
FGF21 Reduces Lipid Accumulation in Bovine Hepatocytes by Enhancing Lipid Oxidation and Reducing Lipogenesis via AMPK Signaling


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation, Culture of Hepatocytes, and FGF21 Treatment
2.2. Oil Red O Staining of Lipids in Hepatocytes
2.3. Determination of Extracellular VLDL and Intracellular TG Content
2.4. RNA Extraction and qRT-PCR Analysis
2.5. Immunofluorescence Staining
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
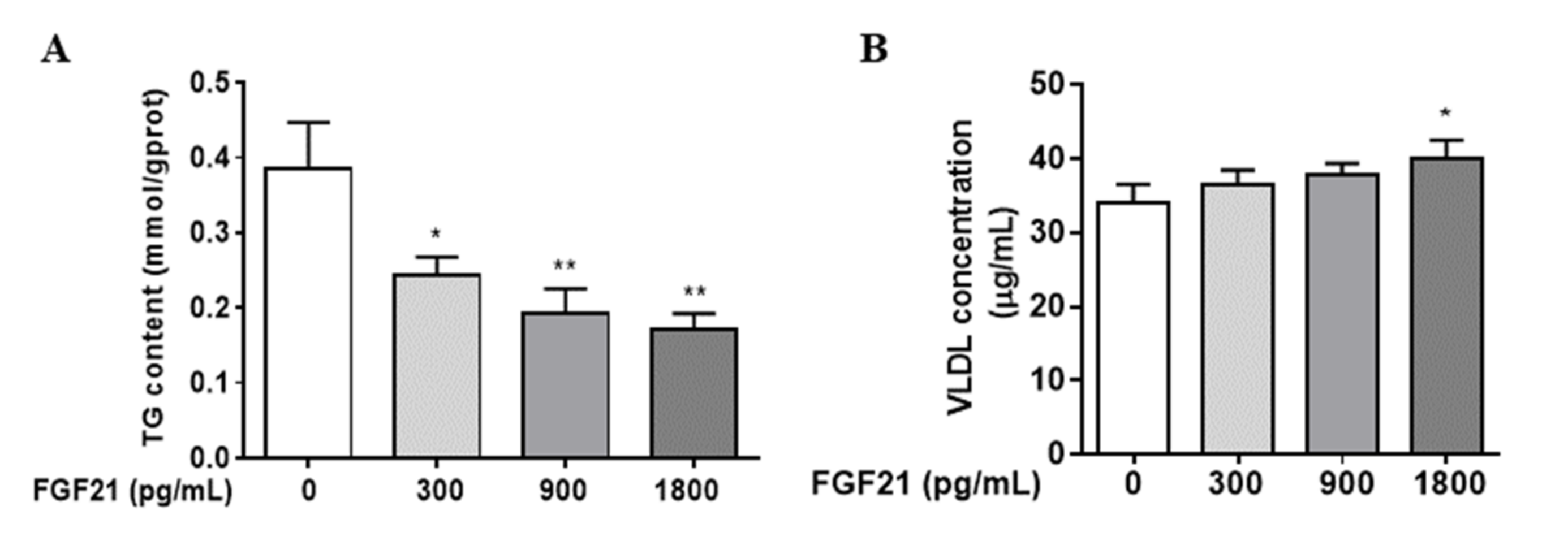
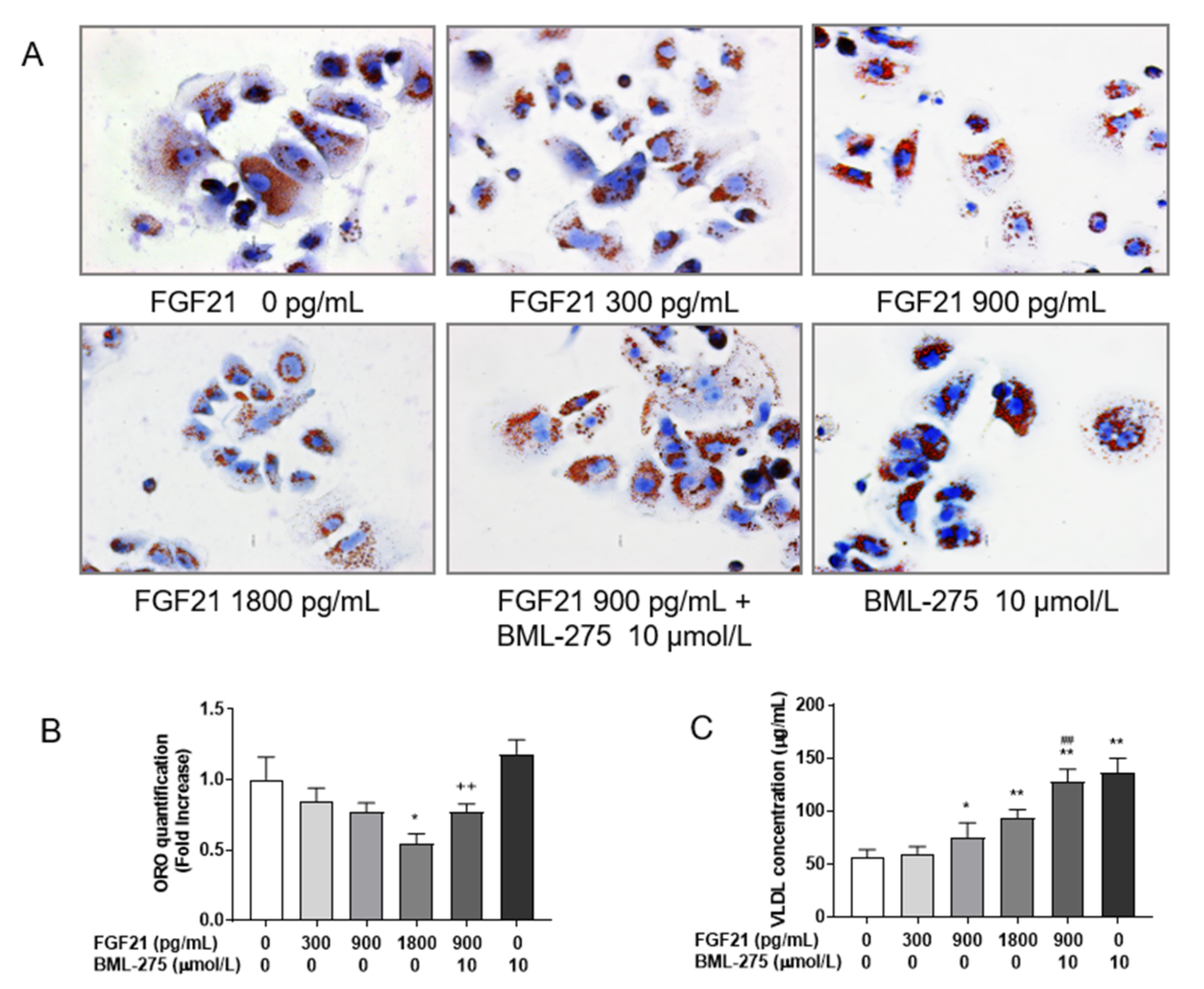
3.1. FGF21 Stimulates Hepatocellular VLDL Secretion, Resulting in a Reduction in Intracellular TG Content
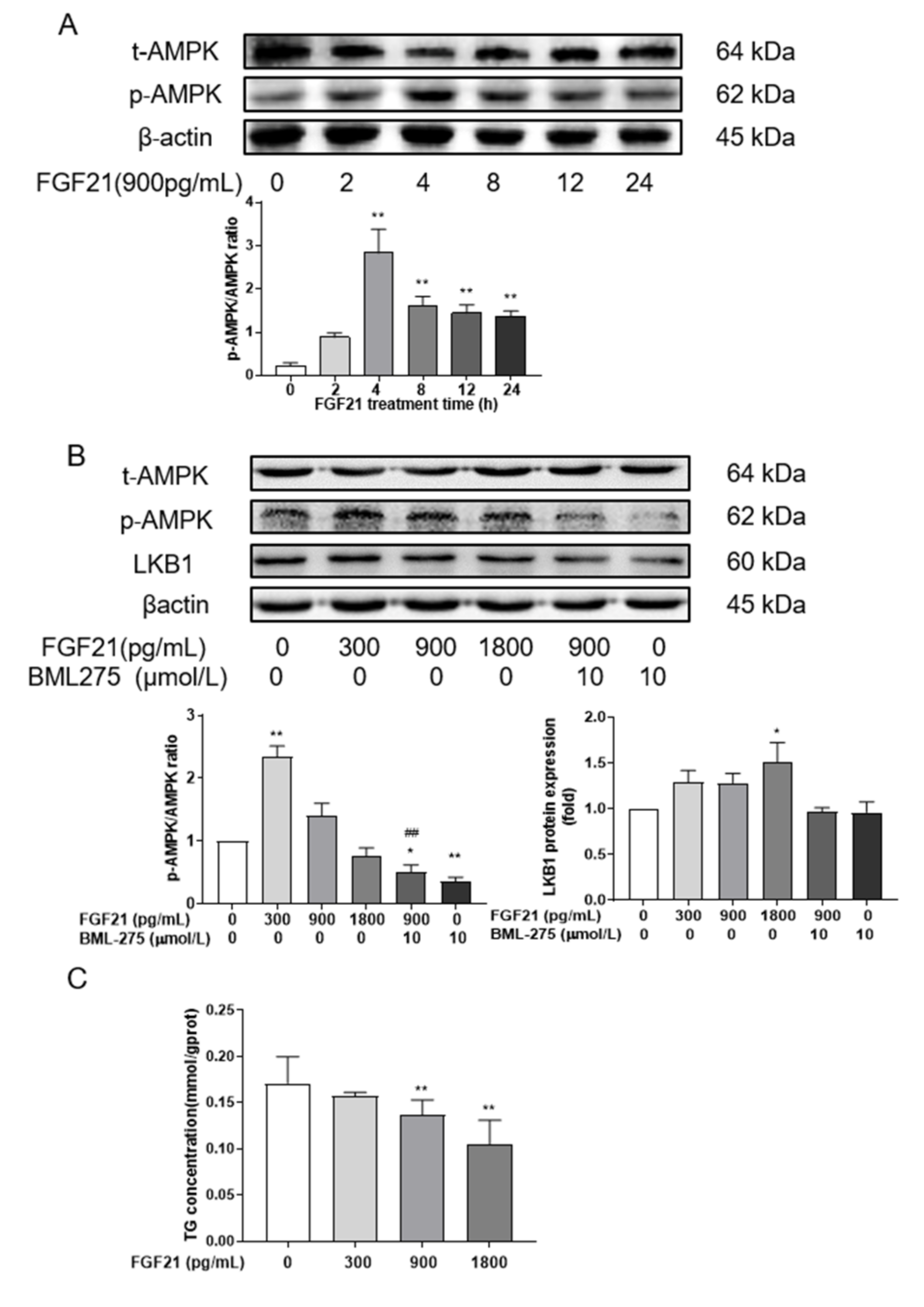
3.2. FGF21 Enhances AMPK Phosphorylation
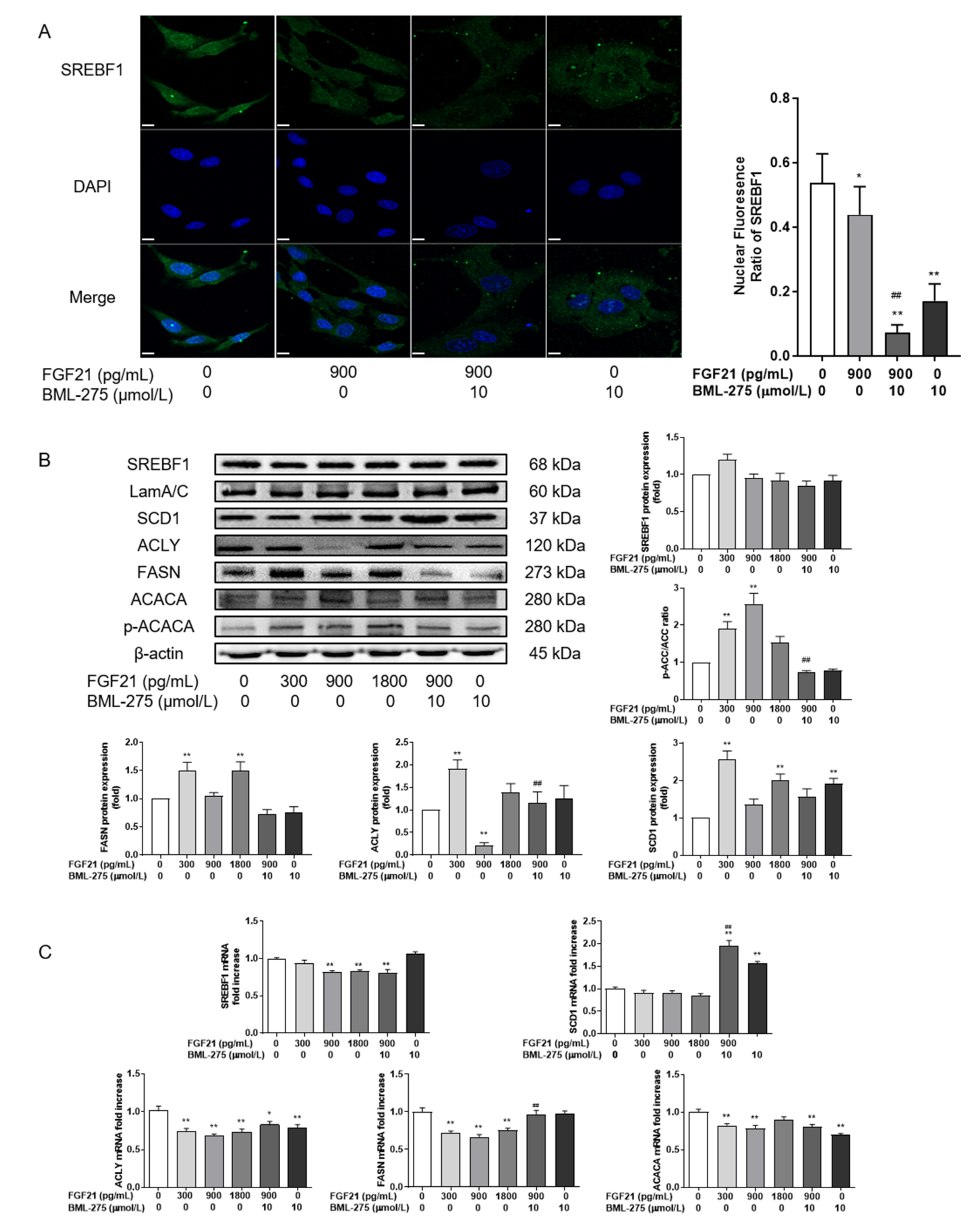
3.3. FGF21 Inhibits Lipid Synthesis by Reducing SREBF1 Nuclear Entry and Increasing ACACA Phosphorylation Ratio
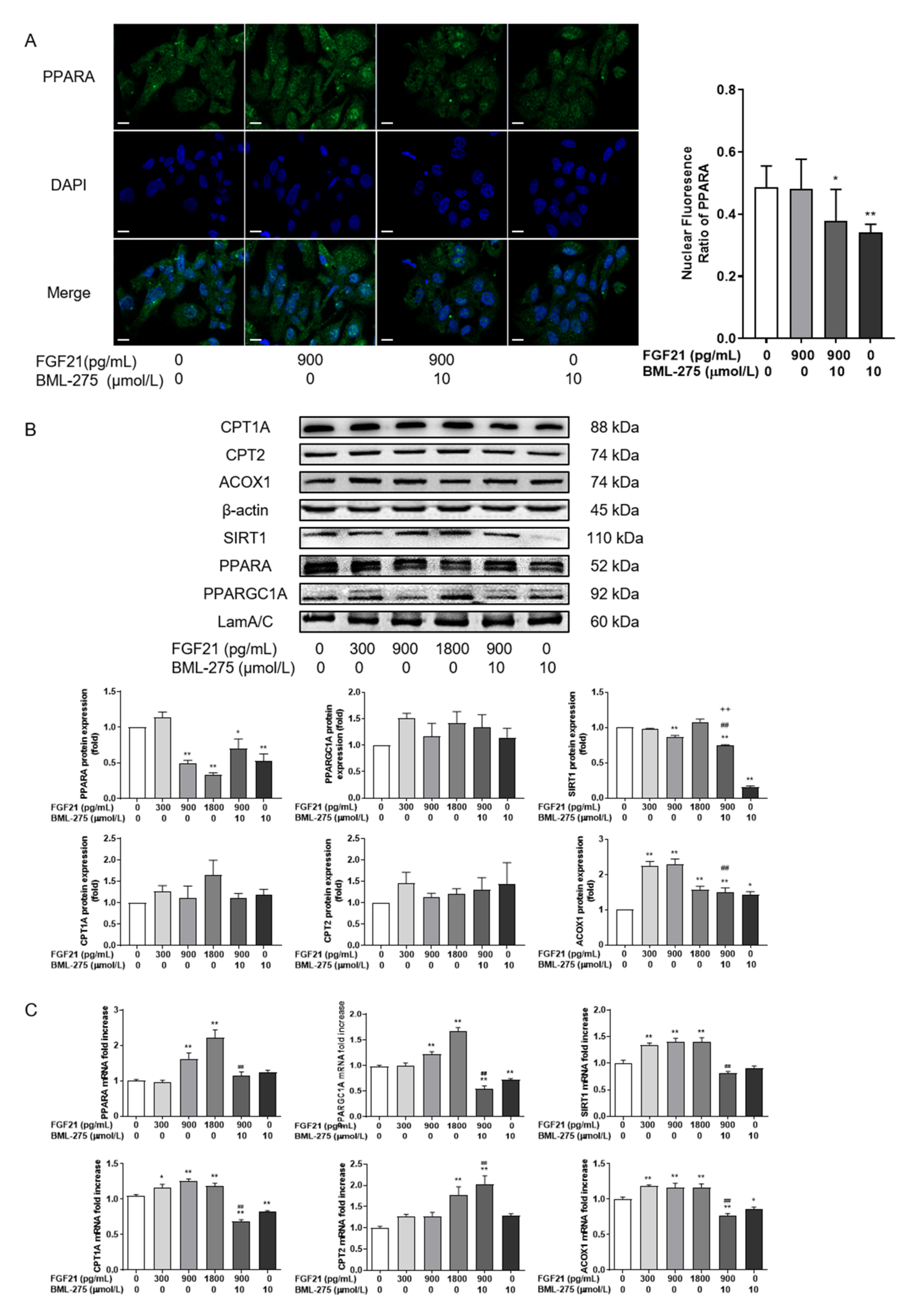
3.4. FGF21 Promotes the Expression of Key Proteins in Intracellular Fatty Acid Oxidation
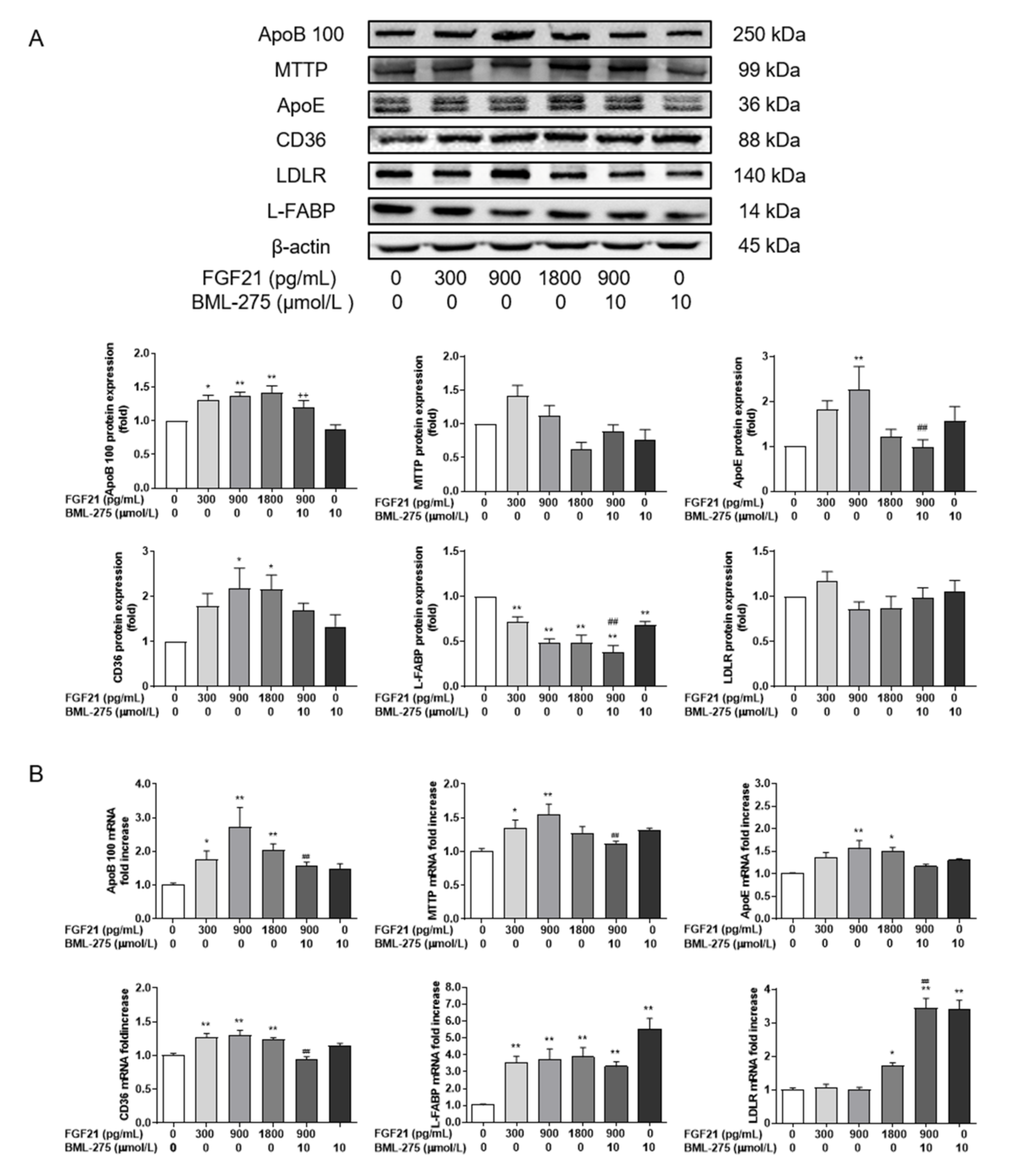
3.5. FGF21 Promotes Lipid Transport and Increases the Secretion of VLDL
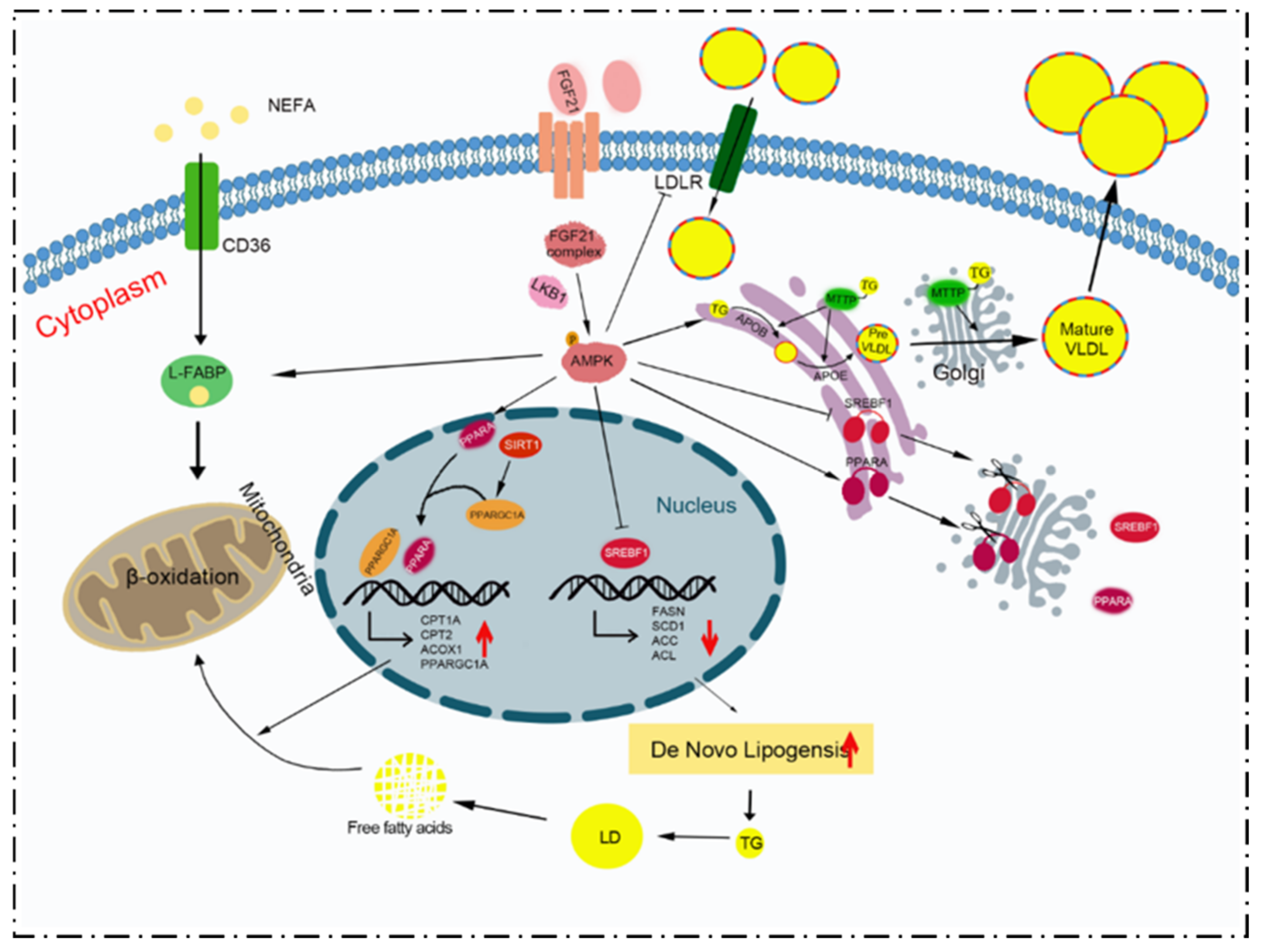
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kelton, D.F.; Lissemore, K.D.; Martin, R.E. Recommendations for Recording and Calculating the Incidence of Selected Clinical Diseases of Dairy Cattle. J. Dairy Sci. 1998, 81, 2502–2509. [Google Scholar] [CrossRef]
- Loor, J.J.; Everts, R.E.; Bionaz, M.; Dann, H.M.; Morin, D.E.; Oliveira, R.; Rodriguez-Zas, S.L.; Drackley, J.K.; Lewin, H.A. Nutrition-induced ketosis alters metabolic and signaling gene networks in liver of periparturient dairy cows. Physiol. Genom. 2007, 32, 105–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ringseis, R.; Gessner, D.K.; Eder, K. Molecular insights into the mechanisms of liver-associated diseases in early-lactating dairy cows: Hypothetical role of endoplasmic reticulum stress. J. Anim. Physiol. Anim. Nutr. 2015, 99, 626–645. [Google Scholar] [CrossRef] [PubMed]
- Woo, Y.C.; Xu, A.; Wang, Y.; Lam, K.S.L. Fibroblast Growth Factor 21 as an emerging metabolic regulator: Clinical perspectives. Clin. Endocrinol. 2013, 78, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Fisher, F.M.; Maratos-Flier, E. Understanding the Physiology of FGF21. Annu. Rev. Physiol. 2016, 78, 223–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Chen, L.; Li, F.; Yuan, Y.; Wang, Y.; Xia, W.; Zhang, Y.; Xu, Y.; Yang, Z.; Gao, B.; et al. HRD1-ERAD controls production of the hepatokine FGF21 through CREBH polyubiquitination. EMBO J. 2018, 37, e98942. [Google Scholar] [CrossRef] [PubMed]
- Camporez, J.P.; Asrih, M.; Zhang, D.; Kahn, M.; Samuel, V.T.; Jurczak, M.J.; Jornayvaz, F.R. Hepatic insulin resistance and increased hepatic glucose production in mice lacking Fgf21. J. Endocrinol. 2015, 226, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Dai, J.; Yang, M.; Deng, G.; Xu, S.; Jia, Y.; Boden, G.; Ma, Z.A.; Yang, G.; Li, L. Silencing of FGF-21 expression promotes hepatic gluconeogenesis and glycogenolysis by regulation of the STAT3-SOCS3 signal. FEBS J. 2014, 281, 2136–2147. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, T.; Dutchak, P.; Zhao, G.; Ding, X.; Gautron, L.; Parameswara, V.; Li, Y.; Goetz, R.; Mohammadi, M.; Esser, V.; et al. Endocrine regulation of the fasting response by PPARalpha-mediated induction of fibroblast growth factor 21. Cell Metab. 2007, 5, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Chau, M.D.; Gao, J.; Yang, Q.; Wu, Z.; Gromada, J. Fibroblast growth factor 21 regulates energy metabolism by activating the AMPK–SIRT1–PGC-1α pathway. Proc. Natl. Acad. Sci. USA 2010, 107, 12553–12558. [Google Scholar] [CrossRef] [Green Version]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Regulation of longevity by FGF21: Interaction between energy metabolism and stress responses. Ageing Res. Rev. 2017, 37, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kauppinen, A.; Kaarniranta, K. FGF21 activates AMPK signaling: Impact on metabolic regulation and the aging process. J. Mol. Med. 2017, 95, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lei, T.; Huang, J.; Wang, S.; Zhou, L.; Yang, Z.; Chen, X. The link between fibroblast growth factor 21 and sterol regulatory element binding protein 1c during lipogenesis in hepatocytes. Mol. Cell. Endocrinol. 2011, 342, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.M.; Hu, Z.M.; Cui, A.Y.; Liu, Z.S.; Ma, F.G.; Xue, Y.Q.; Liu, Y.X.; Zhang, F.F.; Zhao, Z.H.; Yu, Y.Y.; et al. Post-translational regulation of lipogenesis via AMPK-dependent phosphorylation of insulin-induced gene. Nat. Commun. 2019, 10, 623. [Google Scholar] [CrossRef] [Green Version]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell Biol. 2012, 13, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhu, X.; She, G.; Kong, Y.; Guo, Y.; Wang, Z.; Liu, G.; Zhao, B. Serum hepatokines in dairy cows: Periparturient variation and changes in energy-related metabolic disorders. BMC Vet. Res. 2018, 14, 236. [Google Scholar] [CrossRef]
- Wang, J.G.; Guo, Y.Z.; Kong, Y.Z.; Dai, S.; Zhao, B.Y. High non-esterified fatty acid concentrations promote expression and secretion of fibroblast growth factor 21 in calf hepatocytes cultured in vitro. J. Anim. Physiol. Anim. Nutr. 2017, 102, e476–e481. [Google Scholar] [CrossRef] [Green Version]
- Caixeta, L.S.; Giesy, S.L.; Krumm, C.S.; Perfield, J.W.N.; Butterfield, A.; Schoenberg, K.M.; Beitz, D.C.; Boisclair, Y.R. Effect of circulating glucagon and free fatty acids on hepatic FGF21 production in dairy cows. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 313, R526–R534. [Google Scholar] [CrossRef]
- Liu, L.; Li, X.W.; Li, Y.; Guan, Y.; Song, Y.X.; Yin, L.H.; Chen, H.; Lei, L.C.; Liu, J.X.; Li, X.B.; et al. Effects of nonesterified fatty acids on the synthesis and assembly of very low density lipoprotein in bovine hepatocytes in vitro. J. Dairy Sci. 2014, 97, 1328–1335. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Zhao, C.; Kong, Y.; Tan, P.; Liu, S.; Liu, Y.; Zeng, F.; Yuan, Y.; Zhao, B.; Wang, J. Elucidation of the mechanism of NEFA-induced PERK-eIF2α signaling pathway regulation of lipid metabolism in bovine hepatocytes. J. Steroid Biochem. Mol. Biol. 2021, 211, 105893. [Google Scholar] [CrossRef]
- Du, X.; Shen, T.; Wang, H.; Qin, X.; Xing, D.; Ye, Q.; Shi, Z.; Fang, Z.; Zhu, Y.; Yang, Y.; et al. Adaptations of hepatic lipid metabolism and mitochondria in dairy cows with mild fatty liver. J. Dairy Sci. 2018, 101, 9544–9558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, Y.; Zhao, C.; Huang, Y.; Liu, Y.; Liu, S.; Guo, Y.; Li, M.; Xu, T.; Zhao, B.; Wang, J. Angiopoietin-like protein 4 promotes very-low-density lipoprotein assembly and secretion in bovine hepatocytes in vitro. IUBMB Life 2020, 72, 2710–2721. [Google Scholar] [CrossRef] [PubMed]
- Bobe, G.; Young, J.; Beitz, D. Invited Review: Pathology, Etiology, Prevention, and Treatment of Fatty Liver in Dairy Cows. J. Dairy Sci. 2004, 87, 3105–3124. [Google Scholar] [CrossRef] [Green Version]
- Greenfield, R.B.; Cecava, M.J.; Johnson, T.R.; Donkin, S.S. Impact of dietary protein amount and rumen undegradability on intake, peripartum liver triglyceride, plasma metabolites, and milk production in transition dairy cattle. J. Dairy Sci. 2000, 83, 703–710. [Google Scholar] [CrossRef]
- Grummer, R.R. Nutritional and management strategies for the prevention of fatty liver in dairy cattle. Vet. J. 2008, 176, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Caixeta, L.S.; Giesy, S.L.; Krumm, C.S.; Perfield, J.W., 2nd; Butterfield, A.; Boisclair, Y.R. Fibroblast growth factor-21 (FGF21) administration to early-lactating dairy cows. II. Pharmacokinetics, whole-animal performance, and lipid metabolism. J. Dairy Sci. 2019, 102, 11597–11608. [Google Scholar] [CrossRef]
- Schoenberg, K.M.; Giesy, S.L.; Harvatine, K.J.; Waldron, M.R.; Cheng, C.; Kharitonenkov, A.; Boisclair, Y.R. Plasma FGF21 Is Elevated by the Intense Lipid Mobilization of Lactation. Endocrinology 2011, 152, 4652–4661. [Google Scholar] [CrossRef] [Green Version]
- Kurosu, H.; Choi, M.; Ogawa, Y.; Dickson, A.S.; Goetz, R.; Eliseenkova, A.V.; Mohammadi, M.; Rosenblatt, K.P.; Kliewer, S.A.; Kuro, O.M. Tissue-specific expression of betaKlotho and fibroblast growth factor (FGF) receptor isoforms determines metabolic activity of FGF19 and FGF21. J. Biol. Chem. 2007, 282, 26687–26695. [Google Scholar] [CrossRef] [Green Version]
- Eder, K.; Gessner, D.K.; Ringseis, R. Fibroblast growth factor 21 in dairy cows: Current knowledge and potential relevance. J. Anim. Sci. Biotechnol. 2021, 12, 97. [Google Scholar] [CrossRef]
- Potthoff, M.J.; Inagaki, T.; Satapati, S.; Ding, X.; He, T.; Goetz, R.; Mohammadi, M.; Finck, B.N.; Mangelsdorf, D.J.; Kliewer, S.A.; et al. FGF21 induces PGC-1alpha and regulates carbohydrate and fatty acid metabolism during the adaptive starvation response. Proc. Natl. Acad. Sci. USA 2009, 106, 10853–10858. [Google Scholar] [CrossRef] [Green Version]
- Schlegel, G.; Ringseis, R.; Keller, J.; Schwarz, F.J.; Windisch, W.; Eder, K. Expression of fibroblast growth factor 21 in the liver of dairy cows in the transition period and during lactation. J. Anim. Physiol. Anim. Nutr. 2013, 97, 820–829. [Google Scholar] [CrossRef] [PubMed]
- Keinicke, H.; Sun, G.; Mentzel, C.M.J.; Fredholm, M.; John, L.M.; Andersen, B.; Raun, K.; Kjaergaard, M. FGF21 regulates hepatic metabolic pathways to improve steatosis and inflammation. Endocr. Connect. 2020, 9, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.T.; Yudell, B.E.; Loor, J.J. Regulation of energy metabolism by long-chain fatty acids. Prog. Lipid Res. 2014, 53, 124–144. [Google Scholar] [CrossRef] [PubMed]
- Angeli, E.; Trionfini, V.; Gareis, N.C.; Matiller, V.; Huber, E.; Rey, F.; Salvetti, N.R.; Ortega, H.H.; Hein, G.J. Protein and gene expression of relevant enzymes and nuclear receptor of hepatic lipid metabolism in grazing dairy cattle during the transition period. Res. Vet Sci. 2019, 123, 223–231. [Google Scholar] [CrossRef]
- Dobrzyn, A.; Dobrzyn, P.; Miyazaki, M.; Sampath, H.; Chu, K.; Ntambi, J.M. Stearoyl-CoA desaturase 1 deficiency increases CTP:choline cytidylyltransferase translocation into the membrane and enhances phosphatidylcholine synthesis in liver. J. Biol. Chem. 2005, 280, 23356–23362. [Google Scholar] [CrossRef] [Green Version]
- Kimura, I.; Ichimura, A.; Ohue-Kitano, R.; Igarashi, M. Free Fatty Acid Receptors in Health and Disease. Physiol. Rev. 2020, 100, 171–210. [Google Scholar] [CrossRef]
- Dreyer, C.; Krey, G.; Keller, H.; Givel, F.; Helftenbein, G.; Wahli, W. Control of the peroxisomal beta-oxidation pathway by a novel family of nuclear hormone receptors. Cell 1992, 68, 879–887. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Yang, W.; Xiao, C.; Fu, S.; Deng, Q.; Ding, H.; Wang, Z.; Liu, G.; Li, X. SREBP-1c overexpression induces triglycerides accumulation through increasing lipid synthesis and decreasing lipid oxidation and VLDL assembly in bovine hepatocytes. J. Steroid. Biochem. Mol. Biol. 2014, 143, 174–182. [Google Scholar] [CrossRef]
- Kersten, S. Integrated physiology and systems biology of PPARα. Mol. Metab. 2014, 3, 354–371. [Google Scholar] [CrossRef]
- Hostetler, H.A.; McIntosh, A.L.; Atshaves, B.P.; Storey, S.M.; Payne, H.R.; Kier, A.B.; Schroeder, F. L-FABP directly interacts with PPARalpha in cultured primary hepatocytes. J. Lipid Res. 2009, 50, 1663–1675. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Zheng, S.; Attie, A.D.; Keller, M.P.; Bernlohr, D.A.; Blaner, W.S.; Newberry, E.P.; Davidson, N.O.; Chen, A. Perilipin 5 and liver fatty acid binding protein function to restore quiescence in mouse hepatic stellate cells. J. Lipid Res. 2018, 59, 416–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pepino, M.Y.; Kuda, O.; Samovski, D.; Abumrad, N.A. Structure-function of CD36 and importance of fatty acid signal transduction in fat metabolism. Annu. Rev. Nutr. 2014, 34, 281–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nassir, F.; Adewole, O.L.; Brunt, E.M.; Abumrad, N.A. CD36 deletion reduces VLDL secretion, modulates liver prostaglandins, and exacerbates hepatic steatosis in ob/ob mice. J. Lipid Res. 2013, 54, 2988–2997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, H.M.; Holloway, G.P.; Steinberg, G.R. AMPK regulation of fatty acid metabolism and mitochondrial biogenesis: Implications for obesity. Mol. Cell. Endocrinol. 2013, 366, 135–151. [Google Scholar] [CrossRef]
- Degrace, P.; Moindrot, B.; Mohamed, I.; Gresti, J.; Du, Z.Y.; Chardigny, J.M.; Sébédio, J.L.; Clouet, P. Upregulation of liver VLDL receptor and FAT/CD36 expression in LDLR-/- ApoB100/100 mice fed trans-10, cis-12 conjugated linoleic acid. J. Lipid Res. 2006, 47, 2647–2655. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.; Siddiqi, S.A. Intracellular trafficking and secretion of VLDL. Arter. Throm. Vas. 2012, 32, 1079–1086. [Google Scholar] [CrossRef] [Green Version]
- Mensenkamp, A.R.; Jong, M.C.; Van, G.H.; van Luyn, M.J.; Bloks, V.; Havinga, R.; Voshol, P.J.; Hofker, M.H.; van Dijk, K.W.; Havekes, L.M. Apolipoprotein E participates in the regulation of very low density lipoprotein-triglyceride secretion by the liver. J. Biol. Chem. 1999, 274, 35711–35718. [Google Scholar] [CrossRef] [Green Version]
- Kummrow, E.; Hussain, M.M.; Pan, M.; Marsh, J.B.; Fisher, E.A. Myristic acid increases dense lipoprotein secretion by inhibiting apoB degradation and triglyceride recruitment. J. Lipid Res. 2002, 43, 2155–2163. [Google Scholar] [CrossRef] [Green Version]
- Ota, T.; Gayet, C.; Ginsberg, H.N. Inhibition of apolipoprotein B100 secretion by lipid-induced hepatic endoplasmic reticulum stress in rodents. J. Clin. Investig. 2008, 118, 316–332. [Google Scholar] [CrossRef] [Green Version]
- Qiu, W.; Zhang, J.; Dekker, M.J.; Wang, H.; Huang, J.; Brumell, J.H.; Adeli, K. Hepatic autophagy mediates endoplasmic reticulum stress-induced degradation of misfolded apolipoprotein B. Hepatology 2011, 53, 1515–1525. [Google Scholar] [CrossRef]
- Su, Q.; Tsai, J.; Xu, E.; Qiu, W.; Bereczki, E.; Santha, M.; Adeli, K. Apolipoprotein B100 acts as a molecular link between lipid-induced endoplasmic reticulum stress and hepatic insulin resistance. Hepatology 2009, 50, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M. Regulation of lipid metabolism via a connection between the endoplasmic reticulum and lipid droplets. Anat. Sci. Int. 2017, 92, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Dettlaff-Pokora, A.; Sledzinski, T.; Swierczynski, J. Up-regulation MTTP and ApoB gene expression in rat liver is related to post-lipectomy hypertriglyceridemia. Cell. Physiol. Biochem. 2015, 36, 1767–1777. [Google Scholar] [CrossRef] [PubMed]
- Sirwi, A.; Hussain, M.M. Lipid transfer proteins in the assembly of ApoB-containing lipoproteins. J. Lipid Res. 2018, 59, 1094–1102. [Google Scholar] [CrossRef] [Green Version]
- Koerner, C.M.; Roberts, B.S.; Neher, S.B. Endoplasmic reticulum quality control in lipoprotein metabolism. Mol. Cell. Endocrinol. 2019, 498, 110547. [Google Scholar] [CrossRef]
- Blasiole, D.A.; Oler, A.T.; Attie, A.D. Regulation of ApoB secretion by the low density lipoprotein receptor requires exit from the endoplasmic reticulum and interaction with ApoE or ApoB. J. Biol. Chem. 2008, 283, 11374–11381. [Google Scholar] [CrossRef] [Green Version]
- Larsson, S.L.; Josefin, S.; Björkegren, J. The low density lipoprotein receptor prevents secretion of dense ApoB100-containing lipoproteins from the liver. J. Biol. Chem. 2004, 279, 831–836. [Google Scholar] [CrossRef] [Green Version]
- Ferre, P.; Foufelle, F. SREBP-1c transcription factor and lipid homeostasis: Clinical perspective. Horm. Res. Paediatr. 2007, 68, 72–82. [Google Scholar] [CrossRef]
- Do, H.T.; Tselykh, T.V.; Makela, J.; Ho, T.H.; Olkkonen, V.M.; Bornhauser, B.C.; Korhonen, L.; Zelcer, N.; Lindholm, D. Fibroblast Growth Factor-21 (FGF21) Regulates Low-density Lipoprotein Receptor (LDLR) Levels in Cells via the E3-ubiquitin Ligase Mylip/Idol and the Canopy2 (Cnpy2)/Mylip-interacting Saposin-like Protein (Msap). J. Biol. Chem. 2012, 287, 12602–12611. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′–3′) | GenBank Accession No. |
|---|---|---|
| 18S | For. ACCCATTCGAACGTCTGCCCTATT | NR_036642.1 |
| Rev. TCCTTGGATGTGGTAGCCGTTTCT | ||
| SREBF1 | For. CTGACAGCTCCATTGACAAGGC | NM_001113302.1 |
| Rev. GGCTTCATGTAGGAATACCCTC | ||
| PPARGC1A | For. TGCTGCTCTGGTTGGTGAA | NM_177945.3 |
| Rev. AGGCTCGTTGTTGTACTGATTAG | ||
| PPARA | For. ATCAGATGGCTCCGTTATTACAG | NM_001034036.1 |
| Rev. CAGTATTGGCACTTATTCCGATTC | ||
| SIRT1 | For. TGTGTCATAGGTTAGGTGGTGAA | XM_024986767.1 |
| Rev. CTGAAGAATCTGGTGGTGAAGTT | ||
| ACACA | For. ATGAAGGCTGTGGTGATGGA | NM_174224.2 |
| Rev. TGGTGGTCTTGCTGAGTTGA | ||
| FASN | For. CAGCGACGTCAGCACACTGGATG | NM_001012669.1 |
| Rev. GCATGGCATCTCTCAGGACCAC | ||
| SCD1 | For. CCTGGTGTCCTGTTGTTGTG | NM_173959.4 |
| Rev. GTGTGGTGGTAGTTGTGGAAG | ||
| ACLY | For. GTCAACCTCACTCTGGATGGA | NM_001037457.1 |
| Rev. TCGTGGTGGAACAGGACATAG | ||
| CPT1A | For. GGAATCTGTGAAGCCTCTTATGAA | NM_001304989.2 |
| Rev. GCCTGGATGTGAGTCGGTAT | ||
| CPT2 | For. CCTTCCTTCCTGTCTTGGTATG | NM_001045889.2 |
| Rev. TTCAGAGGCACTCACAATGTTC | ||
| ACOX1 | For. TAAGCCTTTGCCAGGTATT | NC_015500.1 |
| Rev. ATGGTCCCGTAGGTCAG | ||
| CD36 | For. CTCATTGCTGGTGCTGTCATT | NM_001278621.1 |
| Rev. CCTTGGCTAGATAACGAACTCTG | ||
| LDLR | For. AATGCGAGTGTGAAGAGG | XM_010806777.3 |
| Rev. GGTGTCGTAGGAGGAGAA | ||
| L-FABP | For. GGAGGAGTGTGAGATGGAGTT | NM_175817.3 |
| Rev. CCTTCGTCATGGTACTGGTAAC | ||
| ApoB 100 | For. GAACAGAATGAGCAAGTGAAGAAC | XM_024999521.1 |
| Rev. AGGTCAAGTGATGGCAGAGAA | ||
| ApoE | For. GCCGCTTCTGGGATTACCT | NM_173991.2 |
| Rev. GGTTCCGCAAGTCCTCCAT | ||
| MTTP | For. GAACAGGATATACCACCAGAATCG | NM_001101834.1 |
| Rev. CTTCAGAACTTGACGGACCATT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, Y.; Zhao, C.; Tan, P.; Liu, S.; Huang, Y.; Zeng, F.; Ma, P.; Guo, Y.; Zhao, B.; Wang, J. FGF21 Reduces Lipid Accumulation in Bovine Hepatocytes by Enhancing Lipid Oxidation and Reducing Lipogenesis via AMPK Signaling. Animals 2022, 12, 939. https://doi.org/10.3390/ani12070939
Kong Y, Zhao C, Tan P, Liu S, Huang Y, Zeng F, Ma P, Guo Y, Zhao B, Wang J. FGF21 Reduces Lipid Accumulation in Bovine Hepatocytes by Enhancing Lipid Oxidation and Reducing Lipogenesis via AMPK Signaling. Animals. 2022; 12(7):939. https://doi.org/10.3390/ani12070939
Chicago/Turabian StyleKong, Yezi, Chenxu Zhao, Panpan Tan, Siqi Liu, Yan Huang, Fangyuan Zeng, Pingjun Ma, Yazhou Guo, Baoyu Zhao, and Jianguo Wang. 2022. "FGF21 Reduces Lipid Accumulation in Bovine Hepatocytes by Enhancing Lipid Oxidation and Reducing Lipogenesis via AMPK Signaling" Animals 12, no. 7: 939. https://doi.org/10.3390/ani12070939