1. Introduction
Wild bird species such as the European starling (
Sturnus vulgaris) provide valuable animal models for studying the biology of foraging behaviour and the regulation of fat reserves and body mass [
1,
2]. A well-developed theoretical literature in evolutionary ecology argues that birds should adaptively regulate their fat reserves to optimize the trade-off between the increased starvation risk caused by having too little fat and the increased predation risk caused by having too much [
3,
4,
5,
6,
7]. These optimality models predict how birds should strategically regulate their body mass and the shape of their mass gain trajectory over the day in response to environmental challenges including perceived predation risk and unpredictable access to food. Laboratory experiments on birds have been central in testing these models of adaptive mass regulation. For example, numerous experimental studies on starlings have confirmed the theoretical prediction that limited and unpredictable access to food should cause birds to gain mass as insurance against starvation [
2,
8,
9,
10,
11]. This ‘insurance hypothesis’ has recently been suggested as a possible evolutionary explanation for the increased odds of overweight observed in food insecure humans, suggesting that studies of birds could provide new insights into the environmental causes of the human obesity epidemic [
12,
13,
14,
15].
Despite the above achievements, progress in understanding the behavioural and physiological mechanisms underlying body mass regulation in birds has been limited by the practical difficulties of obtaining high quality individual-level data on body mass, foraging behaviour, and physiology from birds housed in ecologically valid conditions in the laboratory. Moreover, the common practice of individually housing birds in small cages that facilitate behavioural measurements and frequent capture for manual weighing is likely to be chronically stressful for social species such as the starling. We therefore set out to demonstrate that high quality individual-level data on foraging behaviour and body mass, which are necessary for testing mechanistic hypotheses, can be obtained from group-housed starlings living in large aviaries that simultaneously provide greater ecological validity and improved animal welfare compared with previous methods.
Laboratory studies in birds and mammals have identified a number of methodological variables that are likely to affect the ecological validity of studies of foraging behaviour and mass regulation. The first of these is cage size, which is potentially important because it affects the distances that animals can walk, run, or fly and hence their potential energy expenditure through physical activity. While it is generally assumed that increased body mass under conditions of unpredictable food results from increased energy intake [
16,
17,
18], reduced energy expenditure is also likely to be important; studies of both starlings and zebra finches (
Taeniopygia guttata) showed that birds respond to unpredictable food by reducing their physical activity [
2,
19]. Cage furnishing also affects the range of behaviour patterns that animals can perform in the laboratory and hence their energy expenditure. In rats, the offspring of mothers that are undernourished during pregnancy become obese when housed in standard laboratory cages without the opportunity for exercise, but when provided with daily access to a running wheel, they engaged in voluntary exercise and maintained normal body mass [
20,
21]. Thus, housing animals in small barren cages might constrain them to be sedentary and remove a potentially important mechanism that is present in the natural environment for regulating energy balance.
Housing social animals in groups is likely to be important for body mass for two reasons. First, group housing permits social behaviour, which can form an important contribution to an individual’s time budget and contribute to energy expenditure. In songbirds such as the starling, song is primarily used in intra-specific communication and is energetically costly [
22]. Second, group housing can affect body mass by increasing competition for food. Dominant individuals in a group can displace subordinate animals from a monopolizable food source, creating inequalities within the group in the predictability of food access. In starlings, as predicted by the insurance hypothesis, the effects of unpredictable food access on body mass are greater when there is more competition between birds for access to feeders [
2]. Furthermore, experimentally increasing a subordinate bird’s rank in a group (by removing the most dominant individual) causes a decrease in its body mass [
23]. Thus, housing animals individually removes opportunities for energetically expensive social behaviour, and also removes competition for food, reducing unpredictability in food and thereby eliminating an important source of individual variability in body mass that is present in the natural environment.
All experimental studies of foraging behaviour can be classified as either open or closed-economy designs [
24]. In an open economy experiment, a subject is placed in the experimental manipulation for part of the day and its total daily food intake is determined by the amount of food provided by the experimenter outside of this period. For example, in many psychological studies, subjects are maintained at a fixed percentage of their free-feeding mass, regardless of the amount of food eaten in the experimental session. In contrast, in a closed economy experiment, the manipulation is typically in place for the entire day, and the subject’s response to the manipulation completely determines its daily intake. Theoretical models show that the optimal proportion of time a subject should spend foraging depends on the rate at which it can obtain food, but that the direction of this relationship depends on whether the subject is in open or closed economy [
25]. Thus, the type of economy potentially influences the effects that will be observed in experiments. Closed economy experiments provide a better simulation of the situation present in natural environments.
In summary, to perform ecologically valid studies of body mass regulation and understand the behavioural, psychological, and physiological mechanisms underlying this, we need experimental laboratory studies that incorporate the following features. First, subjects must be housed in enclosures that are large enough and contain the furnishings necessary to allow performance of a natural range of behaviour. Second, subjects of social species should be housed in groups that permit social interactions and the formation of dominance hierarchies. Third, studies must be performed in closed economy, whereby the subjects’ adjustments to the experimental conditions they are placed under directly determine their daily food intake. Finally, human intervention, especially catching and restraint, which are stressful and likely to be perceived by subjects as predation attempts, should be limited as far as possible.
Here, we describe the social foraging system (SFS), a novel technology that allows remotely operated experimental manipulation of food access and permits continuous collection of individual-level data on operant foraging behaviour. We present observational data on the patterns of foraging behaviour and mass gain that can be recorded using the SFS as well as the methods that we have developed to train the birds.
The SFS is based around smart feeding stations that identify individual birds with radio frequency identification (RFID), weigh them with an integrated electronic balance, record foraging behaviour using pecks on an illuminable pecking key, and control food access using a retractable food hopper. The feeding stations are connected to a computer that controls the operant schedule in place on the pecking key and access to the food hopper and collects data on bird identities, body masses, and foraging behaviour continuously between dawn and dusk each day (starlings do not feed at night).
A potential challenge raised by the SFS is the training of the birds. In the past, we have conducted operant training in individual cages where individual experience can be controlled and progression based on individual performance criteria (e.g., [
26]). Starlings are social foragers and we speculated that group training might increase the willingness of more neophobic individuals to approach the SFS and feed from the hopper when they saw bolder birds foraging there. However, we were also concerned that during operant training, some birds might learn to scrounge food rewards earned by other individuals, rather than learning to peck the keys for food themselves. Over a series of pilot studies not described here, we developed a protocol for training starlings to forage from the SFS in groups of three birds. Six of the birds used in the current study were involved in these earlier trials and were trained to forage from the SFS prior to the start of the current study. However, the other six birds were experimentally naïve and we used this study as an opportunity to test a new social training protocol for the SFS.
We start by describing the acquisition of operant key pecking in naïve group-housed birds. Our aim was to discover whether starlings can learn key pecking without the need for social isolation. Following successful operant training, we report the observational foraging and body mass data from two groups of six birds maintained on a fixed-ratio operant schedule under closed economy for 11 consecutive days. Our aims were to describe the patterns of foraging behaviour and mass gain within the day within birds; to establish the repeatability of these measurements between days; and, to explore how variation in foraging behaviour is related to variation in body mass both within and between birds. We discuss the strengths and limitations of the SFS, focusing on both the quality of the scientific data obtained and the likely welfare of the birds.
2. Materials and Methods
2.1. Ethical Statement
The study adhered to ASAB/ABS guidelines for the use of animals in research. Birds were taken from the wild under Natural England permit 20121066 and the research was completed under UK Home Office licence PPL P038AB1D3 with the approval of the Animal Welfare and Ethical Review Body at Newcastle University. Reporting followed the ARRIVE 2.0 guidelines [
27].
2.2. Subjects and Basic Husbandry
Subjects were 12 adult European starlings (
Sturnus vulgaris), six males and six females, originally caught from the wild in March 2019 at Seaton Sluice, Northumberland, UK. Birds were sexed using bill and iris colour [
28]. Tarsus length was measured twice for the right and left leg using digital callipers and a mean tarsus length calculated for each bird as a measure of skeletal size [
29].
Prior to the study described in the current paper, which took place in March 2020, the birds were group-housed in a single indoor ‘home’ aviary (280 cm wide × 300 cm deep × 255 cm high). The aviary was without windows and was artificially lit and ventilated. The temperature and humidity were maintained at ~18–20 °C and ~40% respectively. The lights were on between 0800–1715, providing 9.25 h daylight. Between 0800 and 0815, the lights gradually increased in intensity to simulate dawn and between 1700–1715, the lights dimmed gradually to simulate dusk and allow birds to settle for the night. The aviary was furnished with rope perches, wood chippings on the floor, a water bath, and a drinker providing ad libitum clean water supplemented with vitamins. Birds were fed ad libitum on commercial poultry starter crumb (Special Diets Services Poultry Starter; henceforth ‘crumb’)—a homogeneous complete diet for starlings—supplemented with softbill mix (Orlux Universal Softbill Food), fresh fruit, and live mealworms.
For the study described in the current paper, the birds were caught and transferred to experimental aviaries described below. After completion of the data collection, the birds were retained in the experimental aviaries for an experimental study not reported here. In April 2020, following completion of the latter experiment and inspection by a veterinary surgeon, the birds were released to the wild at the site of original capture.
2.3. Experimental Aviaries and the Social Foraging System
Experimental aviaries were identical to the home aviary described above with the exception that each aviary was equipped with one SFS station for every three birds present in the aviary. The SFS was built to our specifications by Campden Instruments, Loughborough, UK. A single SFS station (
Figure 1) comprised a motorized retractable food hopper filled with crumb and an illuminable pecking key, both of which could only be accessed via a wooden perch designed to accommodate a single bird. The perch was mounted at the apex of a smooth plastic pyramid designed to prevent other birds from perching on the station and protect the balance and RFID aerial that were located beneath it. The pyramid was mounted on an electronic balance that measured to a resolution of 0.01 g. The RFID aerial was tuned to read microchips that were glued to plastic leg rings worn by a starling standing on the perch. The SFS stations were connected to a single computer in an adjacent room running Whisker experimental control software [
30], a custom-written programme (Starfeeder) that managed the RFID and mass data, and additional custom-written Whisker ‘client’ programmes specific to different phases of the study. The computer controlled the operant schedule in place on each station and collected continuous data on the identity and masses of birds visiting each station, plus any key pecks and food rewards delivered. The data files written by the Whisker client programmes could be accessed in real-time by custom-written R scripts [
31] that produced summaries of the birds’ masses and key pecking behaviour for the current day. These latter summaries were checked a minimum of three times daily and were used for welfare monitoring purposes.
Husbandry in the experimental aviaries took place between 1600 and 1700 daily throughout the study. Birds were fed on crumb that was available either from the SFS hopper or from ad libitum bowls that were provided following operant training sessions. The diet was supplemented with four live mealworms per bird given during daily husbandry and supplied in spatially separated bowls to prevent one bird from monopolizing the worms.
2.4. Welfare Monitoring
A welfare monitoring protocol was designed that takes advantage of the mass data provided by the SFS and hence avoids unnecessary catching of the birds (
Supplementary Materials Document: Welfare monitoring in European starlings). Catching for manual weighing was only required by the protocol in the period of habituation, before birds started perching on the SFS and recording automatic body masses.
2.5. Study Design
The study was observational with no experimental manipulation. Data on foraging behaviour and body mass are described from two groups of six birds housed in separate experimental aviaries and maintained under closed economy for 11 consecutive days. The design is summarized in
Figure 2.
2.6. Operant Training
Three days prior to being transferred to the experimental aviaries for the start of operant training, the diet of the birds in the home aviary was restricted to ad libitum crumb in order to habituate them to the diet that would be available from the SFS. At ~1200 on the third day, 12 birds were caught from the home aviary, manually weighed, and fitted with two coloured plastic leg rings, each of which had a unique microchip attached. Birds wore two microchips to guard against identification failure in the event that one microchip fell off, broke, or was not read due to poor alignment with the aerial in the SFS. Six of the birds had been trained to forage from the SFS ~6 months previously and were released directly into a single experimental aviary equipped with two SFS stations running the continuous foraging operant schedule (see below). The other six birds were experimentally-naïve and required operant training.
Training was conducted in groups of three to facilitate monitoring of individual performance during the acquisition of key pecking. Three birds were released into two experimental aviaries (three males in one and three females in the other), each equipped with a single SFS station. The first phase of operant training was to habituate the birds to the SFS stations and to feeding from the SFS hopper. Throughout this phase, crumb was provided ad libitum by permanently raising the hoppers of the SFS stations between 0800–1645. Birds were initially encouraged to approach the SFS stations by placing two bowls of crumb and two bowls of mealworms (highly attractive to starlings) on the base of each station. Feeding was monitored by watching the birds via a live CCTV camera and weighing the bowls and SFS hopper at the end of each day. When the birds started feeding from the bowls, they were removed, and dried mealworms were manually placed in the SFS food hopper entrance to attract the birds to start feeding from the hopper. When the birds started feeding from the hopper, the dried mealworms were discontinued, and the birds were henceforth restricted to feeding on the crumb in the hopper. Throughout habituation, the number of stable masses recorded per bird each day was used to monitor the foraging effort and body mass of each bird and the daily decrease in the mass of the food hopper to monitor crumb consumption by each trio of birds. Until individual birds started perching on the balance, they were caught and manually weighed every second day. As soon as all the birds in an aviary were perching on the SFS and feeding from the hopper, the aviary progressed to the next phase of training.
The second phase of operant training was to teach the birds to peck the lit key on the SFS to raise the hopper and hence obtain access to food. During this phase, a daily operant training session ran from 0830 until ~1400 and was followed by the provision of bowls of ad libitum crumb placed on the base of the SFS between the end of the session and 1645 when birds were food deprived until the start of the session the following morning. The SFS hoppers remained in the inaccessible retracted position outside of the training sessions.
Operant training began with daily sessions of auto-shaping, whereby illumination of the pecking key on the SFS for 15 s predicted unconditional raising of the food hopper for 15 s, starting when the key light extinguished, followed by a 200-s inter-trial interval (ITI) during which the hopper was retracted and the key light was unlit. The aim of this schedule was to set up a Pavlovian association between the lit key and food reward. The acquisition of this association typically results in a conditioned response, whereby the birds spontaneously start to direct appetitive pecks at the lit key (auto-shaping). A peck to the lit key was reinforced by immediate hopper raising, thus additionally establishing an instrumental association between pecking the lit key, and immediate food reward. Daily sessions terminated after 90 trials or 6 h. This unconditional auto-shaping training continued daily until all the birds in an aviary had started pecking the lit key—each bird was required to make a minimum of three pecks in a session before the entire aviary could progress to the next phase of training. When this criterion was met, hopper raising was made conditional on a bird pecking the illuminated key. The stimulus time was maintained at 15 s, but the feeding time was reduced to 5 s and the ITI to 66 s. The shorter feeding time was designed to reduce the potential for birds to scrounge food produced by others, which might impair them from learning the association between pecking and food. The session terminated after 270 trials or 6 h. This conditional training continued daily until all the birds in an aviary were pecking the lit key a minimum of three times in a session. When this criterion was met, the aviary progressed to continuous foraging (see below) and as soon as both aviaries of naïve birds had reached this point, the six trained birds were united in a single experimental aviary equipped with two SFS stations running the continuous foraging schedule.
2.7. Continuous Foraging under Closed Economy
Following operant training, the birds were maintained in a closed economy, obtaining all of their daily food from the SFS (with the exception of the four mealworms given during husbandry). The default position of the food hopper was lowered so that food was unavailable and the key light on the SFS was illuminated to indicate that the SFS station was available for foraging. Food access was delivered on a ratio schedule, whereby a single peck at the lit key caused the key light to extinguish and the hopper to raise for 5 s (henceforth referred to as ‘a reinforcement’). At the end of the food access period, the hopper lowered and there was a 2-s ITI before the key re-lit, signalling the start of the next available trial. The two SFS stations in each aviary operated independently from one another, meaning that it was possible for two starlings in an aviary to forage simultaneously on different stations. The operant sessions began at dawn each day (0800) and ended at lights-off (1715). Thus, the maximum number of reinforcements available per day from a single SFS station was 4162 (calculated assuming that a bird always pecked the lit key one second after it illuminated). Continuous foraging continued for 11 consecutive days in each of the two experimental aviaries.
2.8. Outcome Variables
2.8.1. Operant Foraging Behaviour
In the current study, a single peck to a lit key always resulted in the food hopper raising for 5 s, meaning that the number of pecks equated to the number of reinforcements earned. The time of each peck was recorded, and the peck was attributed to the bird currently on the perch. Therefore, number of reinforcements earned was available at the individual bird level. Reinforcements earned was expressed as the rate × h−1.
2.8.2. Food Consumption
Total crumb consumption in each aviary was estimated daily by calculating the difference in the mass of the SFS food hoppers between the beginning and end of the day and subtracting any crumb collected in a spill tray located beneath each hopper. Therefore, consumption data were only available at the aviary level, but were expressed as g × bird−1·day−1 for ease of comparison between groups of different sizes.
2.8.3. Body Mass
In all experiments, body masses were recorded each day between lights-on and lights-off; each mass was assigned to the microchip recorded from the bird currently on the perch. Therefore, body mass was available at the individual bird level. The balances measured masses at a frequency of 6 Hz. A stable mass was recorded for a bird if the balance measured five consecutive masses of >50 g that were within a range of 5 g. These criteria were chosen to eliminate masses from birds that were perching incorrectly (e.g., by placing one foot on the food hopper), but to maximise the number of stable masses recorded from moving birds. Once a stable mass had been recorded, another stable mass could not be recorded until the balance had registered a mass <10 g, indicating that the current bird had left the perch. Balances were checked with a 100-g test mass a minimum of twice daily and calibrated if necessary. In order to control for build-up of guano on the perch over the day, balances were automatically zeroed regularly throughout the day when no bird was present on the perch. Prior to modelling the mass data, any masses greater than 120 g were removed on the grounds that such masses were at least 10 g above the maximum mass ever recorded for a starling in our laboratory and were thus likely to be the result of measurement error.
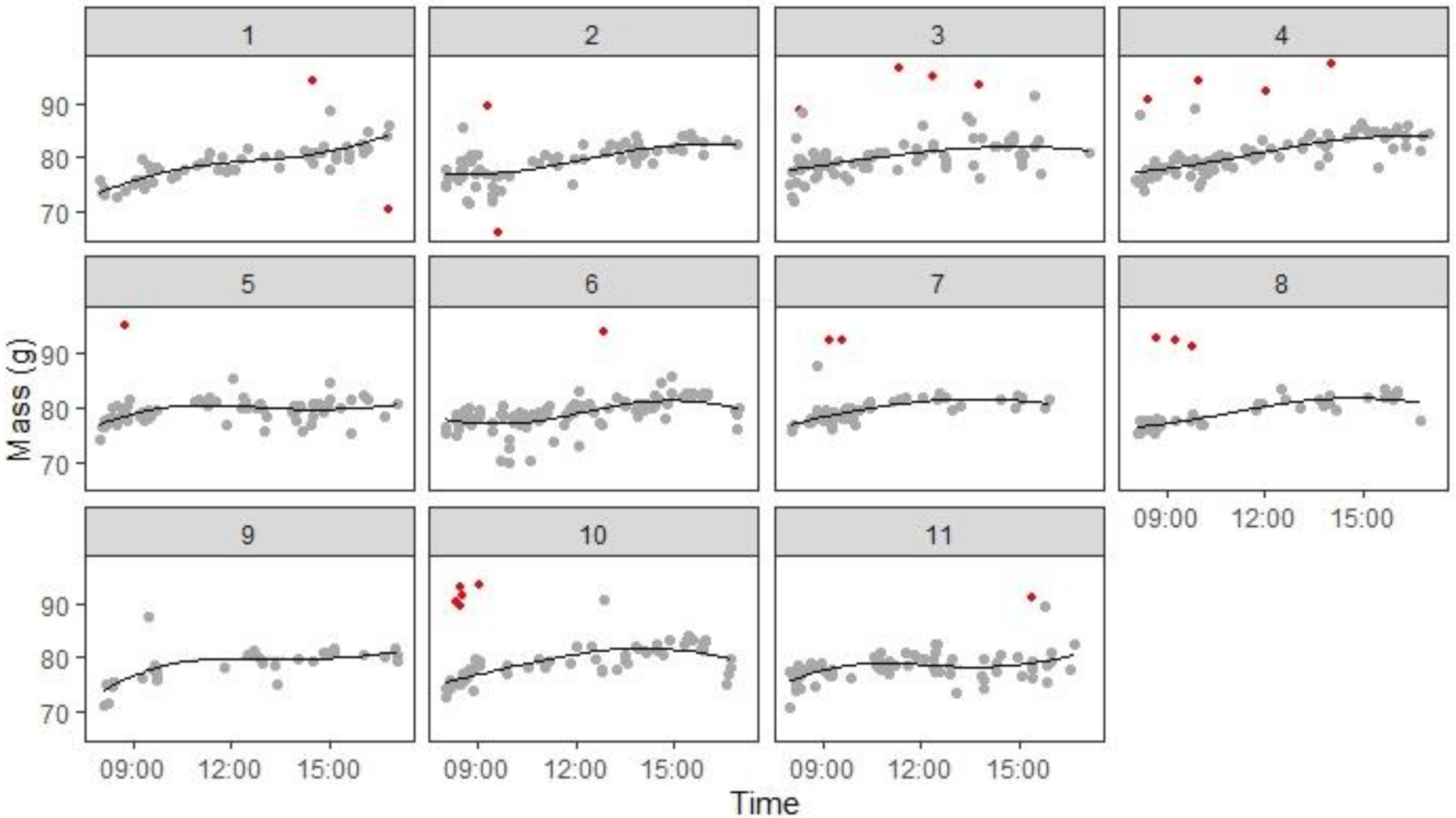
The raw masses showed a clear trend of mass increase over the day; the trajectory was typically non-linear, with mass gain generally being fastest early in the day, slowing down in the middle of the day and either peaking or speeding up again towards dusk. There is substantial random error in mass, due to the imprecision of the balances and movement of the birds whilst on the perch. Furthermore, masses were not always available at all times of every day (example data from one bird are shown in
Figure 3). To estimate comparable masses for each bird, we therefore modelled how individual body mass changed as a function of time of day. As long as a minimum of 10 masses were available for a bird on a given day, the masses were fitted with a regression line (on the choice of the model see below). To remove biologically implausible outliers, any masses >10 g from the fitted line were removed and a new cubic polynomial fitted to the remaining data (
Figure 3). This latter fit was used to estimate body mass at specific times of the day such as dawn or dusk. To avoid extrapolation beyond the measured data, a dawn or dusk mass was only estimated if there was a mass recording within 1 h of the estimate. Dawn was chosen as the time the lights came on (0800) and dusk as 1600, because birds often stopped foraging considerably before lights-off, reducing the number of masses available in the final hour of the day.
2.9. Inferential Statistics
Data were analysed using R version 3.5.1 [
31]. Due to the multi-level structure of the data, with individual observations nested within birds and birds nested within aviaries, we used general linear mixed models (GLMMs) for inferential statistics. GLMMs were fitted using restricted maximum likelihood estimation (REML) in the package ‘lme4′ [
32] and
p-values were calculated using Satterthwaite’s method in the package ‘lmerTest’ [
33]. We included a random intercept for bird in all GLMMs. A random intercept for aviary was also initially added to the models, but since in practice the aviary explained little or no variance, this random effect was dropped from the final version of the models reported. A fixed effect of sex was included in all models, because on average, male starlings are skeletally larger and heavier than females, meaning that there is reason to expect the effects of sex on between-subject differences in foraging behaviour and body mass. As expected, the males had longer tarsi than the females, but the difference was marginally non-significant (linear model: β
male ± se = 0.99 ± 0.47; F
1,10 = 4.41,
p = 0.062).
To assess the reliability of the measurements of reinforcements earned and body mass derived from the SFS across the 11 days of the study, we computed intra-class correlation coefficients (ICCs) and their 95% confidence intervals using the R package ‘psych’ [
34]. ICCs were based on a two-way random-effects model assuming single measurements and absolute-agreement.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}