
Learning Ability and Hippocampal Transcriptome Responses to Early and Later Life Environmental Complexities in Dual-Purpose Chicks

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Data Collection
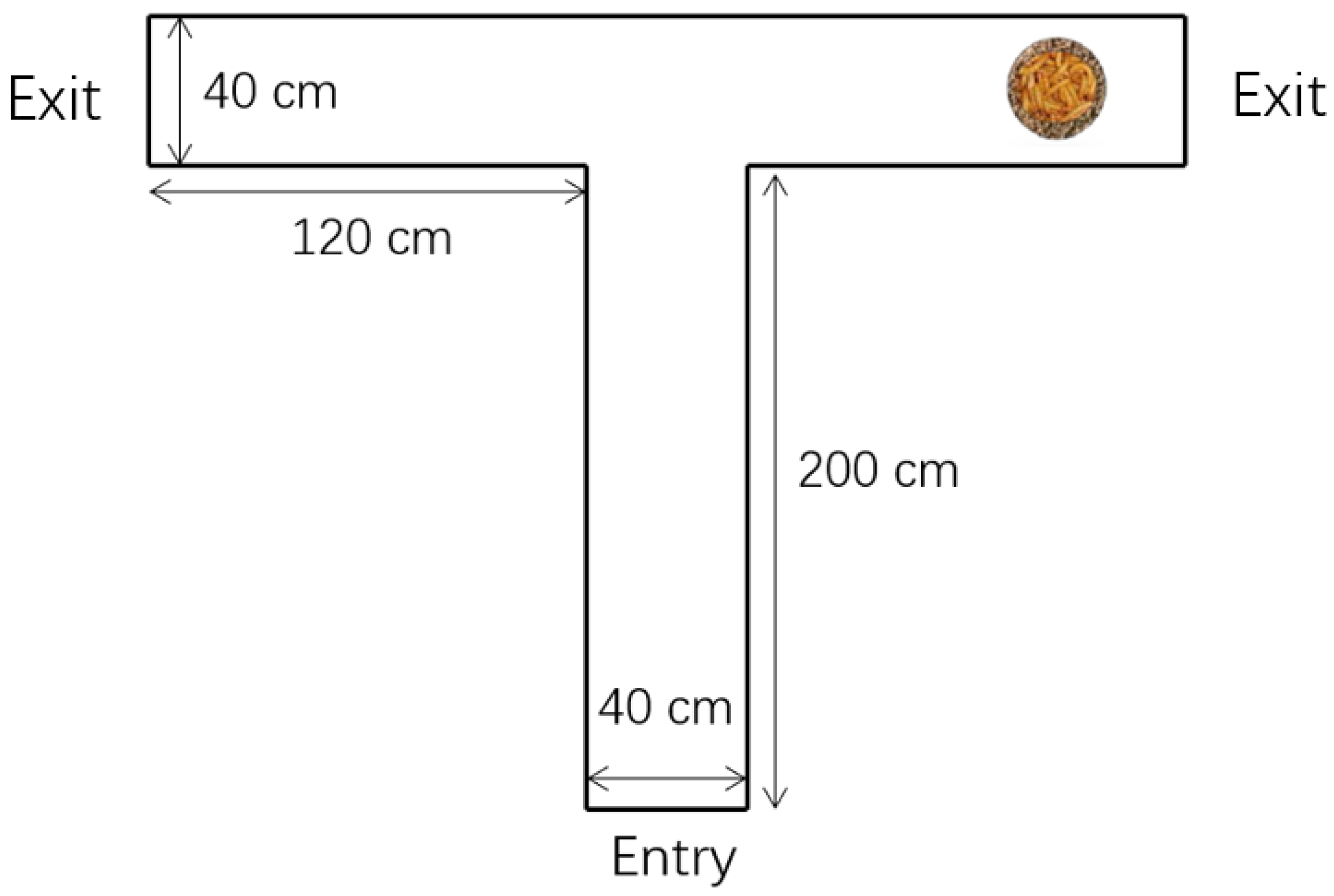
2.2.1. Learning Test
2.2.2. Creatine Kinase Concentration
2.2.3. Transcriptome Profiles of the Hippocampus
2.3. Statical Analysis
3. Results
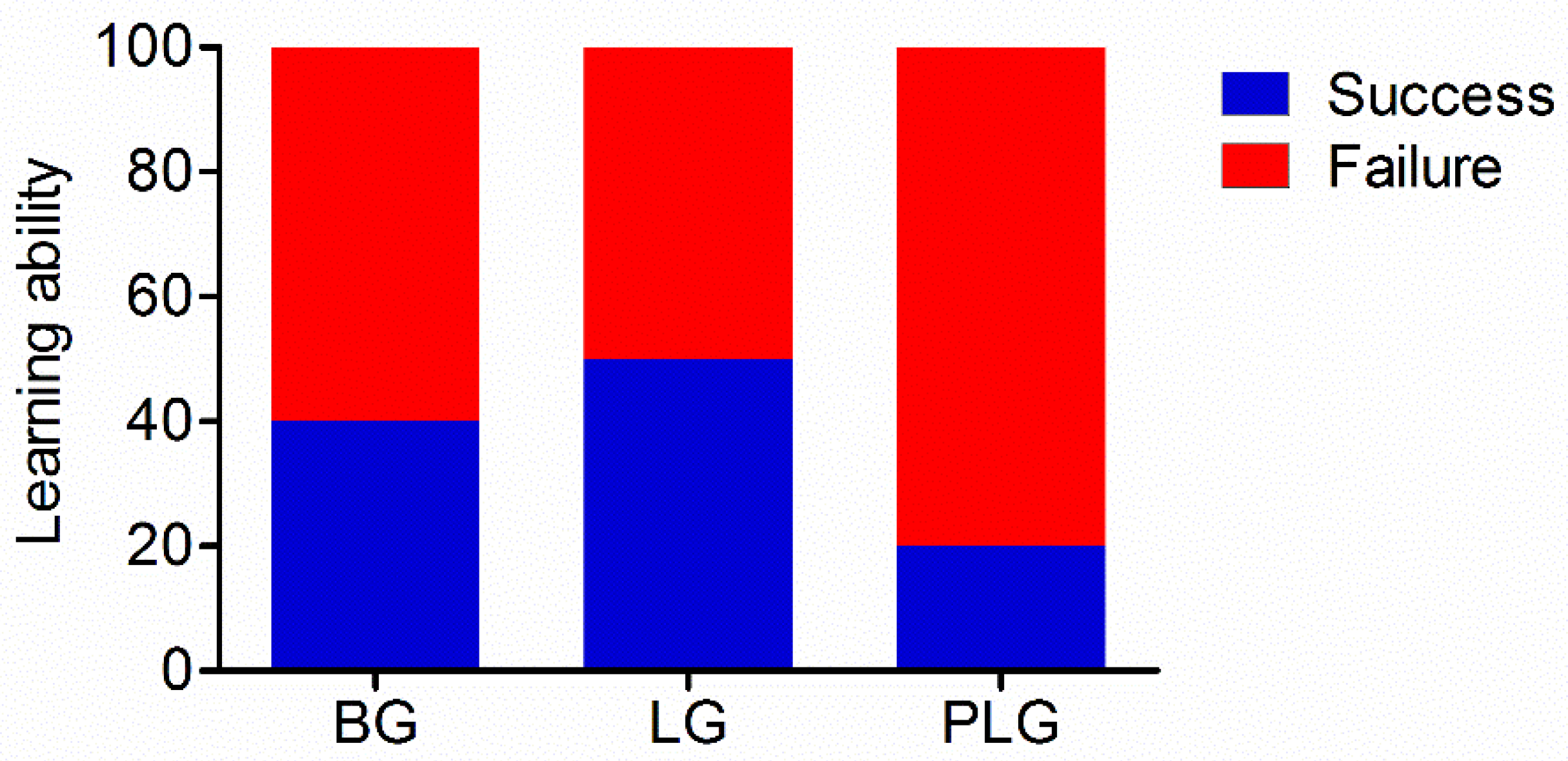
3.1. Learning Ability
3.2. Plasma Hormone Concentrations
3.3. Hippocampal Transcriptome
3.3.1. DEGs
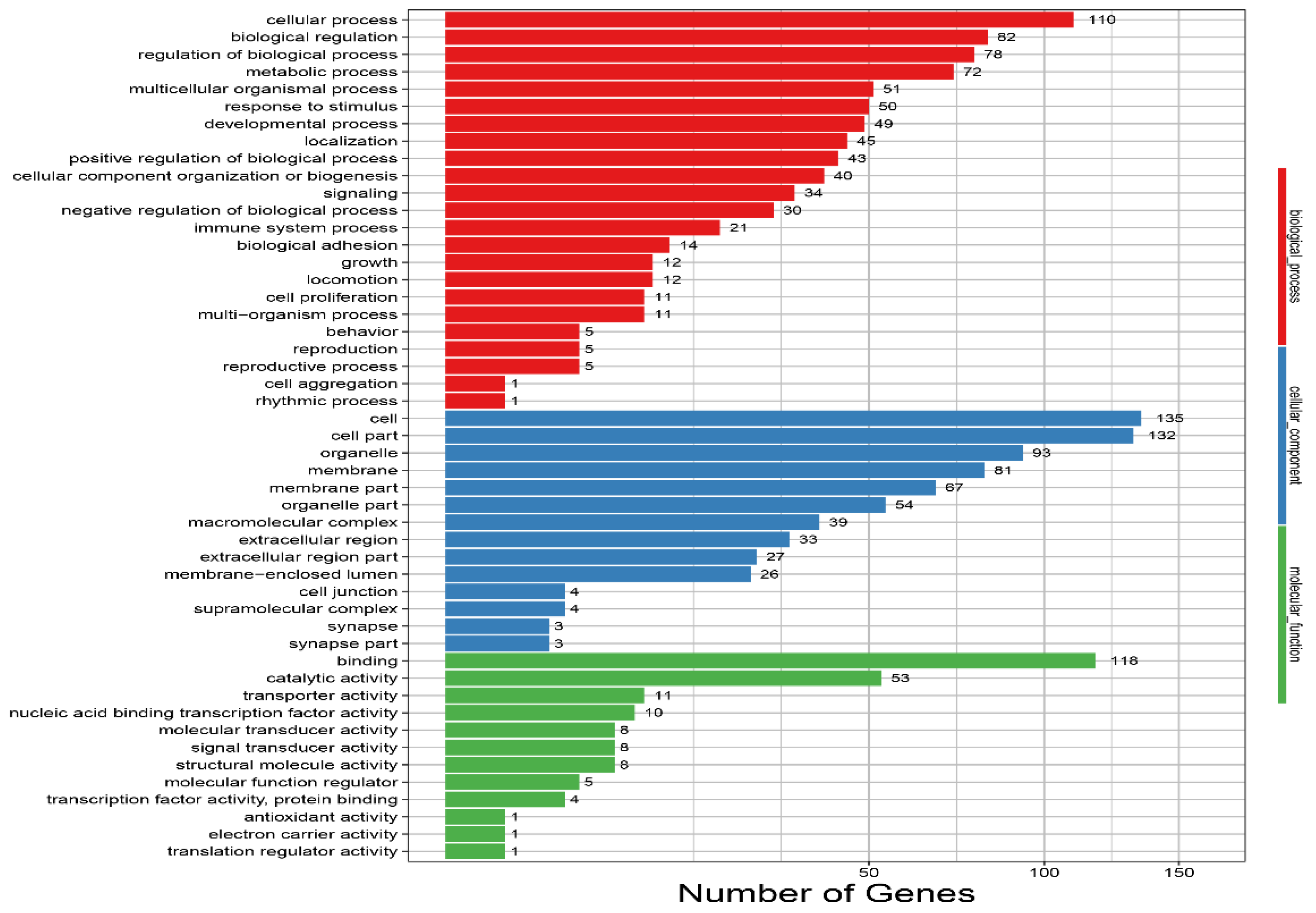
3.3.2. GO Terms
3.3.3. KEGG Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Székely, A.D. The avian hippocampal formation: Subdivisions and connectivity. Behav. Brain Res. 1999, 98, 219–225. [Google Scholar] [CrossRef]
- Sandi, C.; Rose, S.P.R.; Patterson, T.A. Unilateral hippocampal lesions prevent recall of a passive avoidance task in day-old chicks. Neurosci. Lett. 1992, 141, 255–258. [Google Scholar] [CrossRef]
- Pravosudov, V.V.; Kitaysky, A.S.; Omanska, A. The Relationship between Migratory Behaviour, Memory and the Hippocampus: An Intraspecific Comparison. Proc. Biol. Sci. 2006, 273, 2641–2649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nottebohm, B.F. Seasonal Recruitment of Hippocampal Neurons in Adult Free-Ranging Black-Capped Chickadees. Proc. Natl. Acad. Sci. USA 1994, 91, 11217–11221. [Google Scholar] [CrossRef] [Green Version]
- Clayton, N.S.; Krebs, J.R. Hippocampal growth and attrition in birds affected by experience. Proc. Natl. Acad. Sci. USA 1994, 96, 7410–7414. [Google Scholar] [CrossRef] [Green Version]
- Clayton, N.S. Development of memory and the hippocampus: Comparison of food-storing and nonstoring birds on a one-trial associative memory task. J. Neurosci. Off. J. Soc. Neurosci. 1995, 15, 2796. [Google Scholar] [CrossRef]
- Van Praag, H.; Kempermann, G.; Gage, F.H. Neural consequences of environmental enrichment. Nat. Rev. Neurosci. 2000, 1, 191–198. [Google Scholar] [CrossRef]
- Patzke, N.; Ocklenburg, S.; Van der Staay, F.J.; Güntürkün, O.; Manns, M. Consequences of different housing conditions on brain morphology in laying hens. J. Chem. Neuroanat. 2009, 37, 141–148. [Google Scholar] [CrossRef]
- Patrick, B.; Peter, G.; Mark, H. The biology of developmental plasticity and the Predictive Adaptive Response hypothesis. J. Physiol. 2014, 592, 2357–2368. [Google Scholar] [CrossRef]
- Pat, M. Early growth conditions, phenotypic development and environmental change. Philos. Trans. Biol. Sci. 2008, 363, 1635–1645. [Google Scholar] [CrossRef] [Green Version]
- Gluckman, P.D.; Hanson, M.A.; Beedle, A.S. Early life events and their consequences for later disease: A life history and evolutionary perspective. Am. J. Hum. Biol. 2010, 19, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Maniam, J.; Antoniadis, C.; Morris, M.J. Early-Life Stress, HPA Axis Adaptation, and Mechanisms Contributing to Later Health Outcomes. Front. Endocrinol. 2014, 5, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, G.; Hansman, C.; Heckman, J.J.; Novak, M.; Ruggiero, A.; Suomi, S.J. Primate evidence on the late health effects of early-life adversity. Proc. Natl. Acad. Sci. USA 2012, 109, 8866–8871. [Google Scholar] [CrossRef] [Green Version]
- Weaver, I.C.G.; Cervoni, N.; Champagne, F.A.; D’Alessio, A.C.; Sharma, S.; Seckl, J.R.; Dymov, S.; Szyf, M.; Meaney, M.J. Epigenetic programming by maternal behavior. Nat. Neurosci. 2004, 7, 847–854. [Google Scholar] [CrossRef]
- Oomen, C.A.; Soeters, H.; Audureau, N.; Vermunt, L.; Krugers, H. Severe early life stress hampers spatial learning and neurogenesis, but improves hippocampal synaptic plasticity and emotional learning under high-stress conditions in adulthood. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 6635–6645. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Reimert, I.; Middelkoop, A.; Kemp, B.; Bolhuis, J.E. Effects of Early and Current Environmental Enrichment on Behavior and Growth in Pigs. Front. Vet. Sci. 2020, 7, 268. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Reimert, I.; Haas, E.; Kemp, B.; Bolhuis, J.E. Effects of early and later life environmental enrichment and personality on attention bias in pigs (Sus scrofa domesticus). Anim. Cogn. 2019, 22, 959–972. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Jansen, C.A.; Bolhuis, J.E.; Arts, J.; Parmentier, H.K. Early and later life environmental enrichment affect specific antibody responses and blood leukocyte subpopulations in pigs. Physiol. Behav. 2020, 217, 112799. [Google Scholar] [CrossRef] [PubMed]
- Tahamtani, F.M.; Janicke, N.; Nordquist, R.E.; Janczak, A.M. Early Life in a Barren Environment Adversely Affects Spatial Cognition in Laying Hens (Gallus gallus domesticus). Front. Vet. Sci. 2015, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Cramer, T.; Rosenberg, T.; Kisliouk, T.; Meiri, N. Early-life epigenetic changes along the corticotropin-releasing hormone (CRH) gene influence resilience or vulnerability to heat stress later in life. Mol. Psychiatry 2019, 24, 1013–1026. [Google Scholar] [CrossRef]
- Christina, L.; Janczak, A.M.; Daniel, N.T.; Izabella, B.; Niclas, L.; Anette, W.; Joakim, L.; Johan, L.; Torjesen, P.A.; Per, J. Transmission of stress-induced learning impairment and associated brain gene expression from parents to offspring in chickens. PLoS ONE 2007, 2, e364. [Google Scholar]
- Lin, H.; Decuypere, E.; Buyse, J. Acute heat stress induces oxidative stress in broiler chickens. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2006, 144, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Debut, M.; Berri, C.; Arnould, C.; Guemené, D.; Santé-Lhoutellier, V.; Sellier, N.; Baéza, E.; Jehl, N.; Jégo, Y.; Beaumont, C. Behavioural and physiological responses of three chicken breeds to pre-slaughter shackling and acute heat stress. Br. Poult. Sci. 2005, 46, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Daehwan, K.; Ben, L.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Bo, L.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Reiner, A.; Yekutieli, D.; Benjamini, Y. Identifying differentially expressed genes using false discovery rate controlling procedures. Bioinformatics 2003, 19, 368–375. [Google Scholar] [CrossRef]
- Fu, J.J.; Geng, J.C.; Miao, Y.J.; Xu, Y.M.; Hu, T.M.; Yang, P.Z. Transcriptomic analyses reveal genotype- and organ-specific molecular responses to cold stress in Elymus nutans. Biol. Plant. 2018, 62, 671–683. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Khatri, P.; Sirota, M.; Butte, A.J. Ten years of pathway analysis: Current approaches and outstanding challenges. PLoS Comput. Biol. 2012, 8, e1002375. [Google Scholar] [CrossRef]
- Riber, A.B.; Ha, V.D.W.; de Jong, I.C.; Steenfeldt, S. Review of environmental enrichment for broiler chickens. Poult. Sci. 2018, 97, 378–396. [Google Scholar] [CrossRef] [PubMed]
- Tahamtani, F.M.; Pedersen, I.J.; Toinon, C.; Riber, A.B. Effects of environmental complexity on fearfulness and learning ability in fast growing broiler chickens. Appl. Anim. Behav. Sci. 2018, 207, 49–56. [Google Scholar] [CrossRef]
- Riber, A.B.; de Jong, I.C.; van de Weerd, H.A.; Steenfeldt, S. Environmental Enrichment for Broiler Breeders: An Undeveloped Field. Front. Vet. Sci. 2017, 4, 86. [Google Scholar] [CrossRef] [Green Version]
- Brantsæter, M.; Tahamtani, F.M.; Nordgreen, J.; Sandberg, E.; Hansen, T.B.; Rodenburg, T.B.; Moe, R.O.; Janczak, A.M. Access to litter during rearing and environmental enrichment during production reduce fearfulness in adult laying hens. Appl. Anim. Behav. Sci. 2017, 189, 49–56. [Google Scholar] [CrossRef]
- Newberry, R.C. Environmental enrichment—increasing the biological relevance of captive environments. Appl. Anim. Behav. Sci. 1995, 44, 229–243. [Google Scholar] [CrossRef]
- Tang, Y.P.; Wang, H.; Feng, R.; Kyin, M.; Tsien, J.Z. Differential effects of enrichment on learning and memory function in NR2B transgenic mice. Neuropharmacology 2001, 41, 779–790. [Google Scholar] [CrossRef]
- Sakimura, K.; Kutsuwada, T.; Ito, I.; Manabe, T.; Mishina, M. Reduced hippocampal LTP and spatial learning in mice lacking NMDA receptor I1 subunit. Nature 1995, 373, 151–155. [Google Scholar] [CrossRef]
- Bercik, P.; Denou, E.; Collins, J.; Jackson, W.; Lu, J.; Jury, J.; Deng, Y.; Blennerhassett, P.; Macri, J.; McCoy, K.D.; et al. The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology 2011, 141, 599–609.e3. [Google Scholar] [CrossRef] [Green Version]
- Montkowski, A.; Holsboer, F. Intact spatial learning and memory in transgenic mice with reduced BDNF. Neuroreport 1997, 8, 779–782. [Google Scholar] [CrossRef]
- Schrijver, N.C.; Pallier, P.N.; Brown, V.J.; Würbel, H. Double dissociation of social and environmental stimulation on spatial learning and reversal learning in rats. Behav. Brain Res. 2004, 152, 307–314. [Google Scholar] [CrossRef]
- Bolhuis, J.E.; Schouten, W.G.; de Leeuw, J.A.; Schrama, J.W.; Wiegant, V.M. Individual coping characteristics, rearing conditions and behavioural flexibility in pigs. Behav. Brain Res. 2004, 152, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Krause, E.T.; Naguib, M.; Trillmich, F.; Schrader, L. The effects of short term enrichment on learning in chickens from a laying strain (Gallus gallus domesticus). Appl. Anim. Behav. Sci. 2006, 101, 318–327. [Google Scholar] [CrossRef]
- Mitchell, M.A.; Sandercock, D.A. Creatine kinase isoenzyme profiles in the plasma of the domestic fowl (Gallus domesticus): Effects of acute heat stress. Res. Vet. Sci. 1995, 59, 30–34. [Google Scholar] [CrossRef]
- Yan, C.; Hartcher, K.; Liu, W.; Xiao, J.; Xiang, H.; Wang, J.; Liu, H.; Zhang, H.; Liu, J.; Chen, S.; et al. Adaptive response to a future life challenge: Consequences of early-life environmental complexity in dual-purpose chicks. J. Anim. Sci. 2020, 98, skaa348. [Google Scholar] [CrossRef]
- Singh, K.; Loreth, D.; Pöttker, B.; Hefti, K.; Innos, J.; Schwald, K.; Hengstler, H.; Menzel, L.; Sommer, C.J.; Radyushkin, K. Neuronal Growth and Behavioral Alterations in Mice Deficient for the Psychiatric Disease-Associated Negr1 Gene. Front. Mol. Neurosci. 2018, 11, 30. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Moreno, C.G.; Epardo, D.; Balderas-Marquez, J.E.; Fleming, T.; Carranza, M.; Luna, M.; Harvey, S.; Aramburo, C. Regenerative Effect of Growth Hormone (GH) in the Retina after Kainic Acid Excitotoxic Damage. Int. J. Mol. Sci. 2019, 20, 4433. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Sanroman, D.; Sanchis-Segura, C.; Toledo, R.; Hernandez, M.E.; Manzo, J.; Miquel, M. The effects of enriched environment on BDNF expression in the mouse cerebellum depending on the length of exposure. Behav. Brain Res. 2013, 243, 118–128. [Google Scholar] [CrossRef]
- Uma, R.; Li-Ann, C.; Bidesi, A.S.; Shad, M.U.; M Albert, T.; Hammen, C.L. Hippocampal changes associated with early-life adversity and vulnerability to depression. Biol. Psychiatry 2010, 67, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Kannaki, T.R.; Reddy, M.R.; Verma, P.C.; Shanmugam, M. Differential Toll-like receptor (TLR) mRNA expression patterns during chicken embryological development. Anim. Biotechnol. 2015, 26, 130–135. [Google Scholar] [CrossRef]
- Elwenspoek, M.M.C.; Kuehn, A.; Muller, C.P.; Turner, J.D. The effects of early life adversity on the immune system. Psychoneuroendocrinology 2017, 82, 140–154. [Google Scholar] [CrossRef]
- Simon, K. Effects of Early Life Conditions on Immunity in Broilers and Layers; Wageningen University: Wageningen, The Netherlands, 2016. [Google Scholar]
- Calabro, M.; Mandelli, L.; Crisafulli, C.; Lee, S.J.; Jun, T.Y.; Wang, S.M.; Patkar, A.A.; Masand, P.S.; Han, C.; Pae, C.U.; et al. Genes involved in neurodevelopment, neuroplasticity and major depression: No association for CACNA1C, CHRNA7 and MAPK1. Clin. Psychopharmacol. Neurosci. 2019, 17, 364–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegardt, F.G. Mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthase: A control enzyme in ketogenesis. Biochem. J. 1999, 338 Pt 3, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Laughlin, S.B. Energy as a constraint on the coding and processing of sensory information. Curr. Opin. Neurobiol. 2001, 11, 475–480. [Google Scholar] [CrossRef]
- Campbell, D.L.M.; De, H.E.N.; Lee, C. A review of environmental enrichment for laying hens during rearing in relation to their behavioral and physiological development. Poult. Sci. 2018, 98, 9–28. [Google Scholar] [CrossRef] [PubMed]
- Birkenkamp-Demtroder, K.; Wagner, L.; Brandt Sorensen, F.; Bording Astrup, L.; Gartner, W.; Scherubl, H.; Heine, B.; Christiansen, P.; Orntoft, T.F. Secretagogin is a novel marker for neuroendocrine differentiation. Neuroendocrinology 2005, 82, 121–138. [Google Scholar] [CrossRef] [PubMed]
- Zelent, D.; Najafi, H.; Odili, S.; Buettger, C.; Weik-Collins, H.; Li, C.; Doliba, N.; Grimsby, J.; Matschinsky, F.M. Glucokinase and glucose homeostasis: Proven concepts and new ideas. Biochem. Soc. Trans. 2005, 33, 306–310. [Google Scholar] [CrossRef] [Green Version]
- Fan, Q.-W.; Yu, W.; Gong, J.-S.; Zou, K.; Sawamura, N.; Senda, T.; Yanagisawa, K.; Michikawa, M. Cholesterol-dependent modulation of dendrite outgrowth and microtubule stability in cultured neurons. J. Neurochem. 2002, 80, 178–190. [Google Scholar] [CrossRef] [Green Version]
- Pfrieger, F.W.; Ungerer, N. Cholesterol metabolism in neurons and astrocytes. Prog. Lipid Res. 2011, 50, 357–371. [Google Scholar] [CrossRef]
- Parker, K.J.; Buckmaster, C.L.; Sundlass, K.; Schatzberg, A.F.; Lyons, D.M. Maternal mediation, stress inoculation, and the development of neuroendocrine stress resistance in primates. Proc. Natl. Acad. Sci. USA 2006, 103, 3000–3005. [Google Scholar] [CrossRef] [Green Version]
- Van Bodegom, M.; Homberg, J.R.; Henckens, M. Modulation of the hypothalamic-pituitary-adrenal axis by early life stress exposure. Front. Cell. Neurosci. 2017, 11, 87. [Google Scholar] [CrossRef] [Green Version]
- Turner, B.M. Epigenetic responses to environmental change and their evolutionary implications. Philos. Trans. R. Soc. B 2009, 364, 3403–3418. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BG—LG 1 | DEGs | |
|---|---|---|
| Upregulated | Downregulated | |
| Fatty acid biosynthesis | ACSL5 | |
| Primary bile acid biosynthesis | CH25H, BAA | |
| TNF signaling pathway | VCAM1, CASP18, MAP3K5, IL-18, IL18R1, and BGLE | TRAF5 |
| Cell adhesion molecules | VCAM1, TROJAN, ITGBL1, PTPRK, BGLECTIN, MHC-I, SIGLEC1, and ICOS | neuronal growth regulator 1 (NEGR1) and neurexins (NRXN1) |
| Apoptosis | CRCBL, CTSS, CASP18, and MAP3K5 | RAPGEF4, PTPN13, and KNDC1 |
| Regulation of actin cytoskeleton | ITGBL1, AKAIN1, SASH3, MYL9, LPXN, and APC | RASGEF1A, ARPC1A, ABI2, FGF12, ARHGEF4, DAZAP2, and KNDC1 |
| Malaria | TLR2, VCAM1, CR1L, BGLP, and BGLE | |
| Legionellosis | TLR2, CR1L, and CASP18 | |
| Pathways in cancer | ITGBL1, CASP18, AGTR1, SASH3, SPI1, ADCY8, GLI2, CDKN2A, GMSRAL, APC, CSF2RA, LAMA3/5, and NIPA1 | TRAF5, FGF12, RASGEF1A, RUNX1, and KNDC1 |
| BG—PLG 1 | DEGs | |
|---|---|---|
| Upregulated | Downregulated | |
| Proteasome | PSMB7 | |
| Amino sugar and nucleotide sugar metabolism | UXS1, glucokinase (GCK), GPI, and GFPT1 | |
| VEGF signaling pathway | AKT2, MAPKAPK3, and TGFB1I1 | |
| Regulation of lipolysis in adipocytes | MGLL | AKT2 |
| Malaria | TLR2A | |
| Nicotine addiction | Neuronal acetylcholine receptor subunit alpha-7-like (CHRNA8) | EHD2 and GRIA4 |
| LG—PLG 1 | DEGs | |
|---|---|---|
| Upregulated | Downregulated | |
| Synthesis and degradation of ketone bodies | HMGCS1 | |
| Butanoate metabolism | HMGCS1 | |
| Terpenoid backbone biosynthesis | HMGCS1 | |
| Homologous recombination | NBN | |
| Endocrine and other factor-regulated calcium reabsorption | Secretagogin (SCGN) | |
| Aminoacyl-tRNA biosynthesis | KARS | |
| Valine, leucine and isoleucine degradation | HMGCS1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, C.; Liu, W.; Xiao, J.; Xiang, H.; Wang, J.; Zhang, H.; Liu, J.; Chen, S.; Zhao, X. Learning Ability and Hippocampal Transcriptome Responses to Early and Later Life Environmental Complexities in Dual-Purpose Chicks. Animals 2022, 12, 668. https://doi.org/10.3390/ani12050668
Yan C, Liu W, Xiao J, Xiang H, Wang J, Zhang H, Liu J, Chen S, Zhao X. Learning Ability and Hippocampal Transcriptome Responses to Early and Later Life Environmental Complexities in Dual-Purpose Chicks. Animals. 2022; 12(5):668. https://doi.org/10.3390/ani12050668
Chicago/Turabian StyleYan, Chao, Wen Liu, Jinlong Xiao, Hai Xiang, Jikun Wang, Hui Zhang, Jian Liu, Siyu Chen, and Xingbo Zhao. 2022. "Learning Ability and Hippocampal Transcriptome Responses to Early and Later Life Environmental Complexities in Dual-Purpose Chicks" Animals 12, no. 5: 668. https://doi.org/10.3390/ani12050668