
A Whole Genome Sequencing-Based Genome-Wide Association Study Reveals the Potential Associations of Teat Number in Qingping Pigs


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample and Sequencing
2.2. Phenotypic Data
2.3. Genotyping and Quality Control
2.4. SNP-Based Heritability
2.5. Principal Component Analysis
2.6. GWAS Using General Linear Model (GLM)
2.7. GWAS Using FarmCPU
2.8. Comparison with Known QTLs and Haplotype Analysis
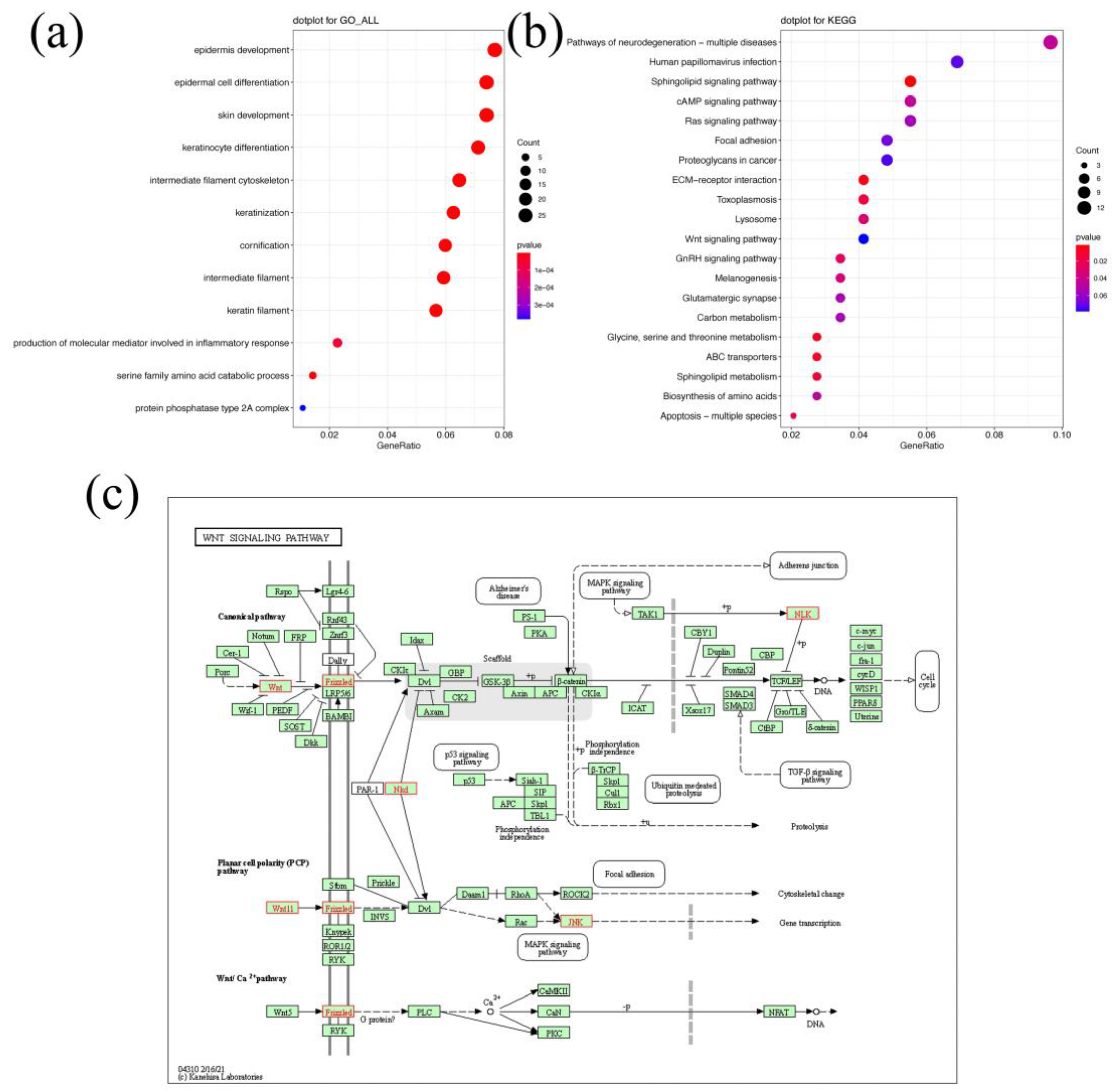
2.9. Annotation of Candidate Genes and Functional Enrichment Analysis
3. Results
3.1. Genotyping and Phenotypic Statistics
3.2. GLM GWAS for Teat Number-Related Traits
3.3. FarmCPU GWAS for ADIFF
3.4. Functional Enrichment of Candidate Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Y.; Pu, L.; Shi, L.; Gao, H.; Zhang, P.; Wang, L.; Zhao, F. Revealing New Candidate Genes for Teat Number Relevant Traits in Duroc Pigs Using Genome-Wide Association Studies. Animals 2021, 11, 806. [Google Scholar] [CrossRef] [PubMed]
- Pig Quantitative Trait Locus (Qtl) Database (Pig Qtldb). Available online: https://www.animalgenome.org/cgi-bin/QTLdb/SS/index (accessed on 27 December 2021).
- Arakawa, A.; Okumura, N.; Taniguchi, M.; Hayashi, T.; Hirose, K.; Fukawa, K.; Ito, T.; Matsumoto, T.; Uenishi, H.; Mikawa, S. Genome-Wide Association Qtl Mapping for Teat Number in a Purebred Population of Duroc Pigs. Anim. Genet. 2015, 46, 571–575. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, S.; Park, J.E.; Moon, S.H.; Choi, S.W.; Go, G.W.; Lim, D.; Kim, J.M. Genome-Wide Association Study and Genomic Predictions for Exterior Traits in Yorkshire Pigs1. J. Anim. Sci. 2019, 97, 2793–2802. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Atsuji, K.; Saito, N.; Okitsu, M.; Sato, S.; Komatsuda, A.; Mitsuhashi, T.; Nirasawa, K.; Hayashi, T.; Sugimoto, Y.; et al. Identification of Quantitative Trait Loci Affecting Corpora Lutea and Number of Teats in a Meishan X Duroc F2 Resource Population. J. Anim. Sci. 2006, 84, 2895–2901. [Google Scholar] [CrossRef]
- Hernandez, S.C.; Finlayson, H.A.; Ashworth, C.J.; Haley, C.S.; Archibald, A.L. A Genome-Wide Linkage Analysis for Reproductive Traits in F2 Large White X Meishan Cross Gilts. Anim. Genet. 2014, 45, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, Y.; Zhang, T.; Zhang, L.; Yan, H.; Liu, X.; Wang, L. Genotyping by Sequencing Reveals a New Locus for Pig Teat Number. Anim. Genet. 2017, 48, 470–472. [Google Scholar] [CrossRef]
- Zhou, L.; Zhao, W.; Fu, Y.; Fang, X.; Ren, S.; Ren, J. Genome-Wide Detection of Genetic Loci and Candidate Genes for Teat Number and Body Conformation Traits at Birth in Chinese Sushan Pigs. Anim. Genet. 2019, 50, 753–756. [Google Scholar] [CrossRef]
- Niu, N.; Wang, H.; Shi, G.; Liu, X.; Liu, H.; Liu, Q.; Yang, M.; Wang, L.; Zhang, L. Genome Scanning Reveals Novel Candidate Genes for Vertebral and Teat Number in the Beijing Black Pig. Anim. Genet. 2021, 52, 734–738. [Google Scholar] [CrossRef]
- Wada, Y.; Akita, T.; Awata, T.; Furukawa, T.; Sugai, N.; Inage, Y.; Ishii, K.; Ito, Y.; Kobayashi, E.; Kusumoto, H.; et al. Quantitative Trait Loci (Qtl) Analysis in a Meishan X Göttingen Cross Population. Anim. Genet. 2000, 31, 376–384. [Google Scholar] [CrossRef] [Green Version]
- Jinghu, Z.; Xiong, Y.; Zuo, B.; Lei, M.; Jiang, S.; Li, F.; Zheng, R.; Li, J.; Xu, D. Detection of Quantitative Trait Loci Associated with Several Internal Organ Traits and Teat Number Trait in a Pig Population. J. Genet. Genom. 2007, 34, 307–314. [Google Scholar]
- Bidanel, J.P.; Rosendo, A.; Iannuccelli, N.; Riquet, J.; Gilbert, H.; Caritez, J.C.; Billon, Y.; Amigues, Y.; Prunier, A.; Milan, D. Detection of Quantitative Trait Loci for Teat Number and Female Reproductive Traits in Meishan X Large White F2 Pigs. Animal 2008, 2, 813–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, C.; Wu, Z.; Ren, J.; Huang, Z.; Liu, D.; He, X.; Prakapenka, D.; Zhang, R.; Li, N.; Da, Y.; et al. Genome-Wide Association Study and Accuracy of Genomic Prediction for Teat Number in Duroc Pigs Using Genotyping-by-Sequencing. Genet. Sel. Evol. 2017, 49, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, Z.; Ding, R.; Peng, L.; Wu, J.; Ye, Y.; Zhou, S.; Wang, X.; Quan, J.; Zheng, E.; Cai, G.; et al. Genome-Wide Association Analyses Identify Known and Novel Loci for Teat Number in Duroc Pigs Using Single-Locus and Multi-Locus Models. BMC Genom. 2020, 21, 344. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Zhang, Z.; Yang, B.; Guo, Y.; Ai, H.; Long, Y.; Su, Y.; Cui, L.; Zhou, L.; Wang, X.; et al. Identification of Loci Affecting Teat Number by Genome-Wide Association Studies on Three Pig Populations. Asian-Australas. J. Anim. Sci. 2017, 30, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Rohrer, G.A.; Nonneman, D.J. Genetic Analysis of Teat Number in Pigs Reveals Some Developmental Pathways Independent of Vertebra Number and Several Loci Which Only Affect a Specific Side. Genet. Sel. Evol. 2017, 49, 4. [Google Scholar] [CrossRef] [Green Version]
- Moscatelli, G.; Dall’Olio, S.; Bovo, S.; Schiavo, G.; Kazemi, H.; Ribani, A.; Zambonelli, P.; Tinarelli, S.; Gallo, M.; Bertolini, F.; et al. Genome-Wide Association Studies for the Number of Teats and Teat Asymmetry Patterns in Large White Pigs. Anim. Genet. 2020, 51, 595–600. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Subgroup Genome Project Data Processing. The Sequence Alignment/Map Format and Samtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- DePristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A Framework for Variation Discovery and Genotyping Using Next-Generation DNA Sequencing Data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The Variant Call Format and Vcftools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-Generation Plink: Rising to the Challenge of Larger and Richer Datasets. Gigascience 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Browning, B.L.; Zhou, Y.; Browning, S.R. A One-Penny Imputed Genome from Next-Generation Reference Panels. Am. J. Hum. Genet. 2018, 103, 338–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Benyamin, B.; McEvoy, B.P.; Gordon, S.; Henders, A.K.; Nyholt, D.R.; Madden, P.A.; Heath, A.C.; Martin, N.G.; Montgomery, G.W.; et al. Common Snps Explain a Large Proportion of the Heritability for Human Height. Nat. Genet. 2010, 42, 565–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Team, R Core. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Yin, L.; Zhang, H.; Tang, Z.; Xu, J.; Yin, D.; Zhang, Z.; Yuan, X.; Zhu, M.; Zhao, S.; Li, X.; et al. Rmvp: A Memory-Efficient, Visualization-Enhanced, and Parallel-Accelerated Tool for Genome-Wide Association Study. Genom. Proteom. Bioinform. 2021, 19, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative Usage of Fixed and Random Effect Models for Powerful and Efficient Genome-Wide Association Studies. PLoS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Tian, F.; Pan, Y.; Buckler, E.S.; Zhang, Z. A Super Powerful Method for Genome Wide Association Study. PLoS ONE 2014, 9, e107684. [Google Scholar] [CrossRef]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and Visualization of Ld and Haplotype Maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [Green Version]
- Durinck, S.; Spellman, P.T.; Birney, E.; Huber, W. Mapping Identifiers for the Integration of Genomic Datasets with the R/Bioconductor Package Biomart. Nat. Protoc. 2009, 4, 1184–1191. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. Clusterprofiler: An R Package for Comparing Biological Themes among Gene Clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Nicodemus, K.K.; Liu, W.; Chase, G.A.; Tsai, Y.Y.; Fallin, M.D. Comparison of Type I Error for Multiple Test Corrections in Large Single-Nucleotide Polymorphism Studies Using Principal Components Versus Haplotype Blocking Algorithms. BMC Genet. 2005, 6 (Suppl. 1), S78. [Google Scholar] [CrossRef] [Green Version]
- Spina, E.; Cowin, P. Embryonic Mammary Gland Development. Semin. Cell Dev. Biol. 2021, 114, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Loisel, F.; Farmer, C.; van Hees, H.; Quesnel, H. Relative Prolactin-to-Progesterone Concentrations around Farrowing Influence Colostrum Yield in Primiparous Sows. Domest. Anim. Endocrinol. 2015, 53, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Wu, G. Regulatory Role for Amino Acids in Mammary Gland Growth and Milk Synthesis. Amino Acids 2009, 37, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Farmer, C. Review: Mammary Development in Lactating Sows: The Importance of Suckling. Animal 2019, 13, s20–s25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siouda, M.; Dujardin, A.D.; Barbollat-Boutrand, L.; Mendoza-Parra, M.A.; Gibert, B.; Ouzounova, M.; Bouaoud, J.; Tonon, L.; Robert, M.; Foy, J.P.; et al. Cdyl2 Epigenetically Regulates Mir124 to Control Nf-Kappab/Stat3-Dependent Breast Cancer Cell Plasticity. iScience 2020, 23, 101141. [Google Scholar] [CrossRef]
- Yang, W.; Feng, B.; Meng, Y.; Wang, J.; Geng, B.; Cui, Q.; Zhang, H.; Yang, Y.; Yang, J. Fam3c-Yy1 Axis Is Essential for Tgfbeta-Promoted Proliferation and Migration of Human Breast Cancer Mda-Mb-231 Cells Via the Activation of Hsf1. J. Cell. Mol. Med. 2019, 23, 3464–3475. [Google Scholar] [CrossRef]
- Pospiech, K.; Pluciennik, E.; Bednarek, A.K. Wwox Tumor Suppressor Gene in Breast Cancer, a Historical Perspective and Future Directions. Front. Oncol. 2018, 8, 345. [Google Scholar] [CrossRef]
- Song, X.; Zhang, C.; Liu, Z.; Liu, Q.; He, K.; Yu, Z. Characterization of Cerna Network to Reveal Potential Prognostic Biomarkers in Triple-Negative Breast Cancer. PeerJ 2019, 7, e7522. [Google Scholar] [CrossRef] [Green Version]
- Deming, S.L.; Lu, W.; Beeghly-Fadiel, A.; Zheng, Y.; Cai, Q.; Long, J.; Shu, X.O.; Gao, Y.T.; Zheng, W. Melatonin Pathway Genes and Breast Cancer Risk among Chinese Women. Breast Cancer Res. Treat. 2012, 132, 693–699. [Google Scholar] [CrossRef] [Green Version]
- Cenni, C.; Andres, S.; Hempel, M.; Strom, T.M.; Thomas, E.; Davies, A.; Timoney, N.; Frigiola, A.; Logan, M.; Holder-Espinasse, M. Tbx3 and Tbx5 Duplication: A Family with an Atypical Overlapping Holt-Oram/Ulnar-Mammary Syndrome Phenotype. Eur. J. Med. Genet. 2021, 64, 104213. [Google Scholar] [CrossRef]
- Slepicka, P.F.; Somasundara, A.V.H.; Santos, C.O.D. The Molecular Basis of Mammary Gland Development and Epithelial Differentiation. Semin. Cell Dev. Biol. 2021, 114, 93–112. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zhang, H.; Zhang, Z.; Gao, J.; Yang, J.; Wu, Z.; Fan, Y.; Xing, Y.; Li, L.; Xiao, S.; et al. Vrtn Is Required for the Development of Thoracic Vertebrae in Mammals. Int. J. Biol. Sci. 2018, 14, 667–681. [Google Scholar] [CrossRef] [PubMed]
- Gjorevski, N.; Nelson, C.M. Integrated Morphodynamic Signalling of the Mammary Gland. Nat. Rev. Mol. Cell Biol. 2011, 12, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.C.; Verheyen, E.M.; Zeng, Y.A. Mammary Development and Breast Cancer: A Wnt Perspective. Cancers 2016, 8, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, E.Y.; Hens, J.; Andl, T.; Kairo, A.; Yamaguchi, T.P.; Brisken, C.; Glick, A.; Wysolmerski, J.J.; Millar, S.E. Canonical Wnt Signaling Promotes Mammary Placode Development and Is Essential for Initiation of Mammary Gland Morphogenesis. Development 2004, 131, 4819–4829. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | N | Mean | SD | Min | Max | C.V. | SE | |
|---|---|---|---|---|---|---|---|---|
| LTN | 100 | 7.36 | 0.48 | 7.00 | 8.00 | 6.55 | 0.29 | 0.21 |
| RTN | 100 | 7.42 | 0.61 | 6.00 | 9.00 | 8.17 | 0.19 | 0.21 |
| TNUM | 100 | 14.78 | 0.97 | 14.00 | 17.00 | 6.56 | 0.36 | 0.23 |
| MAXAP | 100 | 7.50 | 0.59 | 7.00 | 9.00 | 7.93 | 0.38 | 0.22 |
| L-R | 100 | −0.06 | 0.51 | −2.00 | 2.00 | NA | 0.00 | 0.20 |
| ADIFF | 100 | 0.22 | 0.46 | 0.00 | 2.00 | NA | 0.24 | 0.20 |
| SNP | Trait | SSC | Position | Effect | p-Value | QTLs * | Annotation | Gene (Distance from the Gene in bp) |
|---|---|---|---|---|---|---|---|---|
| chr9:130956708 | LTN | 9 | 130956708 | 0.64 | 3.85 × 10−8 | intronic | PACC1(within), NENF(within) | |
| rs345573243 | RTN | 18 | 47399908 | 0.62 | 1.60 × 10−8 | 24,290, 7470 | intronic | OSBPL3(within) |
| rs703282466 | MAXAP | 3 | 106788730 | 0.45 | 2.29 × 10−8 | 5224, 8797, 8798, 4250, 4256 | intergenic | LTBP1(63796), ENSSSCG00000050704(9889) |
| chr15:137183045 | TNUM | 15 | 137183045 | −0.79 | 7.02 × 10−8 | 223,293 | intronic | MLPH(within) |
| rs345573243 | TNUM | 18 | 47399908 | 0.96 | 6.25 × 10−8 | 24,290, 7470 | intronic | OSBPL3(within) |
| rs1108940033 | ADIFF | 1 | 45796983 | 0.61 | 2.19 × 10−8 | intergenic | PHF3(500583),ENSSSCG00000049526(273685) | |
| rs321204530 | ADIFF | 1 | 177056841 | 0.59 | 1.44 × 10−8 | 5223, 5255, 822, 845, 1250 | intronic | MDGA2(within) |
| rs318957512 | ADIFF | 2 | 97849348 | 0.50 | 5.79 × 10−8 | intronic | ADGRV1(within) | |
| rs326371568 | ADIFF | 2 | 123586226 | 0.62 | 3.87 × 10−10 | 4255 | intergenic | FAM170A(119517),PRR16(613282) |
| rs342451777 | ADIFF | 2 | 127224122 | 0.57 | 8.31 × 10−9 | 4255 | intergenic | ENSSSCG00000042143(13463),ENSSSCG00000040936(55836) |
| rs325963999 | ADIFF | 3 | 19301837 | 0.66 | 9.15 × 10−9 | 5224, 7455, 7472 | intronic | KATNIP(within) |
| rs326276043 | ADIFF | 6 | 34051848 | 0.55 | 6.98 × 10−8 | 24,289 | intergenic | ENSSSCG00000050973(273191),CYLD(8093) |
| rs338649298 | ADIFF | 8 | 129552162 | 0.59 | 2.22 × 10−9 | intergenic | SNCA(163855),ENSSSCG00000043431(95940) | |
| rs321470648 | ADIFF | 9 | 10769337 | 0.45 | 5.38 × 10−8 | intergenic | ENSSSCG00000046278(36887),ENSSSCG00000045225(26362) | |
| rs1109963100 | ADIFF | 10 | 2911179 | 0.80 | 3.37 × 10−10 | intergenic | ENSSSCG00000042899(210778),BRINP3(52348) | |
| rs339887165 | ADIFF | 13 | 12135388 | 0.68 | 1.46 × 10−8 | 7479 | intergenic | ENSSSCG00000044771(16544),ENSSSCG00000051554(31818) |
| chr13:39266305 | ADIFF | 13 | 39266305 | 0.72 | 1.88 × 10−8 | 7479 | intronic | DNAH12(within) |
| rs701874475 | ADIFF | 13 | 134665423 | −0.51 | 2.19 × 10−8 | 7479 | intergenic | LMLN(8675),ENSSSCG00000050583(21280) |
| rs338558804 | ADIFF | 13 | 179683904 | −0.45 | 3.49 × 10−8 | intergenic | ENSSSCG00000038062(120975),NRIP1(140958) | |
| rs343864506 | ADIFF | 13 | 187708682 | −0.54 | 3.72 × 10−8 | intergenic | ENSSSCG00000047308(444803),ENSSSCG00000050420(500018) | |
| rs334271954 | ADIFF | 14 | 62959726 | 0.52 | 5.70 × 10−8 | intergenic | FAM13C(33877),SLC16A9(66937) | |
| rs1109225784 | ADIFF | 15 | 12581324 | 0.64 | 1.96 × 10−9 | intergenic | U6(133156),U6(186459) | |
| rs326978910 | ADIFF | 15 | 84015934 | 0.38 | 3.38 × 10−8 | 7468 | intronic | OSBPL6(within) |
| rs334746473 | ADIFF | 15 | 97200323 | 0.75 | 4.23 × 10−10 | 7468 | intergenic | ENSSSCG00000046205(186079),U2(688281) |
| rs322863105 | ADIFF | 17 | 7610979 | −0.88 | 6.01 × 10−11 | intergenic | ENSSSCG00000045345(132678),ENSSSCG00000047202(438984) | |
| chr17:8221026 | ADIFF | 17 | 8221026 | 0.79 | 1.47 × 10−9 | intergenic | U6(19092),FAT1(227734) | |
| rs330045817 | ADIFF | 17 | 8536301 | −0.71 | 3.76 × 10−10 | ncRNA_intronic | FAT1(within) | |
| rs324534432 | ADIFF | 17 | 13364668 | 0.79 | 1.47 × 10−9 | intergenic | PSD3(100469),ENSSSCG00000046441(3798) | |
| Indel | ||||||||
| chr6:7472906 | LTN | 6 | 7472906 | −0.41 | 1.06 × 10−6 | 24,289 | intronic | CDYL2(within) |
| rs695882779 | LTN | 9 | 3690507 | 0.39 | 9.84 × 10−7 | ncRNA_intronic | ENSSSCG00000049604(within) | |
| chr9:119650540 | LTN | 9 | 119650540 | −0.30 | 3.62 × 10−7 | intergenic | ENSSSCG00000044083(337170),ENSSSCG00000050832(4698) | |
| rs792699200 | LTN | 9 | 130899869 | 0.51 | 4.41 × 10−7 | intronic | PACC1(within) | |
| chr12:44292044 | LTN | 12 | 44292044 | −0.37 | 9.55 × 10−7 | 5227, 5261, 6472, 6479, 595, 2929 | intronic | NOS2(within) |
| chr13:194617904 | LTN | 13 | 194617904 | −0.31 | 1.30 × 10−6 | intergenic | KRTAP11-1(111382),ENSSSCG00000047315(5813) | |
| rs709659410 | RTN | 5 | 17969771 | 0.42 | 5.28 × 10−7 | 2927 | intergenic | KRT73(2682),KRT2(14544) |
| chr6:9186279 | RTN | 6 | 9186279 | 0.89 | 2.28 × 10−7 | 24,289 | intronic | WWOX(within) |
| rs790747253 | RTN | 15 | 100810568 | −0.61 | 1.28 × 10−6 | 7468 | intronic | PGAP1(within) |
| chr18:48316684 | RTN | 18 | 48316684 | 0.41 | 7.00 × 10−8 | 24,290, 7470 | intronic | STK31(within) |
| chr3:98429885 | MAXAP | 3 | 98429885 | 0.50 | 2.93 × 10−7 | 5224, 8797, 8798 | intergenic | ENSSSCG00000045166(59604),ENSSSCG00000046007(449247) |
| rs793312568 | MAXAP | 3 | 106792080 | 0.43 | 8.28 × 10−8 | 5224, 8797, 8798, 4250, 4256 | intergenic | LTBP1(67153),ENSSSCG00000050704(6530) |
| chr6:9186279 | MAXAP | 6 | 9186279 | 0.82 | 1.59 × 10−6 | 24,289 | intronic | WWOX(within) |
| chr6:30642639 | MAXAP | 6 | 30642639 | 0.43 | 1.31 × 10−6 | 24,289 | intergenic | ENSSSCG00000047270(70049),ENSSSCG00000041426(189382) |
| chr9:131996847 | MAXAP | 9 | 131996847 | −0.49 | 1.46 × 10−6 | intronic | ENSSSCG00000040650(within) | |
| chr14:12750600 | MAXAP | 14 | 12750600 | 0.43 | 1.31 × 10−6 | intronic | HMBOX1(within) | |
| rs790747253 | MAXAP | 15 | 100810568 | −0.60 | 1.17 × 10−6 | 7468 | intronic | PGAP1(within) |
| rs711984029 | MAXAP | 17 | 13041965 | −0.47 | 1.68 × 10−6 | intronic | PSD3(within) | |
| rs793312568 | TNUM | 3 | 106792080 | 0.63 | 1.41 × 10−6 | 5224, 8797, 8798, 4250, 4256 | intergenic | LTBP1(67153),ENSSSCG00000050704(6530) |
| chr5:75592729 | TNUM | 5 | 75592729 | 0.98 | 8.38 × 10−7 | intronic | NELL2(within) | |
| rs792699200 | TNUM | 9 | 130899869 | 1.00 | 9.67 × 10−7 | intronic | PACC1(within) | |
| chr18:48316684 | TNUM | 18 | 48316684 | 0.62 | 4.01 × 10−7 | 24,290, 7470 | intronic | STK31(within) |
| chr1:44096236 | LR | 1 | 44096236 | 0.69 | 1.25 × 10−7 | intergenic | ENSSSCG00000042072(159101),ENSSSCG00000045405(51729) | |
| chr1:44973455 | ADIFF | 1 | 44973455 | 0.70 | 1.96 × 10−7 | intronic | ZUP1(within),RSPH4A(within) | |
| chr1:46840905 | ADIFF | 1 | 46840905 | 0.71 | 5.46 × 10−8 | intergenic | ENSSSCG00000050391(123491),U6(392319) | |
| chr1:65957275 | ADIFF | 1 | 65957275 | 0.43 | 1.14 × 10−7 | intergenic | FBXL4(21578),FAXC(274880) | |
| chr1:77875820 | ADIFF | 1 | 77875820 | 0.42 | 8.57 × 10−8 | intergenic | FYN(65243),U6(107381) | |
| chr1:118273285 | ADIFF | 1 | 118273285 | −0.64 | 1.96 × 10−9 | 5223, 6481, 5255 | intergenic | ENSSSCG00000049391(9019),ENSSSCG00000045826(5556) |
| rs1113667849 | ADIFF | 2 | 123584099 | 0.55 | 8.50 × 10−7 | 4255 | intergenic | FAM170A(117391),PRR16(615405) |
| chr3:54080763 | ADIFF | 3 | 54080763 | 0.44 | 1.70 × 10−6 | 5224, 7455, 7472, 6465 | intergenic | LONRF2(99093),REV1(54761) |
| chr3:122749868 | ADIFF | 3 | 122749868 | 0.76 | 1.91 × 10−7 | intergenic | LRATD1(17941),ENSSSCG00000045589(179131) | |
| chr6:9702570 | ADIFF | 6 | 9702570 | 0.61 | 3.15 × 10−7 | 24,289 | intronic | WWOX(within) |
| chr6:29725029 | ADIFF | 6 | 29725029 | −0.41 | 1.54 × 10−7 | 24,289 | intergenic | ENSSSCG00000034192(132519),CES5A(117783) |
| chr7:48226837 | ADIFF | 7 | 48226837 | 0.67 | 1.74 × 10−6 | 5257 | intronic | RASGRF1(within) |
| chr8:3878010 | ADIFF | 8 | 3878010 | −0.48 | 4.61 × 10−7 | 7477 | intronic | ENSSSCG00000027349(within) |
| chr8:131927486 | ADIFF | 8 | 131927486 | 0.68 | 1.46 × 10−8 | intergenic | AFF1(2666),ENSSSCG00000032190(41799) | |
| chr13:134712100 | ADIFF | 13 | 134712100 | −0.50 | 2.83 × 10−7 | 7479 | intergenic | ENSSSCG00000050583(20384),OSBPL11(6327) |
| chr13:188309252 | ADIFF | 13 | 188309252 | 0.35 | 4.51 × 10−7 | intergenic | ENSSSCG00000050420(95234),ENSSSCG00000043493(500979) | |
| chr13:191714830 | ADIFF | 13 | 191714830 | 0.76 | 1.91 × 10−7 | intergenic | ENSSSCG00000051384(248028),ENSSSCG00000048685(53357) | |
| chr15:12585471 | ADIFF | 15 | 12585471 | 0.67 | 1.74 × 10−6 | intergenic | U6(137304),U6(182308) | |
| chr15:84277014 | ADIFF | 15 | 84277014 | 0.53 | 6.57 × 10−9 | 7468 | intergenic | ENSSSCG00000036052(46519),ENSSSCG00000038561(122000) |
| rs792656057 | ADIFF | 16 | 69550278 | −0.60 | 8.69 × 10−7 | 5228 | intronic | GRIA1(within) |
| rs793561441 | ADIFF | 17 | 5622328 | −0.33 | 9.14 × 10−8 | intronic | PCM1(within) | |
| rs700363122 | ADIFF | 17 | 8173767 | 0.60 | 1.13 × 10−8 | intergenic | ENSSSCG00000047202(116838),U6(28063) | |
| rs789477433 | ADIFF | 18 | 25541841 | 0.43 | 8.35 × 10−7 | 24,290 | intergenic | ENSSSCG00000048651(270433),FAM3C(27695) |
| SNP | SSC | Position | Effect | p-Value | QTLs * | Annotation | Gene (Distance from the Gene in bp) |
|---|---|---|---|---|---|---|---|
| rs325963999 # | 3 | 19301837 | 0.27 | 7.82 × 10−15 | 5224, 7455, 7472 | intronic | KATNIP(within) |
| rs693622708 | 6 | 39540583 | 0.17 | 8.58 × 10−9 | 24,289 | intergenic | UQCRFS1(166710), ENSSSCG00000050718(41760) |
| rs326134805 | 8 | 88543901 | 0.13 | 3.81 × 10−15 | 7477, 1100 | intergenic | ENSSSCG00000044017(86061),SLC7A11(63983) |
| rs1109963100 # | 10 | 2911179 | 0.33 | 3.74 × 10−13 | intergenic | ENSSSCG00000042899(210778),BRINP3(52348) | |
| rs1113875395 | 12 | 11463144 | 0.16 | 7.51 × 10−9 | 5227, 1128 | intergenic | ABCA8(15848),ENSSSCG00000045738(20026) |
| rs343773900 | 13 | 110154062 | 0.24 | 5.42 × 10−15 | 7479 | intronic | PLD1(within) |
| rs333970515 | 13 | 132158230 | −0.18 | 3.99 × 10−13 | 7479 | ncRNA_exonic | ENSSSCG00000047632(within) |
| rs1109225784 # | 15 | 12581324 | 0.32 | 1.46 × 10−16 | intergenic | U6(133156),U6(186459) | |
| rs322863105 # | 17 | 7610979 | −0.34 | 3.34 × 10−13 | intergenic | ENSSSCG00000045345(132678),ENSSSCG00000047202(438984) | |
| Indel | |||||||
| chr1:77875820 # | 1 | 77875820 | 0.17 | 1.02 × 10−7 | intergenic | FYN(65243),U6(107381) | |
| chr1:118273285 # | 1 | 118273285 | −0.35 | 1.74 × 10−10 | 5223, 6481, 5255 | intergenic | ENSSSCG00000049391(9019),ENSSSCG00000045826(5556) |
| rs788352632 | 2 | 62598411 | −0.12 | 3.81 × 10−10 | 909 | ncRNA_intronic | ENSSSCG00000048292(within) |
| chr11:6896071 | 11 | 6896071 | −0.13 | 1.22 × 10−8 | 5260 | ncRNA_intronic | ENSSSCG00000036846(within) |
| rs787621311 | 13 | 110898097 | 0.23 | 2.27 × 10−7 | 7479 | intronic | FNDC3B(within) |
| rs701717756 | 14 | 38222283 | 0.26 | 9.42 × 10−10 | intergenic | RBM19(17536),ENSSSCG00000042669(3247) | |
| chr14:91416556 | 14 | 91416556 | −0.30 | 1.34 × 10−9 | intergenic | ENSSSCG00000047278(204580),CXCL12(99865) | |
| chr15:84277014 # | 15 | 84277014 | 0.22 | 2.92 × 10−9 | 7468 | intergenic | ENSSSCG00000036052(46519),ENSSSCG00000038561(122000) |
| rs700363122 # | 17 | 8173767 | 0.40 | 9.10 × 10−16 | intergenic | ENSSSCG00000047202(116838),U6(28063) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Li, H.; Zhong, Z.; Jiang, S. A Whole Genome Sequencing-Based Genome-Wide Association Study Reveals the Potential Associations of Teat Number in Qingping Pigs. Animals 2022, 12, 1057. https://doi.org/10.3390/ani12091057
Liu Z, Li H, Zhong Z, Jiang S. A Whole Genome Sequencing-Based Genome-Wide Association Study Reveals the Potential Associations of Teat Number in Qingping Pigs. Animals. 2022; 12(9):1057. https://doi.org/10.3390/ani12091057
Chicago/Turabian StyleLiu, Zezhang, Hong Li, Zhuxia Zhong, and Siwen Jiang. 2022. "A Whole Genome Sequencing-Based Genome-Wide Association Study Reveals the Potential Associations of Teat Number in Qingping Pigs" Animals 12, no. 9: 1057. https://doi.org/10.3390/ani12091057