
Extensive Sheep and Goat Production: The Role of Novel Technologies towards Sustainability and Animal Welfare

 ,
,  , , , , , , , , , ,
, , , , , , , , , ,  ,
,  , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Sheep and Goat Extensive Production Systems: The Importance of Assessing Welfare
3. Animal Welfare in Small Ruminant under Extensive Production Systems: The Role of Novel Technologies
3.1. The Use of Precision Livestock Farming Applied to Sheep and Goat Extensive Production
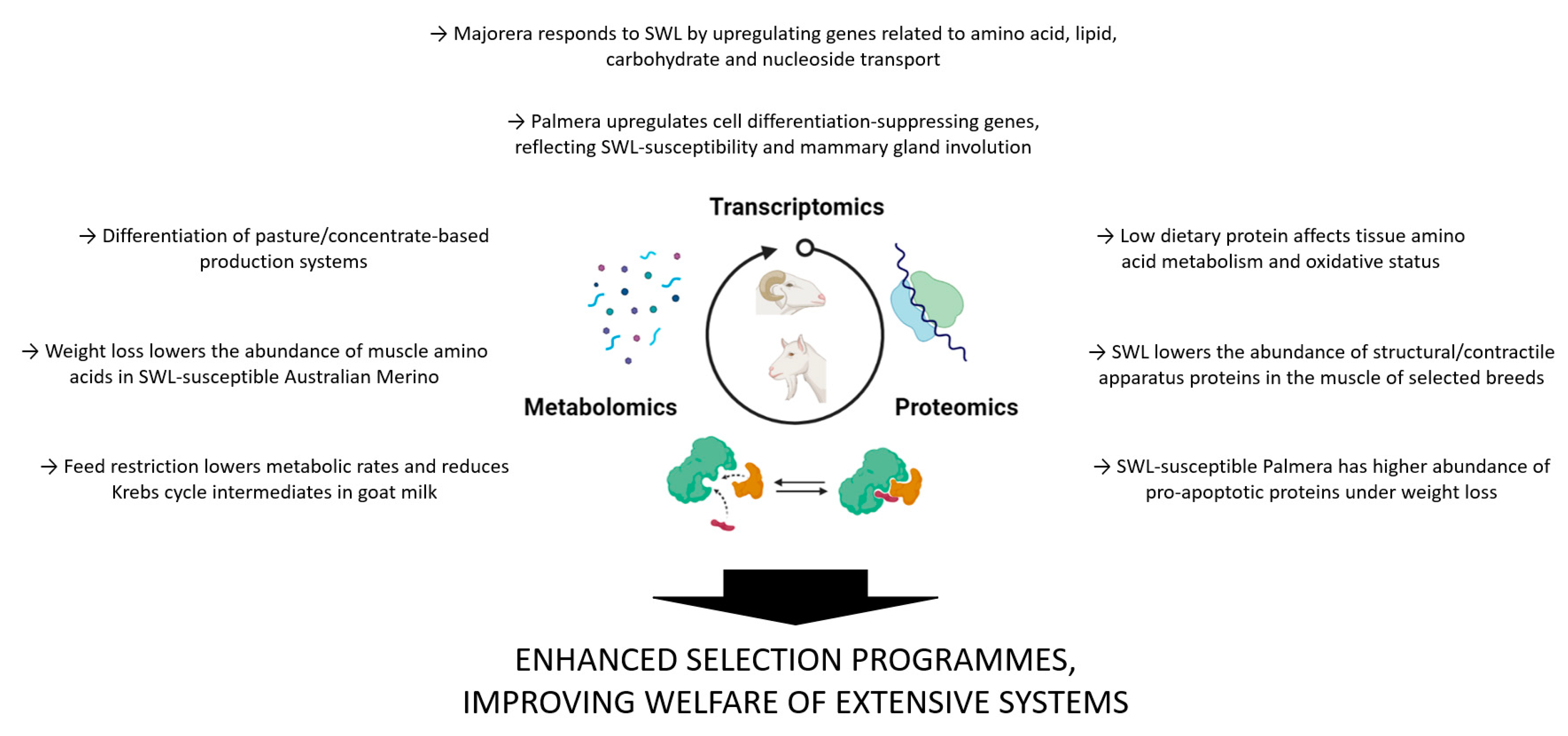
3.2. The Use of Omics Applied to Sheep and Goat Extensive Production
3.3. Novel Technologies and Thermal Stress in Sheep and Goat Extensive Production
3.4. Novel Technologies and Colostrum Intake on Lamb and Kid Survival
3.5. Novel Technologies and the Diagnostics of Metabolic Diseases in Sheep and Goat Extensive Production: Pregnancy Toxemia
3.6. Novel Technologies towards Parasite Resistance in Small Ruminant Extensive Production
4. General Conclusions and Future Prospects
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Rancourt, M.; Fois, N.; Lavín, M.P.; Tchakérian, E.; Vallerand, F. Mediterranean sheep and goats production: An uncertain future. Small Rumin. Res. 2006, 62, 167–179. [Google Scholar] [CrossRef]
- Bertolozzi-Caredio, D.; Garrido, A.; Soriano, B.; Bardaji, I. Implications of alternative farm management patterns to promote resilience in extensive sheep farming. A Spanish case study. J. Rural Stud. 2021, 86, 633–644. [Google Scholar] [CrossRef]
- Teixeira, A.; Silva, S.; Guedes, C.; Rodrigues, S. Sheep and goat meat processed products quality: A Review. Foods 2020, 9, 960. [Google Scholar] [CrossRef] [PubMed]
- Wishart, H.M. Precision Livestock Farming: Potential Application for Sheep Systems in Harsh Environments. Ph.D. Thesis, The University of Edinburgh, Edinburgh, UK, 2019. [Google Scholar]
- Goddard, P.; Waterhouse, T.; Dwyer, C.; Stott, A. The perception of the welfare of sheep in extensive systems. Small Rumin. Res. 2006, 62, 215–225. [Google Scholar] [CrossRef]
- Temple, D.; Manteca, X. Animal welfare in extensive production systems is still an area of concern. Front. Sustain. Food Syst. 2020, 4, 545902. [Google Scholar] [CrossRef]
- Dwyer, C. Welfare of sheep: Providing for welfare in an extensive environment. Small Rumin. Res. 2009, 86, 14–21. [Google Scholar] [CrossRef]
- Villalba, J.J.; Manteca, X.; Vercoe, P.E.; Maloney, S.K.; Blache, D. Integrating nutrition and animal welfare in extensive systems. In Nutrition and the Welfare of Farm Animals; Philips, C.J.C., Ed.; Springer : Cham, Switzerland, 2016; Volume 16, pp. 135–163. [Google Scholar]
- Dwyer, C.M.; Conington, J.; Corbiere, F.; Holmøy, I.H.; Muri, K.; Nowak, R.; Rooke, J.; Vipond, J.; Gautier, J.-M. Invited review: Improving neonatal survival in small ruminants: Science into practice. Animal 2016, 10, 449–459. [Google Scholar] [CrossRef] [Green Version]
- Freitas-De-Melo, A.; Terrazas, A.; Ungerfeld, R.; Hötzel, M.J.; Orihuela, A.; Pérez-Clariget, R. Influence of low pasture allowance during pregnancy on the attachment between ewes and their lambs at birth and during lactation. Appl. Anim. Behav. Sci. 2018, 199, 9–16. [Google Scholar] [CrossRef]
- Karthik, D.; Suresh, J.; Reddy, Y.R.; Sharma, G.R.K.; Ramana, J.V.; Gangaraju, G.; Yasaswini, D.; Adegbeye, M.J.; Reddy, P.R.K. Farming systems in sheep rearing: Impact on growth and reproductive performance, nutrient digestibility, disease incidence and heat stress indices. PLoS ONE 2021, 16, e0244922. [Google Scholar] [CrossRef]
- Waterhouse, A.; Holland, J.P.; McLaren, A.; Arthur, R.; Duthie, C.A.; Kodam, S.; Wishart, H.M. Opportunities and challenges for real-time management (RTM) in extensive livestock systems. In Precision Livestock Farming ’19, Proceedings of the European Conference in Precision Livestock Farming, Cork, Ireland, 26–29 August 2019; O’Brien, B., Hennessy, D., Shalloo, L., Eds.; Scotland’s Rural College: Edinburgh, Scotland, 2019; pp. 20–26. [Google Scholar]
- Munoz, C.A.; Campbell, A.; Hemsworth, P.H.; Doyle, R.E. Evaluating the welfare of extensively managed sheep. PLoS ONE 2019, 14, e0218603. [Google Scholar] [CrossRef]
- Dwyer, C. The behaviour and goats of sheep. In The Ethology of Domestic Animals: An Introductory Text; Jensen, P., Ed.; CABI: Wallingford, UK, 2017; pp. 199–213. [Google Scholar] [CrossRef]
- Sejian, V.; Silpa, M.; Nair, M.R.; Devaraj, C.; Krishnan, G.; Bagath, M.; Chauhan, S.; Suganthi, R.; Fonseca, V.; König, S.; et al. Heat stress and goat welfare: Adaptation and production considerations. Animals 2021, 11, 1021. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Mondal, T. Heat stress and thermoregulatory responses of goats: A review. Biol. Rhythm Res. 2019, 52, 407–433. [Google Scholar] [CrossRef]
- Caja, G.; Castro-Costa, A.; Salama, A.A.; Oliver, J.; Baratta, M.; Ferrer, C.; Knight, C.H. Sensing solutions for improving the performance, health and wellbeing of small ruminants. J. Dairy Res. 2020, 87, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Vaintrub, M.O.; Levit, H.; Chincarini, M.; Fusaro, I.; Giammarco, M.; Vignola, G. Review: Precision livestock farming, automats and new technologies: Possible applications in extensive dairy sheep farming. Animal 2020, 15, 100143. [Google Scholar] [CrossRef]
- Waterhouse, A. Precision livestock farming (PLF) technology and real-time monitoring should improve welfare in extensive systems, but does it change the duty of care and require modification of welfare guidelines for livestock keepers? In Precision Livestock Farming ’19, Proceedings of the European Conference in Precision Livestock Farming, Cork, Ireland, 26–29 August 2019; O’Brien, B., Hennessy, D., Shalloo, L., Eds.; Teagasc, Animal & Grassland Research and Innovation Centre, Moorepark: Fermoy, Ireland, 2019; pp. 256–261. [Google Scholar]
- Herlin, A.; Brunberg, E.; Hultgren, J.; Högberg, N.; Rydberg, A.; Skarin, A. Animal Welfare Implications of Digital Tools for Monitoring and Management of Cattle and Sheep on Pasture. Animals 2021, 11, 829. [Google Scholar] [CrossRef]
- Morgan-Davies, C.; Lambe, N.; Wishart, H.; Waterhouse, A.; Kenyon, F.; McBean, D.; McCracken, D. Impacts of using a precision livestock system targeted approach in mountain sheep flocks. Livest. Sci. 2018, 208, 67–76. [Google Scholar] [CrossRef]
- Umstätter, C.; Waterhouse, A.; Holland, J. An automated sensor-based method of simple behavioural classification of sheep in extensive systems. Comput. Electron. Agric. 2008, 64, 19–26. [Google Scholar] [CrossRef]
- Spigarelli, C.; Zuliani, A.; Battini, M.; Mattiello, S.; Bovolenta, S. Welfare Assessment on Pasture: A Review on Animal-Based Measures for Ruminants. Animals 2020, 10, 609. [Google Scholar] [CrossRef] [Green Version]
- Bodas, R.; García-García, J.J.; Montañés, M.; Benito, A.; Peric, T.; Baratta, M.; Viola, I.; Geß, A.; Ko, N.; Cadavez, V.; et al. On farm welfare assessment of European fattening lambs. Small Rumin. Res. 2021, 204, 106533. [Google Scholar] [CrossRef]
- AWINa. AWIN Welfare Assessment Protocol for Goats. 2015. Available online: https://www.researchgate.net/publication/275341689_AWIN_welfare_assessment_protocol_for_goats (accessed on 20 January 2022). [CrossRef]
- AWINb. AWIN Welfare Assessment Protocol for Sheep. 2015. Available online: https://www.researchgate.net/publication/275887069_AWIN_Welfare_Assessment_Protocol_for_Sheep (accessed on 20 January 2022). [CrossRef]
- Battini, M.; Renna, M.; Giammarino, M.; Battaglini, L.; Mattiello, S. Feasibility and Reliability of the AWIN Welfare Assessment Protocol for Dairy Goats in Semi-extensive Farming Conditions. Front. Veter-Sci. 2021, 8, 731927. [Google Scholar] [CrossRef]
- Kosgey, I.; Rowlands, G.; van Arendonk, J.; Baker, R. Small ruminant production in smallholder and pastoral/extensive farming systems in Kenya. Small Rumin. Res. 2008, 77, 11–24. [Google Scholar] [CrossRef]
- Gizaw, S.; Getachew, T.; Edea, Z.; Mirkena, T.; Duguma, G.; Tibbo, M.; Rischkowsky, B.; Mwai, O.; Dessie, T.; Wurzinger, M.; et al. Characterization of Indigenous Breeding Strategies of the Sheep Farming Communities of Ethiopia: A Basis for Designing Community-Based Breeding Programs; International Center for Agricultural Research in the Dry Areas (ICARDA): Aleppo, Syria, 2013; p. 47. [Google Scholar]
- Gelasakis, A.; Rose, G.; Giannakou, R.; Valergakis, G.; Theodoridis, A.; Fortomaris, P.; Arsenos, G. Typology and characteristics of dairy goat production systems in Greece. Livest. Sci. 2017, 197, 22–29. [Google Scholar] [CrossRef]
- Napolitano, F.; Arney, D.; Mota-Rojas, D.; De Rosa, G. Reproductive technologies and animal welfare. In Reproductive Technologies in Animals; Presicce, G.A., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 275–286. [Google Scholar] [CrossRef]
- Needham, T.; Lambrechts, H.; Hoffman, L. Castration of male livestock and the potential of immunocastration to improve animal welfare and production traits. S. Afr. J. Anim. Sci. 2017, 47, 731–742. [Google Scholar] [CrossRef] [Green Version]
- Cloete, S.W.; Burger, M.; Scholtz, A.J.; Cloete, J.J.; Kruger, A.C.; Dzama, K. Arena behaviour of Merino weaners is heritable and affected by divergent selection for number of lambs weaned per ewe mated. Appl. Anim. Behav. Sci. 2020, 233, 105152. [Google Scholar] [CrossRef]
- Bonato, M.; Cloete, J.; Kruger, A.; Cloete, S.W. Behavioural reactivity of two lines of South African Merino sheep divergently selected for reproductive potential. Appl. Anim. Behav. Sci. 2020, 234, 105209. [Google Scholar] [CrossRef]
- Napolitano, F.; De Rosa, G.; Ferrante, V.; Grasso, F.; Braghieri, A. Monitoring the welfare of sheep in organic and conventional farms using an ANI 35 L derived method. Small Rumin. Res. 2009, 83, 49–57. [Google Scholar] [CrossRef]
- Richmond, S.E.; Wemelsfelder, F.; De Heredia, I.B.; Ruiz, R.; Canali, E.; Dwyer, C.M. Evaluation of Animal-Based Indicators to Be Used in a Welfare Assessment Protocol for Sheep. Front. Vet.-Sci. 2017, 4, 210. [Google Scholar] [CrossRef]
- Salas, M.Á.S.; Mondragón-Ancelmo, J.; Badillo, M.D.R.J.; Licea, G.R.; Napolitano, F. Assessing dairy goat welfare in intensive or semi-intensive farming conditions in Mexico. J. Dairy Sci. 2021, 104, 19557. [Google Scholar] [CrossRef]
- Berihulay, H.; Abied, A.; He, X.; Jiang, L.; Ma, Y. Adaptation mechanisms of small ruminants to environmental heat stress. Animals 2019, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Leite, L.; Stamm, F.; Souza, R.; Filho, J.C.; Garcia, R. On-farm welfare assessment in dairy goats in the Brazilian Northeast. Arq. Bras. Med. Vet. Zootec. 2020, 72, 2308–2320. [Google Scholar] [CrossRef]
- De, K.; Sharma, S.; Kumawat, P.K.; Kumar, D.; Mohapatra, A.; Sahoo, A. Tree shade improves the comfort of sheep during extreme summer. J. Veter-Behav. 2020, 40, 103–107. [Google Scholar] [CrossRef]
- Marcone, G.; Kaart, T.; Piirsalu, P.; Arney, D. Panting scores as a measure of heat stress evaluation in sheep with access and with no access to shade. Appl. Anim. Behav. Sci. 2021, 240, 105350. [Google Scholar] [CrossRef]
- EFSA Panel on Animal Health and Welfare (AHAW) Scientific Opinion on the welfare risks related to the farming of sheep for wool, meat and milk production. EFSA J. 2014, 12, 3933. [CrossRef]
- Kenyon, P.R.; Maloney, S.K.; Blache, D. Review of sheep body condition score in relation to production characteristics. N. Z. J. Agric. Res. 2014, 57, 38–64. [Google Scholar] [CrossRef]
- Munoz, C.; Campbell, A.; Hemsworth, P.; Doyle, R. Animal-based measures to assess the welfare of extensively managed ewes. Animals 2017, 8, 2. [Google Scholar] [CrossRef] [Green Version]
- Mattiello, S.; Battini, M.; De Rosa, G.; Napolitano, F.; Dwyer, C. How can we assess positive welfare in ruminants? Animals 2019, 9, 758. [Google Scholar] [CrossRef] [Green Version]
- Mellor, D.J. Updating animal welfare thinking: Moving beyond the “Five Freedoms” towards “A Life Worth Living”. Animals 2016, 6, 21. [Google Scholar] [CrossRef]
- Hernandez, R.O.; Sánchez, J.A.; Romero, M.H. Iceberg Indicators for animal welfare in rural sheep farms using the five domains model approach. Animals 2020, 10, 2273. [Google Scholar] [CrossRef]
- Mellor, D.J.; Beausoleil, N.J. Extending the ‘Five Domains’ model for animal welfare assessment to incorporate positive welfare states. Anim. Welf. 2015, 24, 241–253. [Google Scholar] [CrossRef]
- Mellor, D.J. Operational details of the five domains model and its key applications to the assessment and management of animal welfare. Animals 2017, 7, 60. [Google Scholar] [CrossRef] [Green Version]
- Mellor, D.J.; Beausoleil, N.J.; Littlewood, K.E.; McLean, A.N.; McGreevy, P.D.; Jones, B.; Wilkins, C. The 2020 Five domains model: Including human–animal interactions in assessments of animal welfare. Animals 2020, 10, 1870. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, E.; Swain, D.; Cronin, G.; Trotter, M. A systematic review of the potential uses of on-animal sensors to monitor the welfare of sheep evaluated using the Five Domains Model as a framework. Anim. Welf. 2019, 28, 407–420. [Google Scholar] [CrossRef]
- Vigors, B. Citizens’ and farmers’ framing of ’positive animal welfare’ and the implications for framing positive welfare in communication. Animals 2019, 9, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halachmi, I.; Guarino, M. Editorial: Precision livestock farming: A ‘per animal’ approach using advanced monitoring technologies. Animal 2016, 10, 1482–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, J.E.; Cronin, G.M.; Bush, R.D. Improving sheep production and welfare in extensive systems through precision sheep management. Anim. Prod. Sci. 2012, 52, 665–670. [Google Scholar] [CrossRef]
- Halachmi, I.; Guarino, M.; Bewley, J.; Pastell, M. Smart animal agriculture: Application of real-time sensors to improve animal well-being and production. Annu. Rev. Anim. Biosci. 2019, 7, 403–425. [Google Scholar] [CrossRef]
- Dawkins, M.S. Does smart farming improve or damage animal welfare? Technology and what animals want. Front. Anim. Sci. 2021, 2, 736536. [Google Scholar] [CrossRef]
- Rutter, M. Smart technologies for detecting animal welfare status and delivering health remedies for rangeland systems. Rev. Sci. Tech. Off. Int. Epizoot. 2014, 33, 181–187. [Google Scholar] [CrossRef]
- Morgan-Davies, C.; Kyle, J.; Boman, I.; Wishart, H.; McLaren, A.; Fair, S.; Creighton, P. A comparison of farm labour, profitability, and carbon footprint of different management strategies in Northern European grassland sheep systems. Agric. Syst. 2021, 191, 103155. [Google Scholar] [CrossRef]
- Kaler, J.; Ruston, A. Technology adoption on farms: Using normalisation process theory to understand sheep farmers’ attitudes and behaviours in relation to using precision technology in flock management. Prev. Vet.-Med. 2019, 170, 104715. [Google Scholar] [CrossRef]
- Sarout, B.N.M.; Waterhouse, A.; Duthie, C.-A.; Poli, C.H.E.C.; Haskell, M.J.; Berger, A.; Umstatter, C. Assessment of circadian rhythm of activity combined with random regression model as a novel approach to monitoring sheep in an extensive system. Appl. Anim. Behav. Sci. 2018, 207, 26–38. [Google Scholar] [CrossRef]
- Di Virgilio, A.; Morales, J.M.; Lambertucci, S.; Shepard, E.; Wilson, R.P. Multi-dimensional Precision Livestock Farming: A potential toolbox for sustainable rangeland management. PeerJ 2018, 30, e4867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansbridge, N.; Mitsch, J.; Bollard, N.; Ellis, K.; Miguel-Pacheco, G.G.; Dottorini, T.; Kaler, J. Feature selection and comparison of machine learning algorithms in classification of grazing and rumination behaviour in sheep. Sensors 2018, 18, 3532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maroto-Molina, F.; Navarro, J.; Príncipe-Aguirre, K.; Gómez-Maqueda, I.; Guerrero-Ginel, J.E.; Garrido-Varo, A.; Pérez-Marín, D.C. A Low-Cost IoT-Based system to monitor the location of a whole herd. Sensors 2019, 19, 2298. [Google Scholar] [CrossRef] [Green Version]
- Ren, K.; Karlsson, J.; Liuska, M.; Hartikainen, M.; Hansen, I.; Jørgensen, G.H.M. A sensor-fusion-system for tracking sheep location and behaviour. Int. J. Distrib. Sens. Netw. 2020, 16, 1776. [Google Scholar] [CrossRef]
- Gonçalves, P.; Nóbrega, L.; Monteiro, A.; Pedreiras, P.; Rodrigues, P.; Esteves, F. SheepIT, an E-Shepherd system for weed control in vineyards: Experimental results and lessons learned. Animals 2021, 11, 2625. [Google Scholar] [CrossRef]
- Fogarty, E.S.; Cronin, G.; Swain, D.; Moraes, L.; Trotter, M. Development of a predictive model to identify the day of lambing in extensive sheep systems using autonomous Global Navigation Satellite System (GNSS). In Proceedings of the European Conference in Precision Livestock Farming, Precision Livestock Farming ’19, Cork, Ireland, 26–29 August 2019; O’Brien, B., Hennessy, D., Shalloo, L., Eds.; Teagasc, Animal & Grassland Research and Innovation Centre, Moorepark: Fermoy, Ireland, 2019; pp. 84–87. [Google Scholar]
- Fogarty, E.; Swain, D.; Cronin, G.; Moraes, L.; Trotter, M. Can accelerometer ear tags identify behavioural changes in sheep associated with parturition? Anim. Reprod. Sci. 2020, 216, 106345. [Google Scholar] [CrossRef]
- Barwick, J.; Lamb, D.; Dobos, R.; Schneider, D.; Welch, M.; Trotter, M. Predicting lameness in sheep activity using tri-axial acceleration signals. Animals 2018, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- Byrne, D.; Esmonde, H.; Berry, D.; McGovern, F.; Creighton, P.; McHugh, N. Sheep lameness detection from individual hoof load. Comput. Electron. Agric. 2019, 158, 241–248. [Google Scholar] [CrossRef]
- Brown, D.J.; Savage, D.B.; Hinch, G.N. Repeatability and frequency of in-paddock sheep walk-over weights: Implications for individual animal management. Anim. Prod. Sci. 2014, 54, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Wishart, H.; Morgan-Davies, C.; Waterhouse, A. A PLF approach for allocating supplementary feed to pregnant ewes in an extensive hill sheep system. In Proceedings of the 7th European Conference on Precision Livestock Farming, Milan, Italy, 15–18 September 2015; Guarino, M., Berckmans, D., Eds.; ECPLF: Milan, Italy, 2015; pp. 256–265. [Google Scholar]
- González-García, E.; Alhamada, M.; Pradel, J.; Douls, S.; Parisot, S.; Bocquier, F.; Menassol, J.; Llach, I.; González, L. A mobile and automated walk-over-weighing system for a close and remote monitoring of liveweight in sheep. Comput. Electron. Agric. 2018, 153, 226–238. [Google Scholar] [CrossRef]
- Kasper, C.; Ribeiro, D.; De Almeida, A.M.; Larzul, C.; Liaubet, L.; Murani, E. Omics application in animal science—a special emphasis on stress response and damaging behaviour in pigs. Genes 2020, 11, 920. [Google Scholar] [CrossRef]
- Yu, K.; Matzapetakis, M.; Horvatić, A.; Terré, M.; Bach, A.; Kuleš, J.; Yeste, N.; Gómez, N.; Arroyo, L.; Rodríguez-Tomàs, E.; et al. Metabolome and proteome changes in skeletal muscle and blood of pre-weaning calves fed leucine and threonine supplemented diets. J. Proteom. 2020, 216, 103677. [Google Scholar] [CrossRef] [PubMed]
- Marco-Ramell, A.; de Almeida, A.M.; Cristobal, S.; Rodrigues, P.; Roncada, P.; Bassols, A. Proteomics and the search for welfare and stress biomarkers in animal production in the one-health context. Mol. BioSyst. 2016, 12, 2024–2035. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.M.; Zachut, M.; Hernández-Castellano, L.E.; Šperanda, M.; Gabai, G.; Mobasheri, A. Biomarkers of fitness and welfare in dairy animals: Healthy living. J. Dairy Res. 2019, 86, 379–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, D.M.; Salama, A.A.; Vitor, A.C.; Argüello, A.; Moncau, C.T.; Santos, E.M.; Caja, G.; de Oliveira, J.S.; Balieiro, J.C.; Hernández-Castellano, L.E.; et al. The application of omics in ruminant production: A review in the tropical and sub-tropical animal production context. J. Proteom. 2020, 227, 103905. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.M.; Ali, S.A.; Ceciliani, F.; Eckersall, P.D.; Hernández-Castellano, L.E.; Han, R.; Hodnik, J.J.; Jaswal, S.; Lippolis, J.D.; McLaughlin, M.; et al. Domestic animal proteomics in the 21st century: A global retrospective and viewpoint analysis. J. Proteom. 2021, 241, 104220. [Google Scholar] [CrossRef]
- Windsor, P.A. Progress with livestock welfare in extensive production systems: Lessons from Australia. Front. Vet.-Sci. 2021, 8, 674482. [Google Scholar] [CrossRef]
- Lee, C.; Fisher, A. Welfare consequences of mulesing of sheep. Aust. Vet.-J. 2007, 85, 89–93. [Google Scholar] [CrossRef]
- Hancock, S.; Inglis, L.; Laurence, M.; Miller, D.; Thompson, A. Facial action units, activity and time spent with dam are effective measures of pain in response to mulesing of Merino lambs. Aust. Vet.-J. 2020, 99, 61–65. [Google Scholar] [CrossRef]
- Almeida, A.M. The Damara in the context of Southern Africa fat-tailed sheep breeds. Trop. Anim. Health Prod. 2011, 43, 1427–1441. [Google Scholar] [CrossRef] [PubMed]
- Scanlon, T.T.; Almeida, A.M.; Van Burgel, A.; Kilminster, T.; Milton, J.; Greeff, J.C.; Oldham, C. Live weight parameters and feed intake in Dorper, Damara and Australian Merino lambs exposed to restricted feeding. Small Rumin. Res. 2013, 109, 101–106. [Google Scholar] [CrossRef]
- Almeida, A.M.; Kilminster, T.; Scanlon, T.; Araújo, S.S.; Milton, J.; Oldham, C.; Greeff, J.C. Assessing carcass and meat characteristics of Damara, Dorper and Australian Merino lambs under restricted feeding. Trop. Anim. Health Prod. 2013, 45, 1305–1311. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.; Grossmann, J.; Fortes, C.; Kilminster, T.; Scanlon, T.; Milton, J.; Greeff, J.; Oldham, C.; Nanni, P.; Almeida, A.M. The sheep (Ovis aries) muscle proteome: Decoding the mechanisms of tolerance to Seasonal Weight Loss using label-free proteomics. J. Proteom. 2017, 161, 57–67. [Google Scholar] [CrossRef]
- Almeida, A.M.; Palhinhas, R.G.; Kilminster, T.; Scanlon, T.; Van Harten, S.; Milton, J.; Blache, D.; Greeff, J.; Oldham, C.; Coelho, A.V.; et al. The effect of weight loss on the muscle proteome in the Damara, Dorper and Australian Merino Ovine breeds. PLoS ONE 2016, 11, e0146367. [Google Scholar] [CrossRef]
- Palma, M.; Scanlon, T.; Kilminster, T.; Milton, J.; Oldham, C.; Greeff, J.; Matzapetakis, M.; Almeida, A.M. The hepatic and skeletal muscle ovine metabolomes as affected by weight loss: A study in three sheep breeds using NMR-metabolomics. Sci. Rep. 2016, 6, 39120. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, D.M.; Madeira, M.S.; Kilminster, T.; Scanlon, T.; Oldham, C.; Greeff, J.; Freire, J.P.B.; Mourato, M.P.; Prates, J.A.M.; Almeida, A.M. Amino acid profiles of muscle and liver tissues of Australian Merino, Damara and Dorper lambs under restricted feeding. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1295–1302. [Google Scholar] [CrossRef]
- Miller, B.; Selevsek, N.; Grossmann, J.; Kilminster, T.; Scanlon, T.; Daniels, M.; Nanni, P.; Milton, J.; Oldham, C.; Greeff, J.; et al. Ovine liver proteome: Assessing mechanisms of seasonal weight loss tolerance between Merino and Damara sheep. J. Proteom. 2019, 191, 180–190. [Google Scholar] [CrossRef]
- Miller, B.A.; Chapwanya, A.; Kilminster, T.; Scanlon, T.; Milton, J.; Osório, H.; Oldham, C.; Greeff, J.; Bergfelt, D.R.; Campos, A.; et al. The ovine hepatic mitochondrial proteome: Understanding seasonal weight loss tolerance in two distinct breeds. PLoS ONE 2019, 14, e0212580. [Google Scholar] [CrossRef]
- Alves, S.P.; Bessa, R.J.B.; Quaresma, M.A.G.; Kilminster, T.; Scanlon, T.; Oldham, C.; Milton, J.; Greeff, J.; Almeida, A.M. Does the Fat Tailed Damara Ovine Breed Have a Distinct Lipid Metabolism Leading to a High Concentration of Branched Chain Fatty Acids in Tissues? PLoS ONE 2013, 8, e77313. [Google Scholar] [CrossRef] [Green Version]
- Lérias, J.; Kilminster, T.; Scanlon, T.; Milton, J.; Oldham, C.; Greeff, J.C.; Martins, L.L.; Mourato, M.P.; Almeida, A.M. The fat-tail of Damara sheep: An assessment of mineral content as influenced by weight loss. Anim. Prod. Sci. 2016, 56, 1492–1495. [Google Scholar] [CrossRef]
- Ribeiro, D.M.; Scanlon, T.; Kilminster, T.; Martins, C.; Greeff, J.; Milton, J.; Oldham, C.; Freire, J.P.B.; Mourato, M.P.; De Almeida, A.M. Mineral profiling of muscle and hepatic tissues of Australian Merino, Damara and Dorper lambs: Effect of weight loss. J. Anim. Physiol. Anim. Nutr. 2020, 104, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.M.; Plowman, J.; Harland, D.; Thomas, A.; Kilminster, T.; Scanlon, T.; Milton, J.; Greeff, J.; Oldham, C.; Clerens, S. Influence of feed restriction on the wool proteome: A combined iTRAQ and fiber structural study. J. Proteom. 2014, 103, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Lérias, J.R.; Hernandez-Castellano, L.E.; Morales-Delanuez, A.; Araújo, S.S.; Castro, N.; Argüello, A.; Capote, J.; Almeida, A.M. Body live weight and milk production parameters in the Majorera and Palmera goat breeds from the Canary Islands: Influence of weight loss. Trop. Anim. Health Prod. 2013, 45, 1731–1736. [Google Scholar] [CrossRef] [PubMed]
- Palma, M.; Alves, S.P.; Hernández-Castellano, L.E.; Capote, J.; Castro, N.; Argüello, A.; Matzapetakis, M.; Bessa, R.J.B.; de Almeida, A.M. Mammary gland and milk fatty acid composition of two dairy goat breeds under feed-restriction. J. Dairy Res. 2017, 84, 264–271. [Google Scholar] [CrossRef]
- Lérias, J.R.; Peña, R.; E Hernández-Castellano, L.; Capote, J.; Castro, N.; Argüello, A.; Araújo, S.S.; Saco, Y.; Bassols, A.; Almeida, A.M. Establishment of the biochemical and endocrine blood profiles in the Majorera and Palmera dairy goat breeds: The effect of feed restriction. J. Dairy Res. 2015, 82, 416–425. [Google Scholar] [CrossRef]
- Parreira, J.R.; Hernández-Castellano, L.E.; Argüello, A.; Capote, J.; Castro, N.; Araújo, S.D.S.; De Almeida, A.M. Understanding seasonal weight loss tolerance in dairy goats: A transcriptomics approach. BMC Genom. 2020, 21, 629. [Google Scholar] [CrossRef]
- Hernandez-Castellano, L.E.; Ferreira, A.; Nanni, P.; Grossmann, J.; Arguello, A.; Capote, J.; Cai, G.; Lippolis, J.; Castro, N.; de Almeida, A.M. The goat (Capra hircus) mammary gland secretory tissue proteome as influenced by weight loss: A study using label free proteomics. J. Proteom. 2016, 145, 60–69. [Google Scholar] [CrossRef]
- Cugno, G.; Parreira, J.R.; Ferlizza, E.; Hernandez-Castellano, L.E.; Carneiro, M.; Renaut, J.; Castro, N.; Argűello, A.; Capote, J.; Campos, A.M.O.; et al. The Goat (Capra hircus) mammary gland mitochondrial proteome: A study on the effect of weight loss using Blue-Native PAGE and two-dimensional Gel Electrophoresis. PLoS ONE 2016, 11, e0151599. [Google Scholar] [CrossRef] [Green Version]
- Palma, M.; Hernández-Castellano, L.E.; Castro, N.; Arguëllo, A.; Capote, J.; Matzapetakis, M.; de Almeida, A.M. NMR-metabolomics profiling of mammary gland secretory tissue and milk serum in two goat breeds with different levels of tolerance to seasonal weight loss. Mol. BioSyst. 2016, 12, 2094–2107. [Google Scholar] [CrossRef]
- Zachut, M.; Šperanda, M.; de Almeida, A.M.; Gabai, G.; Mobasheri, A.; Hernández-Castellano, L.E. Biomarkers of fitness and welfare in dairy cattle: Healthy productivity. J. Dairy Res. 2020, 87, 4–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, M.T.; Vogel, C. Climate change impacts on African rangelands. Rangel. 2008, 30, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Lovarelli, D.; Bacenetti, J.; Guarino, M. A review on dairy cattle farming: Is precision livestock farming the compromise for an environmental, economic and social sustainable production? J. Clean. Prod. 2020, 262, 121409. [Google Scholar] [CrossRef]
- Fehmi, J.S.; Laca, E.A. A note on using a laser-based technique for recording of behaviour and location of free-ranging animals. Appl. Anim. Behav. Sci. 2001, 71, 335–339. [Google Scholar] [CrossRef]
- Swain, D.; Wilson, L.; Dickinson, J. Evaluation of an active transponder system to monitor spatial and temporal location of cattle within patches of a grazed sward. Appl. Anim. Behav. Sci. 2003, 84, 185–195. [Google Scholar] [CrossRef]
- Taylor, D.B.; Schneider, D.A.; Brown, W.Y.; Price, I.R.; Trotter, M.G.; Lamb, D.W.; Hinch, G.N. GPS observation of shelter utilisation by Merino ewes. Anim. Prod. Sci. 2011, 51, 724–737. [Google Scholar] [CrossRef]
- Richeson, J.T.; Lawrence, T.; White, B.J. Using advanced technologies to quantify beef cattle behavior1. Transl. Anim. Sci. 2018, 2, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Theurer, M.E.; Amrine, D.E.; White, B.J. Remote noninvasive assessment of pain and health status in cattle. Veter-Clin. North Am. Food Anim. Pract. 2013, 29, 59–74. [Google Scholar] [CrossRef] [Green Version]
- Alvarenga, F.; Borges, I.; Palkovič, L.; Rodina, J.; Oddy, V.; Dobos, R. Using a three-axis accelerometer to identify and classify sheep behaviour at pasture. Appl. Anim. Behav. Sci. 2016, 181, 91–99. [Google Scholar] [CrossRef]
- Barwick, J.; Lamb, D.W.; Dobos, R.; Welch, M.; Trotter, M. Categorising sheep activity using a tri-axial accelerometer. Comput. Electron. Agric. 2018, 145, 289–297. [Google Scholar] [CrossRef]
- Ikurior, S.J.; Marquetoux, N.; Leu, S.T.; Corner-Thomas, R.A.; Scott, I.; Pomroy, W.E. What are sheep doing? Tri-axial accelerometer sensor data identify the diel activity pattern of ewe lambs on pasture. Sensors 2021, 21, 6816. [Google Scholar] [CrossRef] [PubMed]
- McCafferty, D.J.; Gallon, S.; Nord, A.E. Challenges of measuring body temperatures of free-ranging birds and mammals. Anim. Biotelem. 2015, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Taylor, N.A.; Tipton, M.; Kenny, G.P. Considerations for the measurement of core, skin and mean body temperatures. J. Therm. Biol. 2014, 46, 72–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfrey, R.W.; Preston, W.D.; Joseph, S.R.; LaPlace, L.; Hillman, P.E.; Gebremedhin, K.G.; Lee, C.N.; Collier, R.J. Evaluating the impact of breed, pregnancy, and hair coat on body temperature and sweating rate of hair sheep ewes in the tropics1. J. Anim. Sci. 2017, 95, 2936–2942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joy, A.; Taheri, S.; Dunshea, F.; Leury, B.; DiGiacomo, K.; Osei-Amponsah, R.; Brodie, G.; Chauhan, S. Non-invasive measure of heat stress in sheep using machine learning techniques and infrared thermography. Small Rumin. Res. 2021, 207, 106592. [Google Scholar] [CrossRef]
- Baida, B.E.L.; Swinbourne, A.M.; Barwick, J.; Leu, S.T.; van Wettere, W.H.E.J. Technologies for the automated collection of heat stress data in sheep. Anim. Biotelem. 2021, 9, 4. [Google Scholar] [CrossRef]
- AlZahal, O.; Steele, M.; Van Schaik, M.; Kyriazakis, I.; Duffield, T.; McBride, B.; AlZahal, H. The use of a radiotelemetric ruminal bolus to detect body temperature changes in lactating dairy cattle. J. Dairy Sci. 2011, 94, 3568–3574. [Google Scholar] [CrossRef]
- Ipema, A.; Goense, D.; Hogewerf, P.; Houwers, H.; van Roest, H. Pilot study to monitor body temperature of dairy cows with a rumen bolus. Comput. Electron. Agric. 2008, 64, 49–52. [Google Scholar] [CrossRef]
- Beatty, D.; Barnes, A.; Fleming, P.; Taylor, E.; Maloney, S. The effect of fleece on core and rumen temperature in sheep. J. Therm. Biol. 2008, 33, 437–443. [Google Scholar] [CrossRef]
- Hanušovský, O.; Bíro, D.; Šimko, M.; Gálik, B.; Juráček, M.; Rolinec, M.; Herkeľ, R. Drinking regime evaluation with continuous ruminal monitoring boluses. Acta Fytotech. Zootech. 2017, 20, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Hyder, I.; Ravi Kanth Reddy, P.; Raju, J.; Manjari, P.; Srinivasa Prasad, C.; Aswani Kumar, K.; Seijan, V. Alteration in rumen functions and diet digestibility during heat stress in sheep. In Sheep Production Adapting to Climate Change; Sejian, V., Bhatta, R., Gaughan, J., Malik, P.K., Naqvi, S.M.K., Lal, R., Eds.; Springer Nature: Singapore, 2017; pp. 235–265. [Google Scholar]
- Ootsuka, Y.; Tanaka, M. Control of cutaneous blood flow by central nervous system. Temperature 2015, 2, 392–405. [Google Scholar] [CrossRef]
- Barros, D.V.; Silva, L.K.X.; Kahwage, P.R.; Lourenço Júnior, J.B.; Sousa, J.S.; Silva, A.G.M.; Franco, I.M.; Martorano, L.G.; Garcia, A.R. Assessment of surface temperatures of buffalo bulls (Bubalus Bubalis) raised under tropical conditions using infrared thermography. Arq. Bras. Med. Vet. Zootec. 2016, 68, 422–430. [Google Scholar] [CrossRef] [Green Version]
- Mota-Rojas, D.; Pereira, A.M.F.; Wang, D.; Martínez-Burnes, J.; Ghezzi, M.; Hernández-Avalos, I.; Lendez, P.; Mora-Medina, P.; Casas, A.; Olmos-Hernández, A.; et al. Clinical applications and factors involved in validating thermal windows used in infrared thermography in cattle and river buffalo to assess health and productivity. Animals 2021, 11, 2247. [Google Scholar] [CrossRef] [PubMed]
- Kearton, T.R.; Doughty, A.K.; Morton, C.L.; Hinch, G.N.; Godwin, I.R.; Cowley, F.C. Core and peripheral site measurement of body temperature in short wool sheep. J. Therm. Biol. 2020, 90, 102606. [Google Scholar] [CrossRef] [PubMed]
- Thornton, P.K. Livestock production: Recent trends, future prospects. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2853–2867. [Google Scholar] [CrossRef] [Green Version]
- Silanikove, N. Impact of shelter in hot Mediterranean climate on feed intake, feed utilization and body fluid distribution in sheep. Appetite 1987, 9, 207–215. [Google Scholar] [CrossRef]
- McArthur, A. Forestry and shelter for livestock. For. Ecol. Manag. 1991, 45, 93–107. [Google Scholar] [CrossRef]
- Silva-Pando, F.; Gonzalez-Hernandez, M.; Rozados-Lorenzo, M. Pasture production in a silvopastoral system in relation with microclimate variables in the atlantic coast of Spain. Agrofor. Syst. 2002, 56, 203–211. [Google Scholar] [CrossRef]
- Sousa, L.F.; Maurício, R.; Paciullo, D.S.C.; Silveira, S.; Ribeiro, R.S.; Calsavara, L.; Moreira, G. Forage intake, feeding behavior and bio-climatological indices of pasture grass, under the influence of trees, in a silvopastoral system. Trop. Grassl.-Forrajes Trop. 2015, 3, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K. Body temperatures and respiratory rates of free-ranging Merino sheep in and out of shade during summer. Aust. J. Agric. Res. 1991, 42, 1347–1357. [Google Scholar] [CrossRef]
- Joy, A.; Dunshea, F.R.; Leury, B.J.; Clarke, I.J.; Digiacomo, K.; Chauhan, S.S. Resilience of small ruminants to climate change and increased environmental temperature: A Review. Animals 2020, 10, 867. [Google Scholar] [CrossRef] [PubMed]
- Rogers, F.W. The transmission of immune globulins from the mother to the foetal and newborn young. Proc. Nutr. Soc. 1968, 28, 35–41. [Google Scholar]
- Gitlin, D.; Kumate, J.; Urrusti, J.; Morales, C. The selectivity of the human placenta in the transfer of plasma proteins from mother to fetus. J. Clin. Investig. 1964, 43, 1938–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auad, J.; Cerutti, J.; Cooper, L.G.; Lozano, N.A.; Deltrozzo, J.; Trezza, C.A.; Ponzio, M.; Lozano, A. Structure of the placenta and its impact on the transfer of maternal-fetal immunity: A review in domestic mammals. Rev. Methodo Investig. Apl. Las Cienc. Biol. 2019, 4, 52–62. [Google Scholar]
- O’Brien, J.; Sherman, D. Serum immunoglobulin concentrations of newborn goat kids and subsequent kid survival through weaning. Small Rumin. Res. 1993, 11, 71–77. [Google Scholar] [CrossRef]
- Ontsouka, C.; Bruckmaier, R.; Blum, J. Fractionized milk composition during removal of colostrum and mature milk. J. Dairy Sci. 2003, 86, 2005–2011. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Castellano, L.E.; Almeida, A.M.; Ventosa, M.; Coelho, A.V.; Castro, N.; Argüello, A. The effect of colostrum intake on blood plasma proteome profile in newborn lambs: Low abundance proteins. BMC Vet.-Res. 2014, 10, 85. [Google Scholar] [CrossRef] [Green Version]
- Agenbag, B.; Swinbourne, A.M.; Petrovski, K.; van Wettere, W.H. Lambs need colostrum: A review. Livest. Sci. 2021, 251, 104624. [Google Scholar] [CrossRef]
- Hernandez-Castellano, L.E.; Suárez-Trujillo, A.; Martell-Jaizme, D.; Cugno, G.; Arguello, A.; Castro, N. The effect of colostrum period management on BW and immune system in lambs: From birth to weaning. Animal 2015, 9, 1672–1679. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Castellano, L.; Morales-Delanuez, A.; Sánchez-Macías, D.; Moreno-Indias, I.; Torres, A.; Capote, J.; Argüello, A.; Castro, N. The effect of colostrum source (goat vs. sheep) and timing of the first colostrum feeding (2 h vs. 14 h after birth) on body weight and immune status of artificially reared newborn lambs. J. Dairy Sci. 2015, 98, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Castro, N.; Capote, J.; Bruckmaier, R.M.; Arguello, A. Management effects on colostrogenesis in small ruminants: A review. J. Appl. Anim. Res. 2011, 39, 85–93. [Google Scholar] [CrossRef]
- Nash, M.L.; Hungerford, L.L.; Nash, T.G.; Zinn, G.M. Risk factors for perinatal and postnatal mortality in lambs. Vet. Rec. 1996, 139, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Zarrin, M.; Sanginabadi, M.; Nouri, M.; Ahmadpour, A.; Hernández-Castellano, L. Prepartum and postpartum feed restrictions affect blood metabolites and hormones reducing colostrum and milk yields in fat-tailed dairy sheep. Animals 2021, 11, 1258. [Google Scholar] [CrossRef] [PubMed]
- Bielmann, V.; Gillan, J.; Perkins, N.; Skidmore, A.; Godden, S.; Leslie, K. An evaluation of Brix refractometry instruments for measurement of colostrum quality in dairy cattle. J. Dairy Sci. 2010, 93, 3713–3721. [Google Scholar] [CrossRef] [PubMed]
- Bartier, A.L.; Windeyer, M.C.; Doepel, L. Evaluation of on-farm tools for colostrum quality measurement. J. Dairy Sci. 2015, 98, 1878–1884. [Google Scholar] [CrossRef] [PubMed]
- Castro, N.; Gómez-González, L.A.; Earley, B.; Argüello, A. Use of clinic refractometer at farm as a tool to estimate the IgG content in goat colostrum. J. Appl. Anim. Res. 2018, 46, 1505–1508. [Google Scholar] [CrossRef]
- Roche, J.R.; Blache, D.; Kay, J.K.; Miller, D.R.; Sheahan, A.J.; Miller, D. Neuroendocrine and physiological regulation of intake with particular reference to domesticated ruminant animals. Nutr. Res. Rev. 2008, 21, 207–234. [Google Scholar] [CrossRef] [PubMed]
- Yaxley, K.J.; Joiner, K.F.; Abbass, H. Drone approach parameters leading to lower stress sheep flocking and movement: Sky shepherding. Sci. Rep. 2021, 11, 7803. [Google Scholar] [CrossRef]
- Johnson, P.L.; Welsh, A.; Knowler, K.; Pletnykov, P. Investigating the potential for global positioning satellite data to provide information on ewe behaviour around the time of lambing. N. Z. J. Anim. Si. Prod. 2021, 81, 29–34. [Google Scholar]
- Gurule, S.C.; Tobin, C.T.; Bailey, D.W.; Gifford, J.A.H. Evaluation of the tri-axial accelerometer to identify and predict parturition-related activities of Debouillet ewes in an intensive setting. Appl. Anim. Behav. Sci. 2021, 237, 105296. [Google Scholar] [CrossRef]
- Nowak, R.; Porter, R.H.; Levy, F.; Orgeur, P.; Schaal, B. Role of mother-young interactions in the survival of offspring in domestic mammals. Rev. Reprod. 2000, 5, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Andrews, A. Pregnancy toxaemia in the ewe. Practice 1997, 19, 306–314. [Google Scholar] [CrossRef]
- Cal-Pereyra, L.; Acosta-Dibarrat, J.; Benech, A.; Da Silva, S.; Martin, A.; González-Montaña, J.R. Toxemia de La Gestación En Ovejas. Revisión. Ewe Pregnancy Toxemia. Review. Rev. Mex. Cienc. Pecu. 2012, 3, 247–264. [Google Scholar]
- Rook, J.S. Pregnancy Toxemia of Ewes, Does, and Beef Cows. Vet.-Clin. N. Am. Food Anim. Pract. 2000, 16, 293–317. [Google Scholar] [CrossRef]
- Crilly, J.; Phythian, C.; Evans, M. Advances in managing pregnancy toxaemia in sheep. Practice 2021, 43, 79–94. [Google Scholar] [CrossRef]
- Sargison, N.D. Pregnancy Toxemia. In Diseases of Sheep; Aitken, I.D., Ed.; Blackwell Publishing Professional: Ames, LA, USA, 2007; pp. 359–363. ISBN 9781405134149. [Google Scholar]
- Kasimanickam, R.K. Subclinical Pregnancy Toxemia-Induced Gene Expression Changes in Ovine Placenta and Uterus. Front. Vet. Sci. 2016, 3, 69. [Google Scholar] [CrossRef] [Green Version]
- Lane, J.; Jubb, T.; Shephard, R.; Webb-Ware, J.; Fordyce, G. Priority List of Endemic Diseases for the Red Meat Industries; Meat and Livestock Australia Limited: Sydney, Australia, 2015; ISBN 9781741918946. [Google Scholar]
- Barbagianni, M.; Giannenas, E.; Papadopoulos, E.; Petridis, I.; Spanos, S.; Gouletsou, P.; Valasi, I.; Fthenakis, G. Pregnancy toxaemia in ewes: Development of an experimental model and potential interactions with gastrointestinal nematode infections. Small Rumin. Res. 2015, 133, 102–107. [Google Scholar] [CrossRef]
- Barbagianni, M.; Mavrogianni, V.; Katsafadou, A.; Spanos, S.; Tsioli, V.; Galatos, A.; Nakou, M.; Valasi, I.; Gouletsou, P.; Fthenakis, G. Pregnancy toxaemia as predisposing factor for development of mastitis in sheep during the immediately post-partum period. Small Rumin. Res. 2015, 130, 246–251. [Google Scholar] [CrossRef]
- Barbagianni, M.S.; Gouletsou, P.G.; Valasi, I.; Petridis, I.G.; Giannenas, I.; Fthenakis, G.C. Ultrasonographic findings in the ovine udder during lactogenesis in healthy ewes or ewes with pregnancy toxaemia. J. Dairy Res. 2015, 82, 293–303. [Google Scholar] [CrossRef]
- Santos, F.C.O.; Mendonça, C.L.; Silva Filho, A.P.; Carvalho, C.C.D.; Soares, P.C.; Afonso, J.A.B. Indicadores Bioquímicos e Hormonais de Casos Naturais de Toxemia Da Prenhez Em Ovelhas. Pesqui. Vet. Bras. 2011, 31, 974–980. [Google Scholar] [CrossRef] [Green Version]
- Kabakci, N.; Yarim, G.; Yarim, M.; Duru, O.; Yagci, B.B.; Kisa, U. Pathological, clinical and biochemical investigation of naturally occurring pregnancy toxemia of sheep. Acta Vet. 2003, 53, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Karagiannis, I.; Panousis, N.; Kiossis, E.; Tsakmakidis, I.; Lafi, S.; Arsenos, G.; Boscos, C.; Brozos, C. Associations of pre-lambing body condition score and serum β-hydroxybutyric acid and non-esterified fatty acids concentrations with periparturient health of Chios dairy ewes. Small Rumin. Res. 2014, 120, 164–173. [Google Scholar] [CrossRef]
- Cal-Pereyra, L.; Benech, A.; González-Montaña, J.R.; Dibarrat, J.A.; Da Silva, S.; Martín, A. Changes in the metabolic profile of pregnant ewes to an acute feed restriction in late gestation. N. Z. Vet. J. 2015, 63, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Aly, M.; Elshahawy, I. Clinico-Biochemical Diagnosis of Pregnancy Toxemia in Ewes with Special Reference to Novel Biomarkers. Alex. J. Vet. Sci. 2016, 48, 96. [Google Scholar] [CrossRef]
- Albay, M.; Karakurum, M.; Sahinduran, S.; Sezer, K.; Yildiz, R.; Buyukoglu, T. Selected serum biochemical parameters and acute phase protein levels in a herd of Saanen goats showing signs of pregnancy toxaemia. Vet. Med. (Praha). 2014, 59, 336–342. [Google Scholar] [CrossRef] [Green Version]
- Pethick, D.W.; Lindsay, D.B. Metabolism of ketone bodies in pregnant sheep. Br. J. Nutr. 1982, 48, 549–563. [Google Scholar] [CrossRef] [Green Version]
- Amirul, F.M.A.; Mokrish, A.; Lai, K.S.; Zamri-Saad, M.; Zuki, A.B.; Hassim, H.A. Clinical, Biochemical and Histological Changes during Developement of Pregnancy Ketosis in Goats. J. Vet. Malays. 2016, 28, 1–6. [Google Scholar]
- Scott, P. Analysis of Cerebrospinal Fluid from Field Cases of Some Common Ovine Neurological Diseases. Br. J. Nutr. 1992, 148, 15–22. [Google Scholar] [CrossRef]
- Scott, P. Differential Diagnosis of Common Metabolic Disorders of Sheep. Practice 1995, 17, 266–269. [Google Scholar] [CrossRef]
- Yarim, G.F.; Ciftci, G. Serum Protein Pattern in Ewe with Pregnancy Toxemia. Vet. Res. Commun. 2009, 33, 431–438. [Google Scholar] [CrossRef]
- Barakat, S.E.M.; Al-Bhanasawi, N.M.; Elazhari, G.E.; Bakhiet, A.O. Clinical and Serobiochemical Studies on Naturally-Occurring Pregnancy Toxaemia in Shamia Goats. J. Anim. Vet. Adv. 2007, 6, 768–772. [Google Scholar]
- Hefnawy, A.-E.; Youssef, S.; Shousha, S. Some Immunohormonal Changes in Experimentally Pregnant Toxemic Goats. Vet. Med. Int. 2010, 2010, 768438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cal, L.; Borteiro, C.; Benech, A.; Rodas, E.; Abreu, M.N.; Cruz, J.C.; González-Montaña, J.R. Alterações Histológicas do Fígado e Correlações Metabólicas em Ovelhas Com Toxemia da Prenhez (Histological Changes of the Liver and Metabolic Correlates in Ewes with Pregnancy Toxemia). Arq. Bras. Med. Vet. Zootec. 2009, 61, 306–312. [Google Scholar] [CrossRef]
- González, F.H.D.; Hernández, F.; Madrid, J.; Martinez-Subiela, S.; Tvarijonaviciute, A.; Cerón, J.J.; Tecles, F. Acute Phase Proteins in Experimentally Induced Pregnancy Toxemia in Goats. J Vet Diagn Investig. 2011, 23, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarım, G.F.; Karahan, S.; Nisbet, C. Elevated Plasma Levels of Interleukin 1β, Tumour Necrosis Factor α and Monocyte Chemotactic Protein 1 Are Associated with Pregnancy Toxaemia in Ewes. Vet. Res. Commun. 2007, 31, 565–573. [Google Scholar] [CrossRef] [PubMed]
- de Souza, L.; Mendonça, C.; Assis, R.; Filho, E.O.; Soares, G.; Souto, R.; Soares, P.; Afonso, J. Changes in cardiac biomarkers in goats naturally affected by pregnancy toxemia. Res. Vet. Sci. 2020, 130, 73–78. [Google Scholar] [CrossRef]
- de Souza, L.M.; de Mendonça, C.L.; de Assis, R.N.; Filho, E.F.O.; Gonçalves, D.N.A.; Souto, R.J.C.; Soares, P.C.; Afonso, J.A.B. Cardiac Biomarkers Troponin I and Ck-Mb in Ewes Affected by Pregnancy Toxemia. Small Rumin. Res. 2019, 177, 97–102. [Google Scholar] [CrossRef]
- Pichler, M.; Damberger, A.; Schwendenwein, I.; Gasteiner, J.; Drillich, M.; Iwersen, M. Thresholds of whole-blood β-hydroxybutyrate and glucose concentrations measured with an electronic hand-held device to identify ovine hyperketonemia. J. Dairy Sci. 2014, 97, 1388–1399. [Google Scholar] [CrossRef]
- Araújo, C.A.S.C.; Minervino, A.H.H.; Sousa, R.S.; Oliveira, F.L.C.; Rodrigues, F.A.M.L.; Mori, C.S.; Ortolani, E.L. Validation of a handheld β-hydroxybutyrate acid meter to identify hyperketonaemia in ewes. PeerJ 2020, 8, e8933. [Google Scholar] [CrossRef]
- Sykes, A.R.; Coop, R.L. Interaction between nutrition and gastrointestinal parasitism in sheep. N. Z. Vet. J. 2001, 49, 222–226. [Google Scholar] [CrossRef]
- Vineer, H.R.; Morgan, E.R.; Hertzberg, H.; Bartley, D.J.; Bosco, A.; Charlier, J.; Chartier, C.; Claerebout, E.; De Waal, T.; Hendrickx, G.; et al. Increasing importance of anthelmintic resistance in European livestock: Creation and meta-analysis of an open database. Parasite 2020, 27, 62. [Google Scholar] [CrossRef]
- Scott, I.; Pomroy, W.; Kenyon, P.; Smith, G.; Adlington, B.; Moss, A. Lack of efficacy of monepantel against Teladorsagia circumcincta and Trichostrongylus colubriformis. Vet. Parasitol. 2013, 198, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Athanasiadou, S.; Gray, D.; Younie, D.; Tzamaloukas, O.; Jackson, F.; Kyriazakis, I. The use of chicory for parasite control in organic ewes and their lambs. Parasitology 2006, 134, 299–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thamsborg, S.M.; Roepstorff, A. Parasite problems in organic livestock and options for control. J. Parasitol. 2003, 89, 277–284. [Google Scholar]
- Kakamoukas, G.; Sarigiannidis, P.; Maropoulos, A.; Lagkas, T.; Zaralis, K.; Karaiskou, C. Towards Climate Smart Farming—A Reference Architecture for Integrated Farming Systems. Telecom 2021, 2, 5. [Google Scholar] [CrossRef]
- Ikurior, S.J.; Pomroy, W.E.; Scott, I.; Corner-Thomas, R.; Marquetoux, N.; Leu, S.T. Gastrointestinal nematode infection affects overall activity in young sheep monitored with tri-axial accelerometers. Vet. Parasitol. 2020, 283, 109188. [Google Scholar] [CrossRef]
- Högberg, N.; Hessle, A.; Lidfors, L.; Baltrušis, P.; Claerebout, E.; Höglund, J. Subclinical nematode parasitism affects activity and rumination patterns in first-season grazing cattle. Animal 2021, 15, 100237. [Google Scholar] [CrossRef]
- Vercruysse, J.; Charlier, J.; Van Dijk, J.; Morgan, E.; Geary, T.; Von Samson-Himmelstjerna, G.; Claerebout, E. Control of helminth ruminant infections by 2030. Parasitology 2018, 145, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- Hoste, H.; Martinez-Ortiz-De-Montellano, C.; Manolaraki, F.; Brunet, S.; Ojeda-Robertos, N.; Fourquaux, I.; Torres-Acosta, J.; Sandoval-Castro, C. Direct and indirect effects of bioactive tannin-rich tropical and temperate legumes against nematode infections. Vet. Parasitol. 2012, 186, 18–27. [Google Scholar] [CrossRef]
- Athanasiadou, S.; Kyriazakis, I. Plant secondary metabolites: Antiparasitic effects and their role in ruminant production systems. Proc. Nutr. Soc. 2004, 63, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Kommuru, D.; Whitley, N.; Miller, J.; Mosjidis, J.; Burke, J.; Gujja, S.; Mechineni, A.; Terrill, T. Effect of sericea lespedeza leaf meal pellets on adult female Haemonchus contortus in goats. Vet. Parasitol. 2015, 207, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Jackson, F.; Miller, J. Alternative approaches to control—Quo vadit? Vet. Parasitol. 2006, 139, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Tzamaloukas, O.; Athanasiadou, S.; Kyriazakis, I.; Huntley, J.F.; Jackson, F. The effect of chicory (Cichorium intybus) and sulla (Hedysarum coronarium) on larval development and mucosal cell responses of growing lambs challenged withTeladorsagia circumcincta. Parasitology 2005, 132, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Judson, G.; Brown, T.; Gray, D.; Dewey, D.; Edwards, J.; McFarlane, J. Oxidized copper wire particles for copper therapy in sheep. Aust. J. Agric. Res. 1982, 33, 1073–1083. [Google Scholar] [CrossRef]
- Knox, M.R. Effectiveness of copper oxide wire particles for Haemonchus contortus control in sheep. Aust. Vet. J. 2002, 80, 224–227. [Google Scholar] [CrossRef] [PubMed]
- Bang, K.; Familton, A.; Sykes, A. Effect of copper oxide wire particle 534 treatment on establishment of major gastrointestinal nematodes in lambs. Res. Vet. Sci. 1990, 49, 132–137. [Google Scholar] [CrossRef]
- Burke, J.; Miller, J.; Terrill, T.; Smyth, E.; Acharya, M. Examination of commercially available copper oxide wire particles in combination with albendazole for control of gastrointestinal nematodes in lambs. Vet. Parasitol. 2016, 215, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Burke, J.; Miller, J.; Mosjidis, J.; Terrill, T. Use of a mixed sericea lespedeza and grass pasture system for control of gastrointestinal nematodes in lambs and kids. Vet. Parasitol. 2012, 186, 328–336. [Google Scholar] [CrossRef]
- Vatta, A.; Waller, P.; Githiori, J.; Medley, G. Persistence of the efficacy of copper oxide wire particles against Haemonchus contortus in grazing South African goats. Vet. Parasitol. 2012, 190, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Suttle, N. Safety and effectiveness of cupric oxide particles for increasing liver copper stores in sheep. Res. Vet. Sci. 1987, 42, 219–223. [Google Scholar] [CrossRef]
- Burke, J.M.; Miller, J.E. Sustainable Approaches to Parasite Control in Ruminant Livestock. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 89–107. [Google Scholar] [CrossRef] [PubMed]
- Zaralis, K.; Tolkamp, B.J.; Houdijk, J.G.M.; Wylie, A.R.G.; Kyriazakis, I. Consequences of protein supplementation for anorexia, expression of immunity and plasma leptin concentrations in parasitized ewes of two breeds. Br. J. Nutr. 2008, 101, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Crawford, C.D.; Mata-Padrino, D.J.; Belesky, D.P.; Bowdridge, S.A. Effects of supplementation containing rumen by-pass protein on parasitism in grazing lambs. Small Rumin. Res. 2020, 190, 160191. [Google Scholar] [CrossRef]
- Houdijk, J.G. Differential effects of protein and energy scarcity on resistance to nematode parasites. Small Rumin. Res. 2012, 103, 41–49. [Google Scholar] [CrossRef]
- Bishop, S. Possibilities to breed for resistance to nematode parasite infections in small ruminants in tropical production systems. Animal 2012, 6, 741–747. [Google Scholar] [CrossRef] [Green Version]
- Stear, M.J.; Fitton, L.; Innocent, G.T.; Murphy, L.; Rennie, K.; Matthews, L. The dynamic influence of genetic variation on the susceptibility of sheep to gastrointestinal nematode infection. J. R. Soc. Interface 2007, 4, 767–776. [Google Scholar] [CrossRef]
- Bisset, A.S.; Morris, A.C.; McEwan, J.; Vlassoff, A. Breeding sheep in N. Z. that are less reliant on anthelmintics to maintain health and productivity. N. Z. Vet. J. 2001, 49, 236–246. [Google Scholar] [CrossRef]
- Pernthaner, A.; Cole, S.-A.; Morrison, L.; Hein, W.R. Increased expression of Interleukin-5 (IL-5), IL-13, and tumor necrosis factor alpha genes in intestinal lymph cells of sheep selected for enhanced resistance to nematodes during infection with Trichostrongylus colubriformis. Infect. Immun. 2005, 73, 2175–2183. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.; Faedo, M.; Waller, P.; Hennessy, D. The potential of nematophagous fungi to control the free-living stages of nematode parasites of sheep: Studies with Duddingtonia flagrans. Vet. Parasitol. 1998, 76, 121–128. [Google Scholar] [CrossRef]
- Peña, M.; Miller, J.; Fontenot, M.; Gillespie, A.; Larsen, M.H. Evaluation of Duddingtonia flagrans in reducing infective larvae of Haemonchus contortus in feces of sheep. Vet. Parasitol. 2001, 103, 259–265. [Google Scholar] [CrossRef]
- Chandrawathani, P.; Omar, J.; Waller, P. The control of the free-living stages of Strongyloides papillosus by the nematophagous fungus, Arthrobotrys oligospora. Vet. Parasitol. 1998, 76, 321–325. [Google Scholar] [CrossRef]
- Hoste, H.; Torres-Acosta, J.F.D.J. Non chemical control of helminths in ruminants: Adapting solutions for changing worms in a changing world. Vet. Parasitol. 2011, 180, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Saumell, C.; Fernández, A.; Echevarria, F.; Gonçalves, I.; Iglesias, L.; Sagües, M.; Rodríguez, E. Lack of negative effects of the biological control agent Duddingtonia flagrans on soil nematodes and other nematophagous fungi. J. Helminthol. 2015, 90, 706–711. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M. Biological control of nematode parasites in sheep1. J. Anim. Sci. 2006, 84, E133–E139. [Google Scholar] [CrossRef] [PubMed]
- Bampidis, V.; Azimonti, G.; Bastos, M.L.; Christensen, H.; Dusemund, B.; Kos Durjava, M.; Kouba, M.; López-Alonso, M.; López Puente, S.; Marcon, F.; et al. Scientific Opinion on the safety and efficacy of BioWorma® (Duddingtonia flagrans NCIMB 30336) as a feed additive for all grazing animals. EFSA J. 2020, 18, 6208. [Google Scholar] [CrossRef]
- Barger, I.; Siale, K.; Banks, D.; Le Jambre, L. Rotational grazing for control of gastrointestinal nematodes of goats in a wet tropical environment. Vet. Parasitol. 1994, 53, 109–116. [Google Scholar] [CrossRef]
- Rocha, R.; Bresciani, K.; Barros, T.; Fernandes, L.; Silva, M.; Amarante, A. Sheep and cattle grazing alternately: Nematode parasitism and pasture decontamination. Small Rumin. Res. 2008, 75, 135–143. [Google Scholar] [CrossRef]
- Besier, R. Refugia-based strategies for sustainable worm control: Factors affecting the acceptability to sheep and goat owners. Vet. Parasitol. 2012, 186, 2–9. [Google Scholar] [CrossRef]


{kind=link}
| WMT | n/Breed | Aim | Technique | Results | Ref. |
|---|---|---|---|---|---|
| B | Cheviot ewes | Determine if different behavior types associated with grazing | Sensor accelerometer-integrated collars | Accuracy 90% | [22] |
| B | 29 Scottish Blackface ewes | General activity and circadian rhythm of activity with sheep body weight change | Sensor accelerometer-integrated collars | [60] | |
| B | 3 Merino | Behavioral and movement patterns of individuals | Tri-axial sensors, temperature sensor, and GPS | Accuracy > 75% | [61] |
| B | 6 sheep | Continuous surveillance of eating behavior for monitoring ruminant health, productivity, and welfare | Tri-axial gyroscope and tri-axial accelerometer | Accuracy > 86% | [62] |
| B | 50 Merino | Low-cost solution to monitoring of the location of all the animals in a herd and the continuous updating of location data | GPS collars (25 ewes) and BLE (25 ewes) | [63] | |
| B | 10 Norwegian White | System that automatically generated individual animal behavior and localization | Real-time sensor tags and tri-axial accelerometers (ST LIS2DE) | SE = 98.16% (standing); SE = 100% (lying) | [64] |
| B | Serra da Estrela breed | Autonomous system to control sheep posture and monitor their location in real-time. | Collar set of sensors (inertial and ultrasound) and a microcontroller and actuators (i.e., stimulation devices, namely sound and electrostatic) | [65] | |
| La | 40 ewes | Predictive model to identify the day of lambing in extensive sheep | GNSS tracking collars | Accuracy 83.0%, SE = 63.6%, SP = 84.1% | [66] |
| La | 39 Merino | Monitor changes in sheep behavior around the time of lambing | Accelerometer ear-tags (Axivity AX3) | [67] | |
| L | 10 Merino Poll Dorset ewes | Ability of a tri-axial accelerometer to discriminate between sound and lame gait | Accelerometers (GCDC X16) on 3 points: neck collar, ear, and leg | Accuracy 82% (ear), 35% (collar), and 87% (leg) | [68] |
| L | 20 various breeds | Relationship between sheep hoof-health status and the load a sheep distributes to each hoof | Hoof weigh crate raceway two strain-gauge cantilever load cells | SE = 100%, SP = 95% | [69] |
| LW | 4 flocks | LW as an indicator of nutritional status | WoW | Repeatability 0.20–0.76 | [70] |
| LW | 900 ewes | Ewe performance of two different methods of feed allocation | Automatic weigh and drafting crates coupled with EID technology | Accuracy 52% | [71] |
| LW | Romane ewes | LW data were recorded as each ewe entered voluntarily and walked throughout the WoW | WoW | Accuracy 0.89 and 0.98 | [72] |
| Parameter | Reference Interval and Problematic Values | References |
|---|---|---|
| Glycemia | 50–70 mg/dL (2.8–3.9 mmol/L); falls below 20 mg/dL | [155,158,165,168,169] |
| Blood ketone bodies (especially ß-hydroxybutyrate (BOHB) | <1.1 mmol/L; | [154,155,156,158] |
| >2 mmol/L (36.03 mg/dL); | [170] | |
| >5.0 mmol/L (90.09 mg/dL); | [154,158,159,164,165] | |
| 19.0 mmol/l. | [171] | |
| BOHB in aqueous humor or cerebrospinal fluid | >2.5 mmol/L (45.0 mg/dL) y > 0.5 mmol/L (9.0 mg/dL), respectively. | [172,173] |
| Total proteinemia, albuminemia and globulins in blood | Significant fall, due to liver injury and anorexia; possible false increase due to dehydration. | [164,168,174] |
| Urea, blood urea nitrogen (BUN) and serum creatinine | Increased urea, blood urea nitrogen (BUN), and serum creatinine, due to renal dysfunction. | [154,155,168,175] |
| Serum cortisol and thyroid hormones (T3 and T4) | Increased cortisol, justified by hyperactive adrenal glands. | [164,168] |
| Decreased T3 and T4, due to hypersecretion of cortisol. | [168,176] | |
| Blood enzymes: aspartate aminotransferase (AST); alanine aminotransferase (ALT); gamma glutamyl transferase (GGT); lactate dehydrogenase (LDH) and creatine kinase (CK) | Increased AST, ALT, GGT, LDH, and CK, possibly due to liver dysfunction. Positive correlation between AST activity and the degree of hepatic vacuolization, a consequence of hepatic steatosis, therefore this enzyme could be used as an early and reliable indicator of liver damage in sheep with clinical pregnancy toxemia [177] | AST: [155,164,171,174,175,177] ALT and GGT: [168,174,175] LDH: [168] CK: [164,175] |
| Parameter | Modification of the Values and Justification | References |
|---|---|---|
| Fructosamine and glycosylated hemoglobin | Both indicate not the current glycemia, but over a long period prior to measurement; low values suggest persistent hypoglycemia. | [154,164] |
| Minerals: potassium (K), sodium (Na), calcium (Ca) and phosphorus (P) in blood | Large decrease in K, Na and Ca. | [168] |
| No changes in phosphorus. | [168] | |
| Due by starvation, dehydration, metabolic acidosis, electrolyte imbalance and renal dysfunction, as well as increased lipolysis that can induce hypocalcemia. | [156,168] | |
| High calcium demand of late gestation leads to a significant decrease in maternal calcemia. | [154] | |
| Metabolic acidosis and aciduria | Metabolic acidosis (lactate and pyruvate measured in blood). | [164,175] |
| Aciduria, measured in urine using semi-quantitative test strips. | [164,165] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, S.R.; Sacarrão-Birrento, L.; Almeida, M.; Ribeiro, D.M.; Guedes, C.; González Montaña, J.R.; Pereira, A.F.; Zaralis, K.; Geraldo, A.; Tzamaloukas, O.; et al. Extensive Sheep and Goat Production: The Role of Novel Technologies towards Sustainability and Animal Welfare. Animals 2022, 12, 885. https://doi.org/10.3390/ani12070885
Silva SR, Sacarrão-Birrento L, Almeida M, Ribeiro DM, Guedes C, González Montaña JR, Pereira AF, Zaralis K, Geraldo A, Tzamaloukas O, et al. Extensive Sheep and Goat Production: The Role of Novel Technologies towards Sustainability and Animal Welfare. Animals. 2022; 12(7):885. https://doi.org/10.3390/ani12070885
Chicago/Turabian StyleSilva, Severiano R., Laura Sacarrão-Birrento, Mariana Almeida, David M. Ribeiro, Cristina Guedes, José Ramiro González Montaña, Alfredo F. Pereira, Konstantinos Zaralis, Ana Geraldo, Ouranios Tzamaloukas, and et al. 2022. "Extensive Sheep and Goat Production: The Role of Novel Technologies towards Sustainability and Animal Welfare" Animals 12, no. 7: 885. https://doi.org/10.3390/ani12070885