella, E.L.; Hilton, W.A.; Hof, C.A.; Dutton, P.H. Environmental Warming and Feminization of One of the Largest Sea Turtle Populations in the World. Curr. Biol. 2018, 28, 154–159.e4. [Google Scholar] [CrossRef] [Green Version]
- Ceballos, C.P.; Valenzuela, N. The Role of Sex-specific Plasticity in Shaping Sexual Dimorphism in a Long-lived Vertebrate, the Snapping Turtle Chelydra serpentina. Evol. Biol. 2011, 38, 163–181. [Google Scholar] [CrossRef]
- Ceballos, C.P.; Adams, D.C.; Iverson, J.B.; Valenzuela, N. Phylogenetic Patterns of Sexual Size Dimorphism in Turtles and Their Implications for Rensch’s Rule. Evol. Biol. 2012, 40, 194–208. [Google Scholar] [CrossRef]
- Ceballos, C.P.; Hernández, O.E.; Valenzuela, N. Divergent Sex-Specific Plasticity in Long-Lived Vertebrates with Contrasting Sexual Dimorphism. Evol. Biol. 2013, 41, 81–98. [Google Scholar] [CrossRef]
- Valenzuela, N. Sexual Development and the Evolution of Sex Determination. Sex. Dev. 2008, 2, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, N. Causes and consequences of evolutionary transitions in the level of phenotypic plasticity of reptilian sex de-termination. In Transitions Between Sexual Systems: Understanding the Evolution of Hermaphroditism and Dieocy, Hermaphroditism and Other Sexual Systems; Leonard, J.L., Ed.; Springer Nature: Cham, Switzerland, 2018; pp. 345–363. [Google Scholar]
- American Society of Ichthyologists and Herpetologists. Nature 1937, 139, 1014. [CrossRef] [Green Version]
- Van Der Heiden, A.M.; Briseno-Duenas, R.; Rios-Olmeda, D. A Simplified Method for Determining Sex in Hatchling Sea Turtles. Copeia 1985, 1985, 779. [Google Scholar] [CrossRef]
- Owens, D.W.; Hendrickson, J.R.; Lance, V.A.; Callard, I.P. A Technique for Determining Sex of Immature Chelonia mydas Using a Radioimmunoassay. Herpetologica. 1978, 34, 270–273. Available online: https://www.jstor.org/stable/3891551 (accessed on 1 December 2022).
- Rostal, D.C.; Grumbles, J.S.; Lance, V.A.; Spotila, J.R. Non-Lethal Sexing Techniques for Hatchling and Immature Desert Tortoises (Gopherus agassizii). Herpetol. Monogr. 1994, 8, 83. [Google Scholar] [CrossRef]
- Tezak, B.M.; Guthrie, K.; Wyneken, J. An Immunohistochemical Approach to Identify the Sex of Young Marine Turtles. Anat. Rec. 2017, 300, 1512–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuchling, G.; Kitimasak, W. Endoscopic sexing of juvenile softshell turtles, Amyda cartilaginea. Nat. Hist. Chulalongkorn Univ. 2009, 9, 91–93. [Google Scholar]
- Miller, W.J.; Wagner, F.H. Sexing Mature Columbiformes by Cloacal Characters. Ornithology 1955, 72, 279–285. [Google Scholar] [CrossRef]
- Valenzuela, N.; Adams, D.C.; Bowden, R.M.; Gauger, A.C. Geometric Morphometric Sex Estimation for Hatchling Turtles: A Powerful Alternative for Detecting Subtle Sexual Shape Dimorphism. Copeia 2004, 2004, 735–742. [Google Scholar] [CrossRef]
- Sönmez, B.; Turan, C.; Özdilek, Ş.Y.; Turan, F. Sex determination of green sea turtle (Chelonia mydas) hatchlings on the bases of morphological characters. J. Black Sea/Mediterr Environ. 2016, 22, 93–102. [Google Scholar]
- Boone, J.L.; Holt, E.A. Sexing Young, Free-ranging Desert Tortoises (Gopherus agassizii) Using External Morphology. Chel. Conserv. Biol. 2001, 4, 28–33. [Google Scholar]
- Michel-Morfin, J.E.; Munoz, V.M.G.; Rodriguez, C.N. Morphometric Model for Sex Assessment in Hatchling Olive Ridley Sea Turtles. Chelon. Conserv. Biol. 2001, 4, 53–58. [Google Scholar]
- McKnight, D.T.; Howell, H.J.; Hollender, E.C.; Ligon, D.B. Good vibrations: A novel method for sexing turtles. Acta Herpetol. 2017, 13, 13–19. [Google Scholar]
- Rodrigues, J.; Soares, D.; Silva, J. Sexing freshwater turtles: Penile eversion in Phrynops tuberosus (Testudines: Chelidae). Acta Herpetol. 2014, 9, 259–263. [Google Scholar] [CrossRef]
- Literman, R.; Badenhorst, D.; Valenzuela, N. qPCR-based molecular sexing by copy number variation in rRNA genes and its utility for sex identification in soft-shell turtles. Methods Ecol. Evol. 2014, 5, 872–880. [Google Scholar] [CrossRef]
- Literman, R.; Radhakrishnan, S.; Tamplin, J.; Burke, R.; Dresser, C.; Valenzuela, N. Development of sexing primers in Glyptemys insculpta and Apalone spinifera turtles uncovers an XX/XY sex-determining system in the critically-endangered bog turtle Glyptemys muhlenbergii. Conserv. Genet. Resour. 2017, 9, 651–658. [Google Scholar] [CrossRef]
- Rovatsos, M.; Praschag, P.; Fritz, U.; Kratochvšl, L. Stable Cretaceous sex chromosomes enable molecular sexing in softshell turtles (Testudines: Trionychidae). Sci. Rep. 2017, 7, srep42150. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, N.; Badenhorst, D.; Montiel, E.E.; Literman, R. Molecular Cytogenetic Search for Cryptic Sex Chromosomes in Painted Turtles Chrysemys picta. Cytogenet. Genome Res. 2014, 144, 39–46. [Google Scholar] [CrossRef]
- Tezak, B.; Sifuentes-Romero, I.; Milton, S.; Wyneken, J. Identifying Sex of Neonate Turtles with Temperature-dependent Sex Determination via Small Blood Samples. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurdyukov, S.; Bullock, M. DNA Methylation Analysis: Choosing the Right Method. Biology 2016, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Caetano, L.; Gennaro, F.; Coelho, K.; Araújo, F.; Vila, R.; Araújo, A.; Bernardo, A.D.M.; Marcondes, C.; Lopes, S.C.D.S.; Ramos, E. Differential expression of the MHM region and of sex-determining-related genes during gonadal development in chicken embryos. Genet. Mol. Res. 2014, 13, 838–849. [Google Scholar] [CrossRef] [PubMed]
- Readel, A.M.; Dreslik, M.J.; Warner, J.K.; Banning, W.J.; Phillips, C.A. A Quantitative Method for Sex Identification in Emydid Turtles Using Secondary Sexual Characters. Copeia 2008, 2008, 643–647. [Google Scholar] [CrossRef]
- Valenzuela, N. Egg Incubation and Collection of Painted Turtle Embryos. Cold Spring Harb. Protoc. 2009, 2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cagle, F.R. A System of Marking Turtles for Future Identification. Copeia 1939, 1939, 170. [Google Scholar] [CrossRef]
- Collyer, M.L.; Adams, D.C. RRPP: An r package for fitting linear models to high-dimensional data using residual randomization. Methods Ecol. Evol. 2018, 9, 1772–1779. [Google Scholar] [CrossRef] [Green Version]
- Epigentek. EpiGentek MethylFlash ™ Global DNA Methylation. 2019, pp. 1–13. Available online: https://www.epigentek.com/docs/P-1030.pdf (accessed on 1 December 2022).
- Canellas, P.F.; Karu, A.E. Statistical package for analysis of competition ELISA results. J. Immunol. Methods 1981, 47, 375–385. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Babraham Bioinformatics. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 1 December 2022).
- Krueger, F.; Trim galore. A Wrapper Tool Around Cutadapt and FastQC to Consistently Apply Quality and Adapter Trimming to FastQ Files. Babraham Bioinformatics. 2017. Available online: https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/ (accessed on 1 December 2022).
- Badenhorst, D.; Hillier, L.W.; Literman, R.; Montiel, E.E.; Radhakrishnan, S.; Shen, Y.; Minx, P.; Janes, D.E.; Warren, W.C.; Edwards, S.V.; et al. Physical Mapping and Refinement of the Painted Turtle Genome (Chrysemys picta) Inform Amniote Genome Evolution and Challenge Turtle-Bird Chromosomal Conservation. Genome Biol. Evol. 2015, 7, 2038–2050. [Google Scholar] [CrossRef] [Green Version]
- Krueger, F.; Andrews, S.R. Bismark: A flexible aligner and methylation caller for Bisulfite-Seq applications. Bioinformatics 2011, 27, 1571–1572. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allaire, J.J. RStudio: Integrated Development Environment for R. 2012. Available online: https://www.r-project.org/conferences/useR-2011/abstracts/180111-allairejj.pdf (accessed on 1 December 2022).
- Akalin, A.; Kormaksson, M.; Li, S.; Garrett-Bakelman, F.E.; Figueroa, M.E.; Melnick, A.; E Mason, C. methylKit: A comprehensive R package for the analysis of genome-wide DNA methylation profiles. Genome Biol. 2012, 13, R87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akalin, A.; Franke, V.; Vlahoviček, K.; Mason, C.E.; Schübeler, D. Genomation: A toolkit to summarize, annotate and visualize genomic intervals. Bioinformatics 2014, 31, 1127–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhou, J.; White, K.P. RNA-seq differential expression studies: More sequence or more replication? Bioinformatics 2013, 30, 301–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Tran, H.; Porter, J.; Sun, M.; Xie, H.; Zhang, L. Objective and Comprehensive Evaluation of Bisulfite Short Read Mapping Tools. Adv. Bioinform. 2014, 2014, 1–11. [Google Scholar] [CrossRef]
- Gu, H.; Bock, C.; Mikkelsen, T.S.; Jäger, N.; Smith, Z.D.; Tomazou, E.; Gnirke, A.; Lander, E.S.; Meissner, A. Genome-scale DNA methylation mapping of clinical samples at single-nucleotide resolution. Nat. Methods 2010, 7, 133–136. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A.; Ozaki, Y.; A Stockwell, P.; Horsfield, J.A.; Morison, I.M.; Nakagawa, S. Mapping the zebrafish brain methylome using reduced representation bisulfite sequencing. Epigenetics 2013, 8, 979–989. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Hart, S.N.; Bamlet, W.R.; Moore, R.M.; Nandakumar, K.; Eckloff, B.W.; Lee, Y.K.; Petersen, G.M.; McWilliams, R.R.; Couch, F.J. Prevalence of pathogenic mutations in cancer predisposition genes among pancreatic cancer patients. Cancer Epidemiol. Biomark. Prev. 2016, 25, 207–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraga, M.F.; Esteller, M. DNA Methylation: A Profile of Methods and Applications. BioTechniques 2002, 33, 632–649. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Choi, J.-Y.; Lee, K.-M.; Sung, H.; Park, S.K.; Oze, I.; Pan, K.-F.; You, W.-C.; Chen, Y.-X.; Fang, J.-Y.; et al. DNA Methylation in Peripheral Blood: A Potential Biomarker for Cancer Molecular Epidemiology. J. Epidemiol. 2012, 22, 384–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Chen, A.C.-Q.; Manev, H. Enzymatic Regional Methylation Assay for Determination of CpG Methylation Density. Anal. Chem. 2004, 76, 6829–6832. [Google Scholar] [CrossRef] [PubMed]
- Galm, O.; Rountree, M.R.; Bachman, K.E.; Jair, K.-W.; Baylin, S.B.; Herman, J.G. Enzymatic Regional Methylation Assay: A Novel Method to Quantify Regional CpG Methylation Density. Genome Res. 2001, 12, 153–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, A. Parle-McDermott, A. DNA Methylation: A Timeline of Methods and Applications. Front. Genet. 2011, 2, 74. [Google Scholar] [CrossRef]
- Kunde-Ramamoorthy, G.; Coarfa, C.; Laritsky, E.; Kessler, N.; Harris, R.; Xu, M.; Chen, R.; Shen, L.; Milosavljevic, A.; Waterland, R.A. Comparison and quantitative verification of mapping algorithms for whole-genome bisulfite sequencing. Nucleic Acids Res. 2014, 42, e43. [Google Scholar] [CrossRef]
- Smallwood, S.A.; Lee, H.J.; Angermueller, C.; Krueger, F.; Saadeh, H.; Peat, J.; Andrews, S.R.; Stegle, O.; Reik, W.; Kelsey, G. Single-cell genome-wide bisulfite sequencing for assessing epigenetic heterogeneity. Nat. Methods 2014, 11, 817–820. [Google Scholar] [CrossRef]
- Hong, S.R.; Shin, K.-J. Bisulfite-Converted DNA Quantity Evaluation: A Multiplex Quantitative Real-Time PCR System for Evaluation of Bisulfite Conversion. Front. Genet. 2021, 12, 618955. [Google Scholar] [CrossRef]
- Lance, V.; Valenzuela, N.; Hildebrand, P. A hormonal method to determine sex of hatchling giant river turtles, Podocnemis expansa: Application to endangered species. Am. Zool. 1992, 270, 16A. [Google Scholar]
- Gómez-Saldarriaga, C.; Valenzuela, N.; Ceballos, C.P. Effects of Incubation Temperature on Sex Determination in the Endangered Magdalena River Turtle, Podocnemis lewyana. Chelonian Conserv. Biol. 2016, 15, 43–53. [Google Scholar] [CrossRef]
- Radhakrishnan, S.; Literman, R.; Neuwald, J.; Severin, A.; Valenzuela, N. Transcriptomic responses to environmental temperature by turtles with temperature-dependent and genotypic sex determination assessed by RNAseq inform the genetic architecture of embryonic gonadal development. PLoS ONE 2017, 12, e0172044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radhakrishnan, S.; Literman, R.; Neuwald, J.L.; Valenzuela, N. Thermal Response of Epigenetic Genes Informs Turtle Sex Determination with and without Sex Chromosomes. Sex. Dev. 2018, 12, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Mizoguchi, B.; Valenzuela, N. Alternative splicing and thermosensitive expression of Dmrt1 during urogenital development in the painted turtle, Chrysemys picta. PeerJ 2020, 8, e8639. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, N.; Neuwald, J.L.; Literman, R. Transcriptional evolution underlying vertebrate sexual development. Dev. Dyn. 2012, 242, 307–319. [Google Scholar] [CrossRef]
- Valenzuela, N. Relic Thermosensitive Gene Expression In A Turtle With Genotypic Sex Determination. Evolution 2007, 62, 234–240. [Google Scholar] [CrossRef]
- Valenzuela, N.; LeClere, A.; Shikano, T. Comparative gene expression of steroidogenic factor 1 in Chrysemys picta and Apalone mutica turtles with temperature-dependent and genotypic sex determination. Evol. Dev. 2006, 8, 424–432. [Google Scholar] [CrossRef]
- Berry, J.F.; Shine, R. Sexual size dimorphism and sexual selection in turtles (order testudines). Oecologia 1980, 44, 185–191. [Google Scholar] [CrossRef]
- Ernst, C.H. Sexual Cycles And Maturity Of The Turtle, Chrysemys Picta. Biol. Bull. 1971, 140, 191–200. [Google Scholar] [CrossRef]
- Xiao, F.-H.; Chen, X.-Q.; He, Y.; Kong, Q.-P. Accelerated DNA methylation changes in middle-aged men define sexual dimorphism in human lifespans. Clin. Epigenetics 2018, 10, 133. [Google Scholar] [CrossRef] [Green Version]
- Weidner, C.I.; Lin, Q.; Koch, C.M.; Eisele, L.; Beier, F.; Ziegler, P.; Bauerschlag, D.O.; Jöckel, K.-H.; Erbel, R.; Mühleisen, T.W.; et al. Aging of blood can be tracked by DNA methylation changes at just three CpG sites. Genome Biol. 2014, 15, R24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lofton-Day, C.; Model, F.; DeVos, T.; Tetzner, R.; Distler, J.; Schuster, M.; Song, X.; Lesche, R.; Liebenberg, V.; Ebert, M.; et al. DNA Methylation Biomarkers for Blood-Based Colorectal Cancer Screening. Clin. Chem. 2008, 54, 414–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsit, C.J.; Koestler, D.C.; Christensen, B.C.; Karagas, M.R.; Houseman, E.A.; Kelsey, K.T. DNA Methylation Array Analysis Identifies Profiles of Blood-Derived DNA Methylation Associated With Bladder Cancer. J. Clin. Oncol. 2011, 29, 1133–1139. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| ANOVA—Methylation Percentage (Full Factorial Model) | ||||
|---|---|---|---|---|
| Sum of Squares | Mean Square | F | p-Value | |
| Temperature | 14.4 | 14.376 | 2.62 | 0.138 |
| Sex | 0.0 | 0.015 | 0.002 | 0.961 |
| Temperature:Sex | 8.4 | 8.39 | 1.320 | 0.255 |
| Residuals | 356 | 6.357 | ||
| ANOVA—methylation percentage (reduced model) | ||||
| Sum of Squares | Mean Square | F | p-value | |
| Temperature | 14.4 | 14.376 | 2.249 | 0.139 |
| Sex | 0.0 | 0.015 | 0.002 | 0.961 |
| Residuals | 364.4 | 6.393 | ||
| ANOVA—methylation percentage by sex | ||||
| Sum of Squares | Mean Square | F | p-value | |
| Sex | 9.5 | 9.511 | 1.494 | 0.227 |
| Residuals | 369.3 | 6.367 | ||
| ANOVA—methylation percentage by temperature | ||||
| Sum of Squares | Mean Square | F | p-value | |
| Temperature | 14.4 | 14.376 | 2.288 | 0.136 |
| Residuals | 364.4 | 6.283 | ||
| ANOVA—absorbance (full factorial model) | ||||
| Sum of Squares | Mean Square | F | p-value | |
| Temperature | 4.26 | 4.262 | 5.509 | 0.5942 |
| Sex | 0.22 | 0.222 | 0.287 | 2.33e−0.6 |
| Temperature:Sex | 21.41 | 21.414 | 27.679 | |
| Residuals | 43.33 | 0.774 | ||
| Pairwise comparisons of absorbance. Mean absorbance per temperature by sex combination (diagonal, underlined), mean squares (below diagonal, italics), p-values (above diagonal, significant values are denoted in bold) | ||||
| 26 °C—Male | 28 °C—Male | 31 °C—Female | 28 °C—Female | |
| 26 °C—Male | 0.5999 | 0.0008 | 0.126 | 0.005 |
| 28 °C—Male | 32.56 | −1.0218 | 0.035 | 0.771 |
| 31 °C—Female | 7.507 | 10.722 | −0.2172 | 0.067 |
| 28 °C—Female | 24.402 | 0.084 | 8.803 | −0.9854 |
| Regions | |||
|---|---|---|---|
| CPI 3.0.3. Scaffold | Start Position | End Position | Gene |
| NW_007359905.1 | 3,604,500 | 3,605,000 | Intergenic |
| NC_024218.1 | 55,635,611 | 55,636,011 | Intergenic |
| NW_007281443.1 | 1,473,502 | 1,474,001 | FAM170B |
| Sites | |||
| CPI 3.0.3. Scaffold | Position | Gene | |
| NC_024218.1 | 18,209,143 | CASQ2 | |
| NC_024220.1 | 23,784,593 | FOSL2 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mizoguchi, B.A.; Valenzuela, N. A Cautionary Tale of Sexing by Methylation: Hybrid Bisulfite-Conversion Sequencing of Immunoprecipitated Methylated DNA in Chrysemys picta Turtles with Temperature-Dependent Sex Determination Reveals Contrasting Patterns of Somatic and Gonadal Methylation, but No Unobtrusive Sex Diagnostic. Animals 2023, 13, 117. https://doi.org/10.3390/ani13010117
Mizoguchi BA, Valenzuela N. A Cautionary Tale of Sexing by Methylation: Hybrid Bisulfite-Conversion Sequencing of Immunoprecipitated Methylated DNA in Chrysemys picta Turtles with Temperature-Dependent Sex Determination Reveals Contrasting Patterns of Somatic and Gonadal Methylation, but No Unobtrusive Sex Diagnostic. Animals. 2023; 13(1):117. https://doi.org/10.3390/ani13010117
Chicago/Turabian StyleMizoguchi, Beatriz A., and Nicole Valenzuela. 2023. "A Cautionary Tale of Sexing by Methylation: Hybrid Bisulfite-Conversion Sequencing of Immunoprecipitated Methylated DNA in Chrysemys picta Turtles with Temperature-Dependent Sex Determination Reveals Contrasting Patterns of Somatic and Gonadal Methylation, but No Unobtrusive Sex Diagnostic" Animals 13, no. 1: 117. https://doi.org/10.3390/ani13010117