
Estimates of Effective Population Size in Commercial and Hatchery Strains of Coho Salmon (Oncorhynchus kisutch (Walbaum, 1792))


 ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Commercial and Hatchery Strains
2.2. Genotypic Data
2.3. Estimation of Linkage Disequilibrium and Effective Population Size (Ne)
3. Results
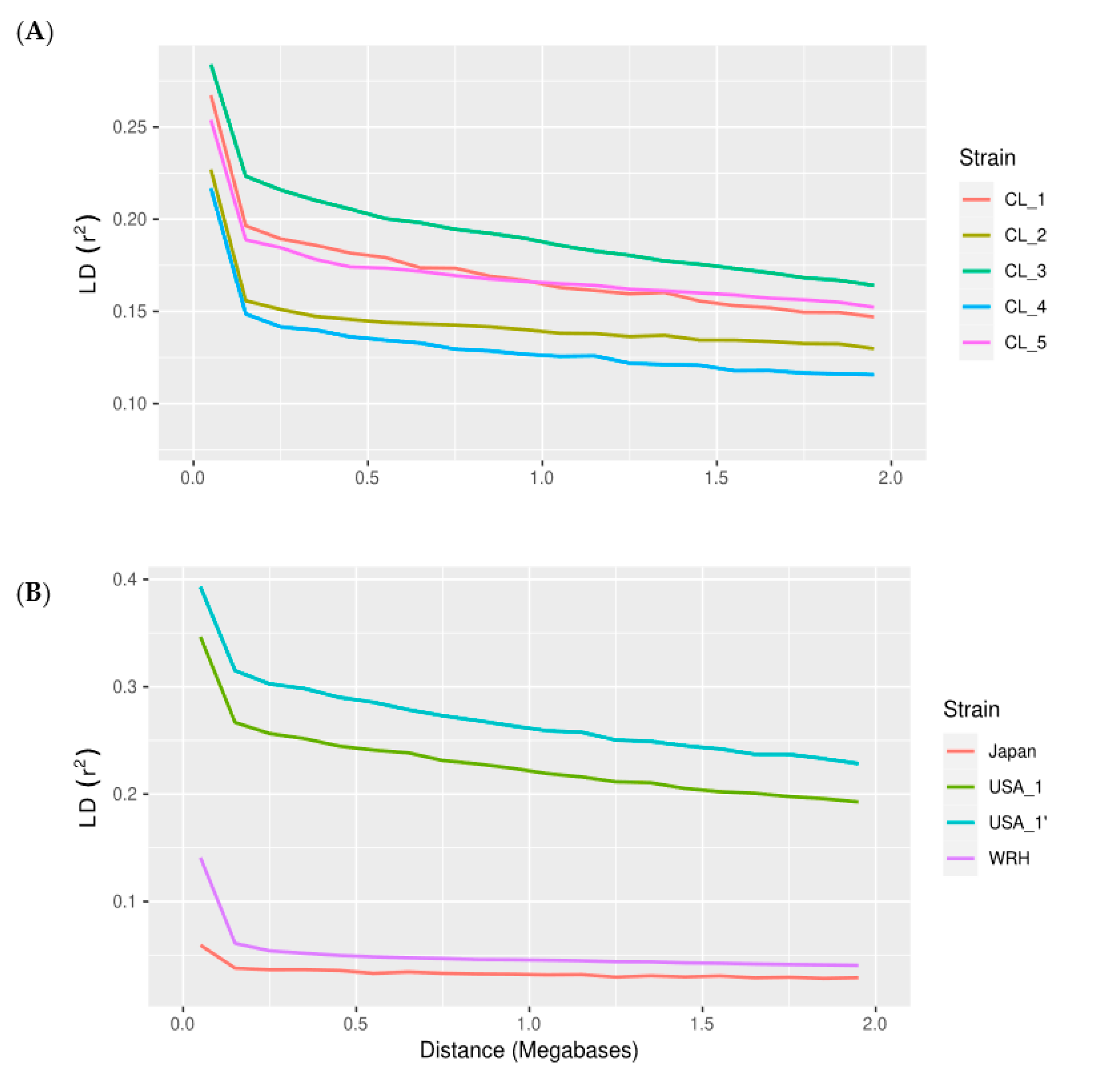
3.1. Linkage Disequilibrium (LD)
Marker Data
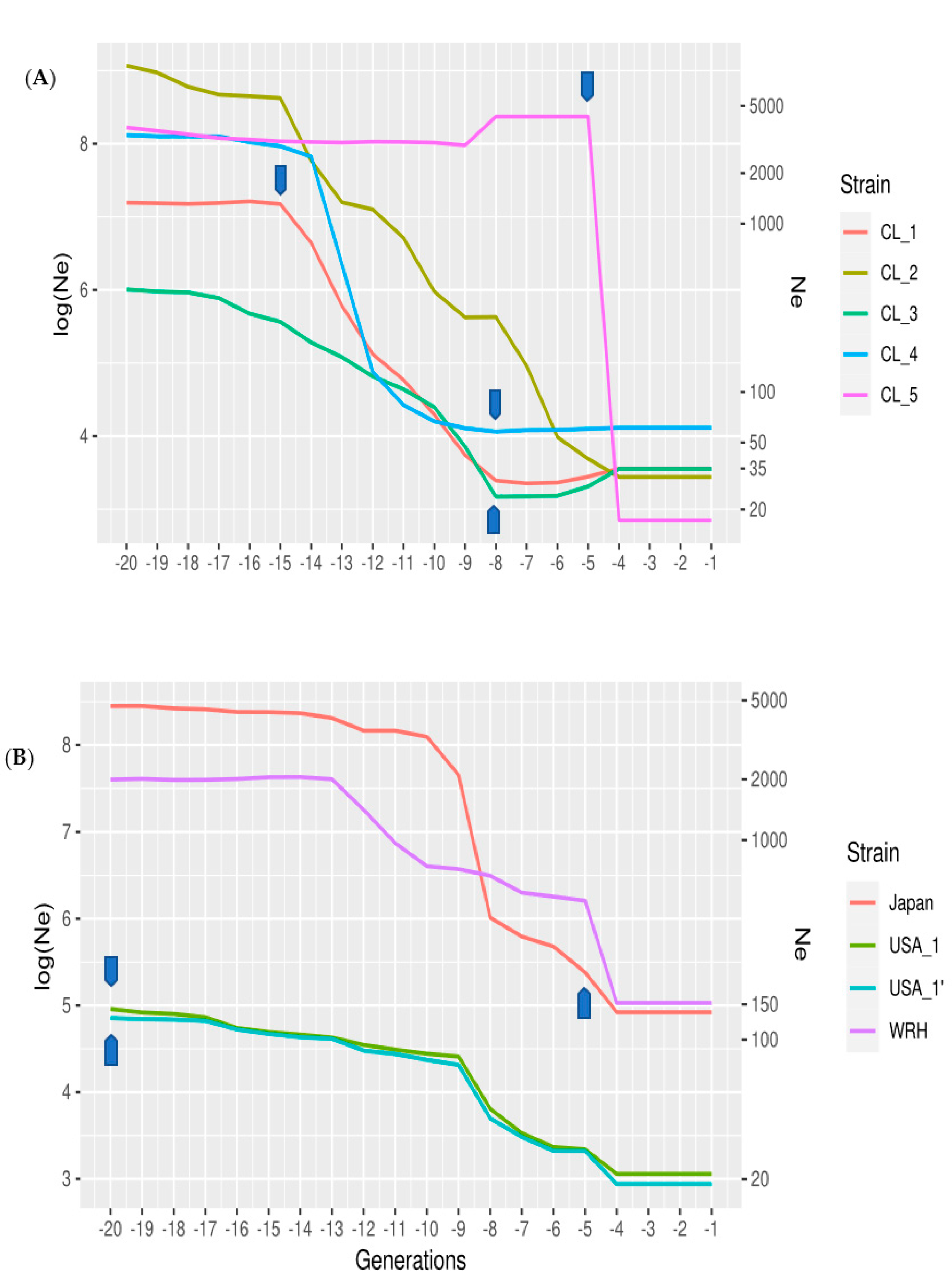
3.2. Patterns of Effective Population Size, Ne, across Generations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Winkler, F.M.; Bartley, D.; Díaz, N.F. Genetic differences among year classes in a hatchery population of coho salmon (Oncorhynchus kisutch (Walbaum, 1792)) in Chile. Aquaculture 1999, 173, 425–433. [Google Scholar] [CrossRef]
- Carrera, N.Í. Breve historia de la acuicultura y salmonicultura en el sur de Chile (1856–2000). Territ. Y Reg. 2020, 3, 36–49. [Google Scholar] [CrossRef]
- Sackton, T.B.; Clark, N. Convergent evolution in the genomics era: New insights and directions. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190102. [Google Scholar] [CrossRef] [PubMed]
- Eldridge, W.; Naish, K. Long-term effects of translocation and release numbers on fine-scale population structure among coho salmon (Oncorhynchus kisutch). Mol. Ecol. 2007, 16, 2407–2421. [Google Scholar] [CrossRef] [PubMed]
- Falconer, D.; Mackay, T. Introduction to Quantitative Genetics; Longman: Essex, UK, 1996. [Google Scholar]
- Martinez, V.; Kause, A.; Mäntysaari, E.; Mäki-Tanila, A. The use of alternative breeding schemes to enhance genetic improvement in rainbow trout (Oncorhynchus mykiss): I. One-stage selection. Aquaculture 2006, 254, 182–194. [Google Scholar] [CrossRef]
- Rozas, M.; Enriquez, R. Piscirickettsiosis and Piscirickettsia salmonis in fish: A review. J. Fish Dis. 2014, 37, 163–188. [Google Scholar] [CrossRef]
- Bravo, S. Piscirickettsiosis in freshwater. Bull. Eur. Assoc. Fish Pathol. 1994, 14, 137–138. [Google Scholar]
- Almendras, F.E.; Fuentealba, I.C.; Frederick Markham, R.F.; Speare, D.J. Pathogenesis of liver lesions caused by experimental infection with Piscirickettsia salmonis in juvenile Atlantic salmon, Salmo salar L. J. Vet. Diagn. Invest. 2000, 12, 552–557. [Google Scholar] [CrossRef]
- Martinez, V.; Naish, K.; Bravo, C.; Cichero, D.; Lien, S.; Nome, T.; Iwamoto, R.; Moen, T. Development of a high-density genomic platform for Coho salmon and its use in population assessment in Chile and North America, Proceedings of the World Congress on Genetics Applied to Livestock Production Electronic Poster Session—Species. Aquaculture 2018, 2, 968. [Google Scholar]
- Myers, J.; Hershberger, W.; Saxton, A.; Iwamoto, R. Estimates of genetic and phenotypic parameters for length and weight of marine net-pen reared coho salmon (Oncorhynchus kisutch Walbaum). Aquac. Res. 2002, 32, 277–285. [Google Scholar] [CrossRef]
- Withler, R.E.; Evelyn, T.P.T. Genetic Variation in Resistance to Bacterial Kidney Disease within and between Two Strains of Coho Salmon from British Columbia. Trans. Am. Fish. Soc. 1990, 119, 1003–1009. [Google Scholar] [CrossRef]
- Hosoya, S.; Kikuchi, K.; Nagashima, H.; Onodera, J.; Sugimoto, K.; Satoh, K.; Matsuzaki, K.; Yasugi, M.; Nagano, A.J.; Kumagayi, A.; et al. Assessment of genetic diversity in Coho salmon (Oncorhynchus kisutch) populations with no family records using ddRAD-seq. BMC Res. Notes 2018, 11, 548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neely, K.; Myers, J.; Hard, J. A Comparison of Early Development between a Domesticated Stock of Coho Salmon and Its Parental Stock. Trans. Am. Fish. Soc. 2012, 141, 1504–1509. [Google Scholar] [CrossRef]
- Hershberger, W.K.; Myers, J.M.; Iwamoto, R.N.; McAuley, W.C.; Saxton, A.M. Genetic changes in the growth of coho salmon (Oncorhynchus kisutch) in marine net-pens, produced by ten years of selection. Aquaculture 1990, 85, 187–197. [Google Scholar] [CrossRef]
- Iwamoto, R.N.; Towner, R.; Myers, J.; Hudson, G.; Munsell, P. Coho salmon broodstock development: A case study of the Domsea Coho salmon (1977 to 2015). Bull. Jpn. Fish. Res. Educ. Agency 2017, 45, 107–112. [Google Scholar]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Tarasov, A.; Vilella, A.J.; Cuppen, E.; Nijman, I.J.; Prins, P. Sambamba: Fast processing of NGS alignment formats. Bioinformatics 2015, 31, 2032–2034. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. 1000 Genomes Project Analysis group, The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Browning, B.L.; Zhou, Y.; Browning, S.R. A One-Penny Imputed Genome from Next-Generation Reference Panels. Am. J. Hum. Genet. 2018, 103, 338–348. [Google Scholar] [CrossRef] [Green Version]
- Hill, W.G.; Robertson, A. Linkage disequilibrium in finite populations. Theor. Appl. Genet. 1968, 38, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4, s13742-015. [Google Scholar] [CrossRef] [PubMed]
- Holte, R.C. Very Simple Classification Rules Perform Well on Most Commonly Used Datasets. Mach. Learn. 1993, 11, 63–90. [Google Scholar] [CrossRef]
- Santiago, E.; Novo, I.; Pardiñas, A.F.; Saura, M.; Wang, J.; Caballero, A. Recent Demographic History Inferred by High-Resolution Analysis of Linkage Disequilibrium. Mol. Biol. Evol. 2020, 37, 3642–3653. [Google Scholar] [CrossRef] [PubMed]
- Saura, M.; Caballero, A.; Santiago, E.; Fernández, A.; Morales-González, E.; Fernández, J.; Cabaleiro, S.; Millán, A.; Martínez, P.; Palaiokostas, B.; et al. Estimates of recent and historical effective population size in turbot, seabream, seabass and carp selective breeding programmes. Genet. Sel. Evol. 2021, 53, 85. [Google Scholar] [CrossRef] [PubMed]
- Corbin, L.J.; Liu, A.Y.; Bishop, S.C.; Woolliams, J.A. Estimation of historical effective population size using linkage disequilibria with marker data. J. Anim. Breed Genet. 2012, 129, 257–270. [Google Scholar] [CrossRef]
- Hayes, B.J.; Visscher, P.M.; McPartlan, H.C.; Goddard, M.E. Novel multilocus measure of linkage disequilibrium to estimate past effective population size. Genome Res. 2003, 13, 635–643. [Google Scholar] [CrossRef] [Green Version]
- Barbato, M.; Orozco-terWengel, P.; Tapio, M.; Bruford, M.W. SNeP: A tool to estimate trends in recent effective population size trajectories using genome-wide SNP data. Front. Genet. 2015, 6, 109. [Google Scholar] [CrossRef] [Green Version]
- Dettleff, P.; Hernandez, E.; Partridge, G.; Lafarga-De la Cruz, F.; Martinez, V. Understanding the population structure and reproductive behavior of hatchery-produced yellowtail kingfish (Seriola lalandi). Aquaculture 2020, 522, 734948. [Google Scholar] [CrossRef]
- D’Ambrosio, J.; Phocas, F.; Haffray, P.; Bestin, A.; Brard-Fudulea, S.; Poncet, C.; Quillet, E.; Dechamp, N.; Fraslin, C.; Charles, M.; et al. Genome-wide estimates of genetic diversity, inbreeding and effective size of experimental and commercial rainbow trout lines undergoing selective breeding. Genet. Sel. Evol. 2019, 51, 26. [Google Scholar] [CrossRef] [Green Version]
- Frankham, R.; Ballou, S.E.J.D.; Briscoe, D.A.; Ballou, J.D. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Martínez, V.C.; Neira, R.; Gall, G.A.E. Estimation of genetic parameters from pedigreed populations: Lessons from analysis of alevin weight in Coho salmon (Oncorhynchus kisutch). Aquaculture 1999, 180, 223–236. [Google Scholar] [CrossRef]
- Di Prinzio, C.Y.; Penaluna, B.; Grech, M.G.; Manzo, L.M.; Miserendino, M.L.; Casaux, R. Impact of Chaitén Volcano ashfall on native and exotic fish recovery, recolonization, and abundance. Sci. Total Environ. 2021, 752, 141864. [Google Scholar] [CrossRef] [PubMed]
- Ryman, N.; Laikre, L. Effects of supportive breeding on the genetically effective population size. Conserv. Biol. 1991, 5, 325–329. [Google Scholar] [CrossRef]
- Thériault, V.; Moyer, G.R.; Jackson, L.S.; Blouin, M.S.; Banks, M.A. Reduced reproductive success of hatchery coho salmon in the wild: Insights into most likely mechanisms. Mol. Ecol. 2011, 20, 1860–1869. [Google Scholar] [CrossRef]
- Blouin, M.S.; Wrey, M.C.; Bollmann, S.R.; Skaar, J.C.; Twibell, R.G.; Fuentes, C. Offspring of first-generation hatchery steelhead trout (Oncorhynchus mykiss) grow faster in the hatchery than offspring of wild fish, but survive worse in the wild: Possible mechanisms for inadvertent domestication and fitness loss in hatchery salmon. PLoS ONE 2021, 16, e0257407. [Google Scholar] [CrossRef]
- Wang, L.; Sørensen, P.; Janss, L.; Ostersen, T.; Edwards, D. Genome-wide and local pattern of linkage disequilibrium and persistence of phase for 3 Danish pig breeds. BMC Genet. 2013, 14, 115. [Google Scholar] [CrossRef] [Green Version]
- Klerks, P.L.; Athrey, G.N.; Leberg, P.L. Response to Selection for Increased Heat Tolerance in a Small Fish Species, With the Response Decreased by a Population Bottleneck. Front. Ecol. Evol. 2019, 7, 270. [Google Scholar] [CrossRef] [Green Version]
- Razgour, O.; Forester, B.; Taggart, J.B.; Bekaert, M.; Juste, J.; Ibáñez, C.; Puechmaille, S.J.; Novella-Fernandez, R.; Alberdi, A.; Manel, S. Considering adaptive genetic variation in climate change vulnerability assessment reduces species range loss projections. Proc. Natl. Acad. Sci. USA 2019, 116, 10418. [Google Scholar] [CrossRef] [Green Version]
- Debes, P.V.; Solberg, M.F.; Matre, I.H.; Dyrhovden, L.; Glover, K.A. Genetic variation for upper thermal tolerance diminishes within and between populations with increasing acclimation temperature in Atlantic salmon. Heredity 2021, 127, 455–466. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Population | Abbreviation | Number of samples | Number of SNPs Genotyped | Median MAF |
|---|---|---|---|---|
| Chile Strain 1 | CL_1 | 45 | 146,945 | 0.19 |
| Chile Strain 2 | CL_2 | 20 | 135,663 | 0.23 |
| Chile Strain 3 | CL_3 | 44 | 138,893 | 0.31 |
| Chile Strain 4 | CL_4 | 40 | 135,868 | 0.21 |
| Chile Strain 5 | CL_5 | 36 | 135,743 | 0.28 |
| Domsea Even | USA_1 | 30 | 135,624 | 0.17 |
| Domesea Odd | USA_1′ | 25 | 136,297 | 0.18 |
| Wallace River | WRH | 46 | 146,288 | 0.24 |
| Japan | JAPAN | 112 | 51,794 | 0.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez, V.; Dettleff, P.J.; Galarce, N.; Bravo, C.; Dorner, J.; Iwamoto, R.N.; Naish, K. Estimates of Effective Population Size in Commercial and Hatchery Strains of Coho Salmon (Oncorhynchus kisutch (Walbaum, 1792)). Animals 2022, 12, 647. https://doi.org/10.3390/ani12050647
Martinez V, Dettleff PJ, Galarce N, Bravo C, Dorner J, Iwamoto RN, Naish K. Estimates of Effective Population Size in Commercial and Hatchery Strains of Coho Salmon (Oncorhynchus kisutch (Walbaum, 1792)). Animals. 2022; 12(5):647. https://doi.org/10.3390/ani12050647
Chicago/Turabian StyleMartinez, Victor, Phillip J. Dettleff, Nicolás Galarce, Cristian Bravo, Jessica Dorner, Robert N. Iwamoto, and Kerry Naish. 2022. "Estimates of Effective Population Size in Commercial and Hatchery Strains of Coho Salmon (Oncorhynchus kisutch (Walbaum, 1792))" Animals 12, no. 5: 647. https://doi.org/10.3390/ani12050647