1. Introduction
High ambient temperature is a primary problem in broiler breeding, since it has a negative impact on performance and other traits that affect profits [
1]. Evidence in several studies indicates that glucose homeostasis can be influenced under heat stress in animals [
2,
3], including changes in blood glucose, insulin, and NEFA [
4,
5,
6]. More seriously, when target tissues cannot respond well to insulin and the insulin level becomes higher than the level of glucose in the circulation, animals develop insulin resistance as a result. A previous study observed a state of insulin resistance in human hyperthermia [
7]. Our previous research found that heat stress caused insulin resistance in broilers [
8]. High temperature leads to reduced glucose absorption efficiency. Recent evidence suggests that excessive temperature affects glucose transporter expression in broilers [
9]. Furthermore, excessive temperature impacts the composition of the gut microbial community and gastrointestinal peptides. Our previous research suggested that high temperature significantly alters the proportion of bacteria and abundance of metabolites and that the concentrations of several gastrointestinal peptides changed owing to excessive temperature [
8].
Chromium is a trace nutrient and an important active component of the glucose tolerance factor, which regulates the metabolism of carbohydrates, fat, and protein [
10]. It has been confirmed that heat stress will increase the excretion of chromium in broilers, and that chromium deficiency will lead to an imbalance in glucose homeostasis, as well as blunt glucose response [
11,
12]. Numerous research studies have indicated that dietary trivalent chromium can alleviate the adverse impact caused by heat stress [
13,
14,
15]. Trivalent chromium could improve insulin sensitivity and glucose utilization in animals and humans who suffer from metabolic diseases such as type 2 diabetes and obesity [
16,
17]. In several studies, the glucose transporters in heat-stressed poultry could be improved by supplementing their diets with organic chromium, which is conducive to maintaining glucose homeostasis [
18,
19,
20]. However, there has been no report about the influence of chromium on glucose homeostasis involved in the microflora in heat-stressed chickens.
Thus, the objective of this paper was to assess the influence of chromium picolinate supplementation on gut microbiota, glucose transporters, gastrointestinal peptides, glucose homeostasis, and performance in broilers raised under heat stress, then, to explore the effect of chromium picolinate on alleviating heat stress through the microbiota and provide a new regulatory target for alleviating heat stress in broilers.
2. Materials and Methods
2.1. Full Name and Description of Abbreviations
For abbreviations, see the abbreviations list presented in
Table 1.
2.2. Animals
A total of 270 male 1-day-old AA broilers were accessed from a local hatchery and reared in breeding cages (three layers, 8400 cm
2 per layer) according to AA broiler breeding management requirements. All birds were provided access to water and feed ad libitum. A total of 220 birds were selected according to average weight, then randomly assigned to four groups with five replicates (11 chickens/pen) according to individual body weight. The treatment group design is shown in
Table 2. The adaptation period was entered at the age of 21 days, and the trial began at the age of 28 days and lasted from 14 days to 42 days. Ambient temperature and humidity control were adjusted by real-time temperature and humidity control chambers developed by our Institute. The chamber with one door has good thermal insulation performance. The average body weight of the four groups of broilers was similar. The temperature in the chambers of each group was constant, and the humidity was maintained at 60% during the experiment. The broilers were fed on a crumble diet that was selected to meet the NRC nutrient recommendations (1994,
Table 3). Birds were provided with continuous light (20 lux, 18 h light, 6 h dark) throughout the experimental period. To reduce stress, the number of times in and out of the chamber was minimized. Ethical approval was obtained based on the animal welfare and ethics checklist of the institute of animal science of the Chinese academy of agricultural sciences (ethics code number: IAS 2021-75).
2.3. Samples and Data Collection
A scale with accuracy to 0.01 g was used to measure the body weight of birds in each replicate. The ADFI, ADG, and FCR for each replicate were calculated at the end of the trial. One bird was randomly selected out of the 11 birds in each replicate for further sampling; serum was collected and stored in a refrigerator for determination of the levels of ghrelin, CCK, GIP, and other parameters. When serum sample collection was completed, the birds with blood samples taken were immediately humane-sacrificed and the cecal contents, hypothalamus, and intestinal mucosa were obtained. The samples were stored in a −80 °C refrigerator. Another chicken was randomly selected from each replicate after 12 h fasting for insulin and insulin resistance index determination.
2.4. Determination of Gastrointestinal Peptides
ELISA was used to measure the contents of CCK, ghrelin, and GIP in the hypothalamus, intestinal mucosa, and serum. The intestinal mucosa was ground into a powdery form, diluted with PBS (1:9), then centrifuged at 2000–3000 rpm for 20 min; the supernatant was collected for further determination. Standard wells, sample wells, and blank wells were set, then 50 μL serum samples were added into each sample well. The diluent and enzyme label reagent were successively added and incubated at 37 °C for 60 min. A washing solution was used to wash the ELISA plate, then a color reagent and stop solution were added, and eventually the enzyme-labeled instrument (Bio-tek, Gen 5 1.10) was used to determine the absorbance of each well. The levels of peptides were calculated according to the standard curve.
2.5. Analysis of the Cecal Microbiota
DNA of microbiota was extracted from cecal content samples using the E.Z.N.A.® soil DNA Kit (Omega Bio-tek, Norcross, GA, USA). The hypervariable region V3-V4 of the 16S rRNA gene was amplified with primer pairs 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTC-TAAT-3′). The PCR amplification procedure was as follows: 95 °C for 3 min, 27 cycles at 95 °C for 30 s, 55 °C for 30 s, 72 °C for 45 s, 72 °C for 10 min, ending at 10 °C. The PCR product was extracted from 2% agarose gel and purified. Purified amplicons were pooled in equimolar ratios and paired-end sequenced on an Il-lumina MiSeq PE300 platform/NovaSeq PE250 platform (Illumina, San Diego, CA, USA) according to the standard protocols.
2.6. Determination of SCFAs
One gram of digesta from the cecum samples was weighed accurately. The samples after dilution with ultra-pure water were subjected to short-chain acid analysis by gas chromatography. Chromatographic conditions were as follows. Column: db-ffap (30 m × 250 μm × 0.25 μm), flow rate: 0.8 mL/min, auxiliary gas: high purity hydrogen (99.999%), the temperature of detector FID: 280 °C, temperature of injection port: 250 °C, split ratio: 50:1, injection volume: 1 μL, temperature programming: 60 °C→220 °C (20 °C/min).
2.7. Determination of Glucose Transporter Gene Expression
The expression levels of SGLT1 and GLUT2 in the jejunum were measured using qPCR. The general total RNA extraction kit (Hooseen biology, Beijing, China) was used to extract RNA from tissue samples, and the experimental operation was carried out according to the product instructions. Real-time PCR was conducted using the Line Gene 9620 Real-time PCR System (Bioer, Hangzhou, China). Primer-set sequences are provided in
Table 4. Real-time PCR reactions were conducted at 95 °C for 10 min, followed by 40 cycles of 95 °C for 20 s, 55 °C for 20 s and 72 °C for 20 s. The mRNA levels were normalized to glyceraldehyde-3-phosphate dehydrogenase levels (ΔCt). The relative gene expression amount of each sample was calculated according to the formula for relative quantities: RQ = 2
−ΔΔCt.
2.8. Determination of Glucose Homeostasis and Other Serum Parameters
Glucose homeostasis determination was performed by insulin resistance index, using the formula insulin resistance index = insulin/(22.5e-ln glucose) to calculate the insulin resistance index [
21]. The levels of insulin, NEFA, TG, and TC were determined using the kits provided by Nanjing Jiancheng Bioengineering Institute (Jiangsu, China).
2.9. Statistical Analysis
All data were analyzed using GraphPad Prism 8 software (GraphPad Prism 8.3.0 for Mac OS, GraphPad Software, Inc., La Jolla, CA, USA). One-way ANOVA followed by post hoc (Bonferroni’s multiple comparison test) analysis was used for the comparison of four treatments. The confidence interval of 95% was considered, p < 0.05 means a significant difference, and values are presented as the mean.
4. Discussion
Trivalent chromium has a positive effect on livestock under heat stress [
13,
22,
23]. It has been reported that diets supplemented with 0.2 mg/kg Cr as Cr-Pic and Cr-His both significantly increased feed intake and body weight gain in heat-stressed broilers [
24]. In the current study, the ADFI and ADG significantly increased in the 0.4 mg/kg supplemented group compared with the HT group, indicating that dietary supplementation with 0.4 mg/kg Cr as Cr-Pic can reduced the severity of impaired performance under heat stress. This corresponds with previous results indicating that Cr-Pic has a positive effect on growth performance in broilers under heat stress [
25,
26].
Several critical factors in glucose homeostasis include intestinal glucose uptake, peripheral tissue glucose uptake and utilization, and liver glucose output efficiency. Insulin is an important factor in controlling glucose homeostasis. Insulin resistance will lead to an imbalance in glucose homeostasis, abnormal changes in blood glucose, and even lead to metabolic diseases such as diabetes mellitus with hyperinsulinemia. Trivalent chromium has been proved to regulate glucose metabolism in coordination with insulin, which is conducive to maintaining glucose homeostasis [
27]. In the current study, compared with the HT group, the serum insulin concentration and insulin resistance index of the 0.4 mg/kg group were significantly reduced. Chromium is a cofactor of insulin activity and an essential element for glucose utilization [
28]. This may be due to increased insulin sensitivity resulting from chromium supplementation. Therefore, it indicated that 0.4 mg/kg Cr as Cr-Pic supplement was beneficial to relieve the insulin resistance caused by heat stress.
The intestinal microbiota is the largest and most complex micro-ecosystem in animals. The microflora are not only involved in a variety of metabolic pathways to regulate metabolism, but also play an important role as a mediator between the diet and the host. Undoubtedly, diet is a factor that influences the composition of the intestinal microbiota. In the 0.4 mg/kg chromium supplemented group of the current study, at the class level, the proportion of Coriobacteria decreased. In a previous study, it was mentioned that an increased proportion of Coriobacteria will increase the risk of insulin resistance and obesity [
29]. At the family level, the proportions of Christensenellaceae, Clostridiaceae, and Erysipelotrichaceae increased; these bacteria were reported to be beneficial to normal glucose metabolism, alleviate insulin resistance, and lower the risk of obesity [
30,
31]. Our previous study showed that the abundance of Christensenellaceae was significantly reduced by heat stress, while the abundance of Christensenellaceae was significantly increased in the 0.4 mg/kg supplemented group [
8], indicating that glucose metabolism in the chromium supplement group improved. At the genus level, the proportion of Ruminococcus decreased. Previous research suggests that microbiota can induce insulin resistance, and Ruminococcus is one of them [
32]. Therefore, these alterations in microflora suggest that 0.4 mg/kg supplementation was beneficial to alleviate insulin resistance and improve glucose homeostasis.
The absorption of carbohydrates in the gut is carried out by glucose transporters. In poultry, the duodenum and jejunum are the main sites for the absorption and uptake of glucose [
33]. SGLT1 transfers glucose to the epithelial cells of the small intestine through active transport. After intracellular transport, glucose is transported to all parts of the body through GLUT2 in the basement membrane in a diffusion-assisted manner [
34]. GLUT2 is more sensitive when the body is hyperglycemic, which helps the liver to efficiently regulate the blood glucose level [
34]. In addition, SGLT1 and GLUT2 play a vital role in incretin secretion [
35]. The importance of the SGLT1 transporter in sodium-glucose homeostasis was confirmed by the disappearance of intestinal glucose uptake in SGLT1-deficient mice [
36]. The current study suggested that the level of SGLT1 in the jejunum in the 0.4 mg/kg supplemented group was significantly up-regulated. These discoveries are consistent with those of Cemal Orhan et al., who cited a significant increase in SGLT1 expression in ileum of laying hens by supplementation with Cr-Pic [
19]. Chromium factors are involved in glucose metabolism under heat stress; these forms of chromium can not be reabsorbed, leading to a lack of chromium and reduced efficiency of glucose absorption in the intestine. Thus, the improvement in SGLT1 expression could be due to the increase in chromium levels in the intestine from diet-supplemented chromium. Kashiwagi et al. also found decreased glucose transporters in insulin-resistant patients with type II diabetes [
37]. Insulin resistance is associated with the efficiency of glucose absorption and transport. Therefore, the improvement in insulin resistance may also be related to the up-regulation of SGLT1 expression. Whether gut microbiota modulates the expression of nutrient-responsive receptors and transporters remains unclear. A previous study found up-regulation of SGLT1 and GLUT2 expression in mice lacking gut microbiota [
38], implying that the microbiota could be a potential regulator of glucose homeostasis. This may be worth pursuing in future research.
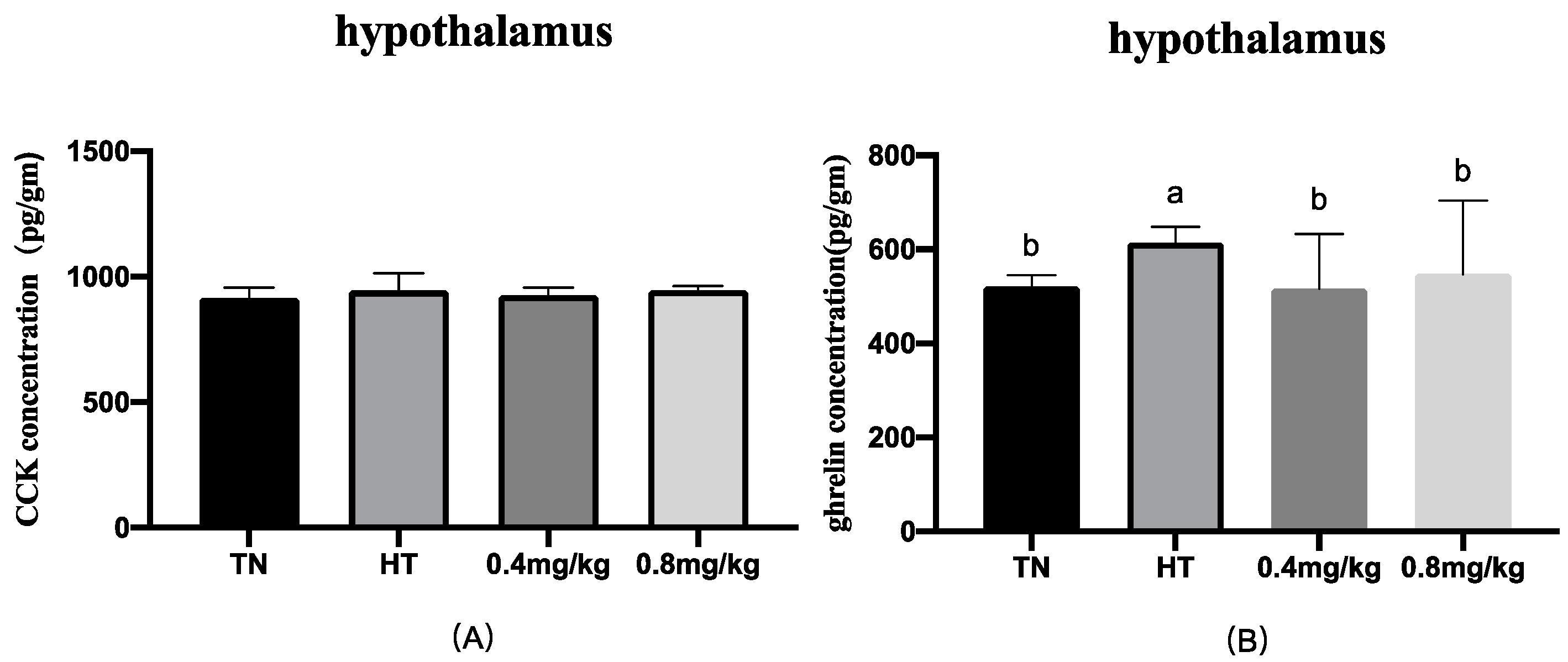
The peptides in the gut are produced by specific enteroendocrine cells. The gut releases hormones to regulate glucose metabolism and balance glucose homeostasis when feeding. Our previous research indicated that high temperature significantly affects the levels of CCK, ghrelin, and GIP [
8]. In the current study, the results showed that 0.4 mg/kg Cr as supplemented Cr-Pic decreased the levels of CCK in the jejunum and circulation, increasing the GIP level in the jejunum. CCK is an anorexigenic hormone and, in broilers, an important regulator of appetite inhibition that is involved in the neurohumoral regulation of appetite [
39]. We previously mentioned that the feed intake of the 0.4 mg/kg Cr supplemented group significantly increased compared with that of the HT group, which means that the effect of appetite inhibition of CCK was weakened. GIP plays an important role in the regulation of blood glucose and insulin levels [
40]. In addition, GIP could regulate glucose uptake and lipoprotein lipase lipolysis, which plays a prominent role in glucose metabolism in adipose tissue [
41]. It was reported that GIP has been used as a novel hypoglycemic agent in diabetic patients [
42]. Interestingly, the concentration of GIP in the jejunum was significantly increased in the current study. The rate of glucose absorption determines the amount of GIP [
43]. In addition to playing a role in glucose transport, SGLT1 is also considered to be a glucose sensor in intestinal endocrine cells, promoting the secretion of GIP [
36,
44]. Hence, this may be due to up-regulated SGLT1 expression, because glucose-induced GIP secretion depends primarily on SGLT1 in the normal state [
45]. In addition, several forms of evidence suggest the microbiota could promote or inhibit the secretion of gut peptides such as GIP and PYY by SCFAs [
46,
47]. However, the exact mechanisms through which CCK regulates glucose levels and appetite are not known, and its regulation may be similar to other gut peptides such as GIP. So far, there have been no relevant reports on this, and further research is needed. Nevertheless, as research on the brain–gut axis is still very limited, this study can provide a reference for the pathway and mechanisms of the brain–gut axis.






{kind=link}
{kind=link}
{kind=link}