
Mismatches between Morphology and DNA in Italian Partridges May Not Be Explained Only by Recent Artificial Release of Farm-Reared Birds

 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples, Genetic Marker and Procedures
2.1.1. Displacement Loop (DLOOP)
2.1.2. Cytochrome Oxidase Subunit I (COI)
2.1.3. Oocyte Maturation Factor Mos (CMOS)
2.1.4. Additional Procedures
2.2. Phylogenetic Analyses
3. Results
3.1. The Dataset
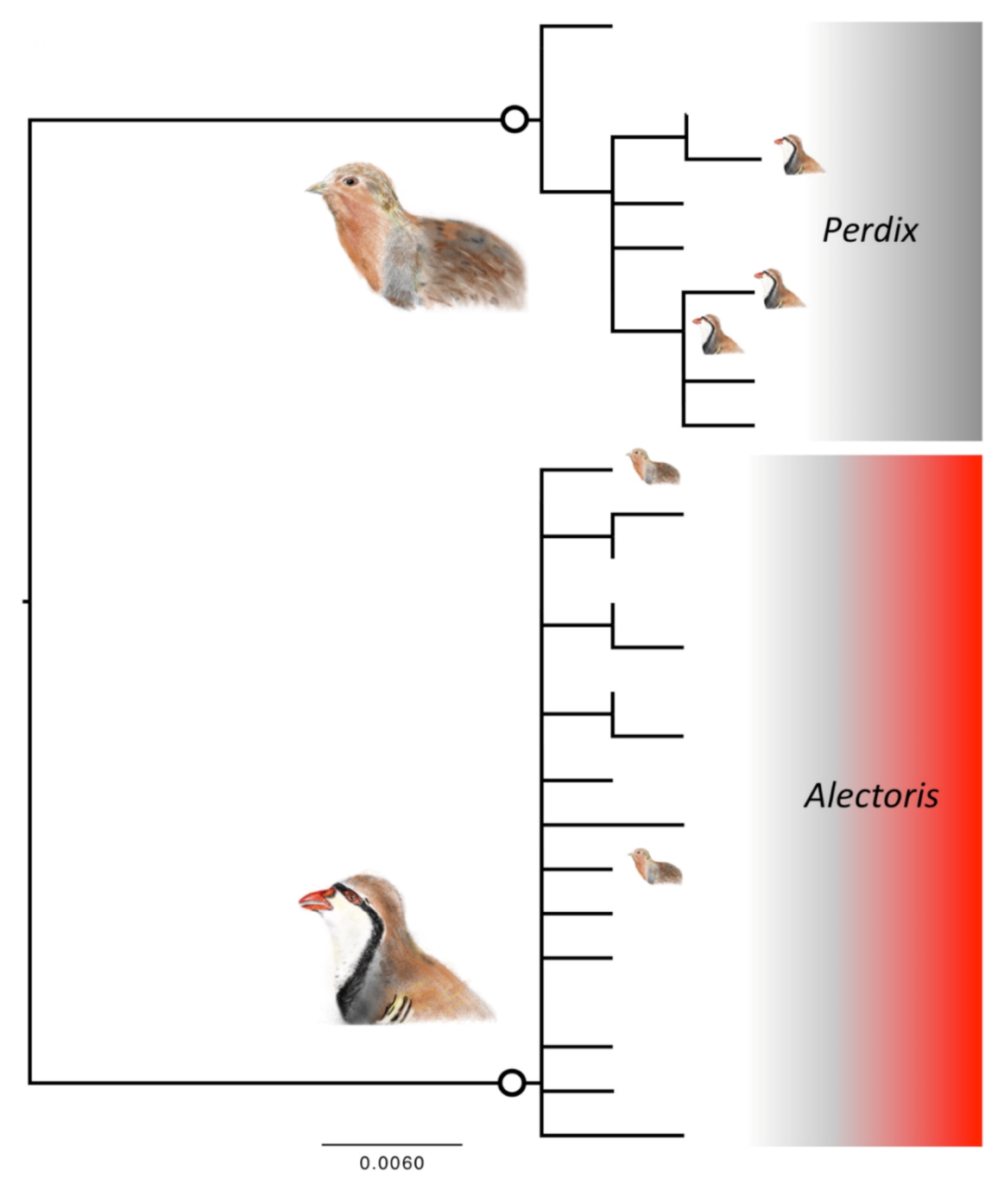
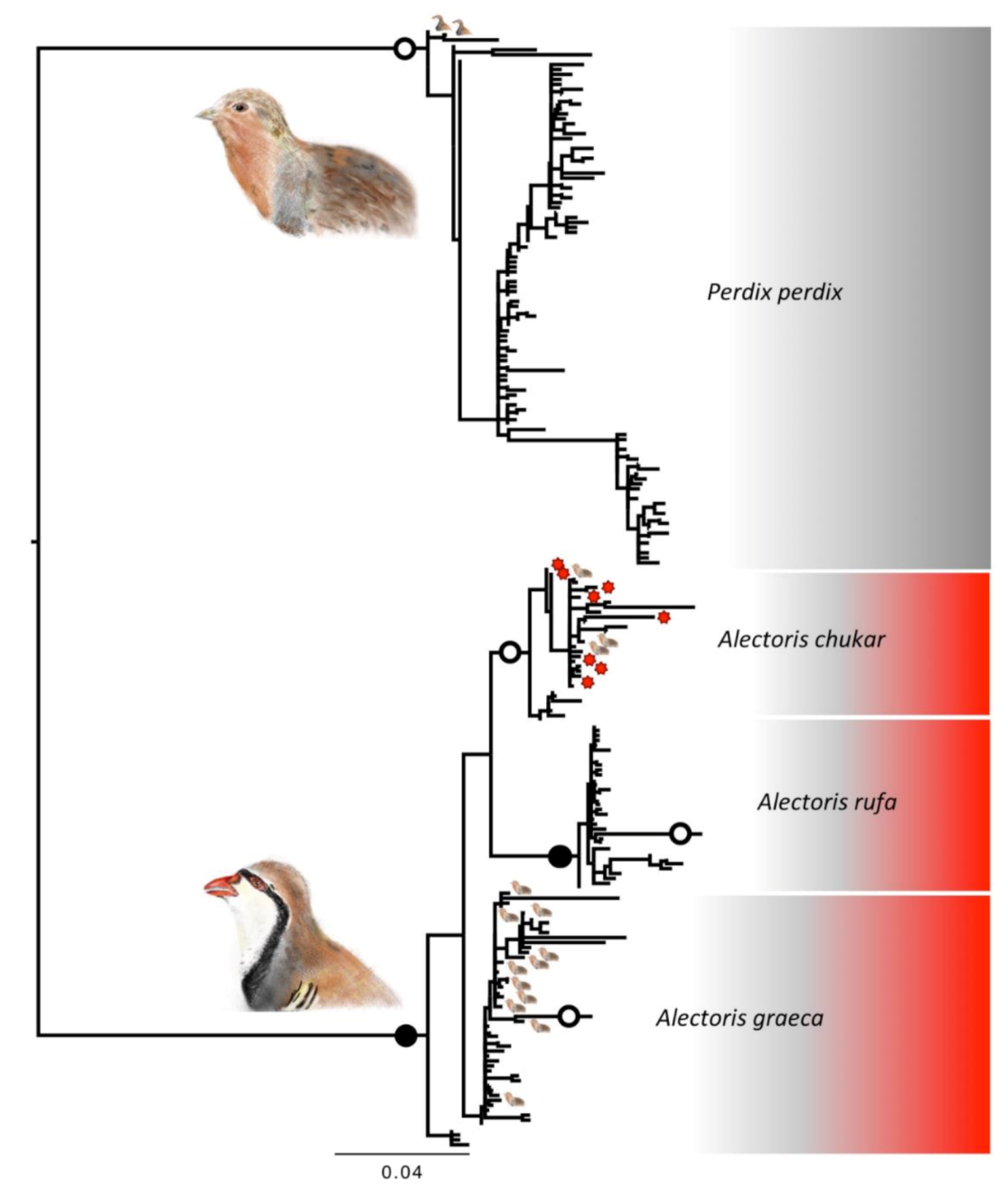
3.2. Phylogenetic Reconstructions
3.3. Spread of Mismatch between Morphological Classification and DNA Taxonomy
3.4. Temporal Progression of Mismatch between Morphological Classification and DNA Taxonomy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cramp, S.; Simmons, K.E.L. Handbook of the Birds of Europe, the Middle East, and North Africa: The Birds of the Western Palearctic—Volume II—Hawks to Bustards; Oxford University Press: Oxford, UK, 1980. [Google Scholar]
- Wang, N.; Kimball, R.T.; Braun, E.L.; Liang, B.; Zhang, Z. Assessing phylogenetic relationships among Galliformes: A multigene phylogeny with expanded taxon sampling in Phasianidae. PLoS ONE 2013, 8, e64312. [Google Scholar] [CrossRef]
- Van Tuinen, M.; Dyke, G.J. Calibration of galliform molecular clocks using multiple fossils and genetic partitions. Mol. Phylogenet. Evol. 2004, 30, 74–86. [Google Scholar] [CrossRef]
- Shen, Y.-Y.; Liang, L.; Sun, Y.-B.; Yue, B.-S.; Yang, X.-J.; Murphy, R.W.; Zhang, Y.-P. A mitogenomic perspective on the ancient, rapid radiation in the Galliformes with an emphasis on the Phasianidae. BMC Evol. Biol. 2010, 10, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, M.H.; Braun, E.L.; Kimball, R.T. Phylogenetic utility of avian Ovomucoid Intron G: A comparison of nuclear and mitochondrial phylogenies in Galliformes. Auk 2001, 118, 799–804. [Google Scholar] [CrossRef]
- Dyke, G.J.; Gulas, B.E.; Crowe, T.M. Suprageneric relationships of galliform birds (Aves, Galliformes): A cladistic analysis of morphological characters. Zool. J. Linn. Soc. 2003, 137, 227–244. [Google Scholar] [CrossRef] [Green Version]
- Kimball, R.T.; Braun, E.L. Does more sequence data improve estimates of galliform phylogeny? Analyses of a rapid radiation using a complete data matrix. PeerJ 2014, 2, e361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnsgard, P.A. The Quails, Partridges, and Francolins of the World; Oxford University Press: New York, NY, USA, 1988. [Google Scholar]
- Kimball, R.T.; Braun, E.L.; Zwartjes, P.W.; Crowe, T.M.; Ligon, J.D. A molecular phylogeny of the pheasants and partridges suggests that these lineages are not monophyletic. Mol. Phylogenet. Evol. 1999, 11, 38–54. [Google Scholar] [CrossRef] [PubMed]
- Forcina, G.; Guerrini, M.; Khaliq, I.; Khan, A.A.; Barbanera, F. Human-modified biogeographic patterns and conservation in game birds: The dilemma of the black francolin (Francolinus francolinus, Phasianidae) in Pakistan. PLoS ONE 2018, 13, e0205059. [Google Scholar] [CrossRef] [Green Version]
- Forcina, G.; Guerrini, M.; Barbanera, F. Non-native and hybrid in a changing environment: Conservation perspectives for the last Italian red-legged partridge (Alectoris rufa) population with long natural history. Zoology 2020, 138, 125740. [Google Scholar] [CrossRef]
- Bernard-Laurent, A.; De Franceschi, P.F. Statut, évolution et facteurs limitant les populations de perdrix bartavelle (Alectoris graeca): Synthèse bibliographique. Gibier Faune Sauvag. 1994, 11, 267–307. [Google Scholar]
- Bro, E.; Reitz, F.; Landry, P. Grey partridge Perdix perdix population status in central northern France: Spatial variability in density and 1994-2004 trend. Wildl. Biol. 2005, 11, 287–298. [Google Scholar] [CrossRef] [Green Version]
- Panek, M. Monitoring grey partridge (Perdix perdix) populations in Poland: Methods and results. Wildl. Biol. Pract. 2006, 2, 72–78. [Google Scholar] [CrossRef]
- Barilani, M.; Sfougaris, A.; Giannakopoulos, A.; Mucci, N.; Tabarroni, C.; Randi, E. Detecting introgressive hybridisation in rock partridge populations (Alectoris graeca) in Greece through Bayesian admixture analyses of multilocus genotypes. Conserv. Genet. 2007, 8, 343–354. [Google Scholar] [CrossRef]
- Barilani, M.; Bernard-Laurent, A.; Mucci, N.; Tabarroni, C.; Kark, S.; Perez Garrido, J.A.; Randi, E. Hybridisation with introduced chukars (Alectoris chukar) threatens the gene pool integrity of native rock (A. graeca) and red-legged (A. rufa) partridge populations. Biol. Conserv. 2007, 137, 57–69. [Google Scholar] [CrossRef]
- Meriggi, A.; della Stella, R.M.; Brangi, A.; Ferloni, M.; Masseroni, E.; Merli, E.; Pompilio, L. The reintroduction of grey and red-legged partridges (Perdix perdix and Alectoris rufa) in central Italy: A metapopulation approach. Ital. J. Zool. 2007, 74, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Andersen, L.W.; Kahlert, J. Genetic indications of translocated and stocked grey partridges (Perdix perdix): Does the indigenous Danish grey partridge still exist? Biol. J. Linn. Soc. 2012, 105, 694–710. [Google Scholar] [CrossRef]
- Barbanera, F.; Negro, J.J.; Di Giuseppe, G.; Bertoncini, F.; Cappelli, F.; Dini, F. Analysis of the genetic structure of red-legged partridge (Alectoris rufa, Galliformes) populations by means of mitochondrial DNA and RAPD markers: A study from central Italy. Biol. Conserv. 2005, 122, 275–287. [Google Scholar] [CrossRef]
- Barbanera, F.; Guerrini, M.; Khan, A.A.; Panayides, P.; Hadjigerou, P.; Sokos, C.; Gombobaatar, S.; Samadi, S.; Khan, B.Y.; Tofanelli, S.; et al. Human-mediated introgression of exotic chukar (Alectoris chukar, Galliformes) genes from East Asia into native Mediterranean partridges. Biol. Invasions 2009, 11, 333–348. [Google Scholar] [CrossRef]
- Gandolfi, A.; Crestanello, B.; Fagotti, A.; Simoncelli, F.; Chiesa, S.; Girardi, M.; Giovagnoli, E.; Marangoni, C.; Di Rosa, I.; Lucentini, L. New evidences of mitochondrial dna heteroplasmy by putative paternal leakage between the rock partridge (Alectoris graeca) and the chukar partridge (Alectoris chukar). PLoS ONE 2017, 12, e0170507. [Google Scholar] [CrossRef]
- Randi, E.; Bernard-Laurent, A. Population genetics of a hybrid zone between the red-legged partridge and rock partridge. Auk 1999, 116, 324–337. [Google Scholar] [CrossRef]
- Priolo, A. Variabilità in Alectoris graeca e descrizione di A graeca orlandoi subsp. nova degli Appennini. Riv. Ital. Ornitol. 1984, 54, 45–76. [Google Scholar]
- Randi, E.; Tabarroni, C.; Rimondi, S.; Lucchini, V.; Sfougaris, A. Phylogeography of the rock partridge (Alectoris graeca). Mol. Ecol. 2003, 12, 2201–2214. [Google Scholar] [CrossRef] [PubMed]
- Bernard-Laurent, A. Rock partridge. In The EBCC Atlas of European Breeding Birds, Their Distribution and Abundance; Hagemeijer, W.J.M., Blair, M.J., Eds.; T. & A.D. Poyser Ltd.: London, UK, 1997; p. 207. [Google Scholar]
- Peronace, V.; Cecere, J.G.; Gustin, M.; Rondinini, C. Lista rossa 2011 degli uccelli nidificanti in Italia. Avocetta 2012, 36, 11–58. [Google Scholar]
- Rondinini, C.; Battistoni, A.; Peronace, V.; Teofili, C. Lista Rossa dei Vertebrati Italiani; Comitato Italiano IUCN and MATTM: Rome, Italy, 2013. [Google Scholar]
- BirdLife International Alectoris graeca. The IUCN Red List of Threatened Species. Available online: https://doi.org/10.2305/IUCN.UK.2020-3.RLTS.T22678684A183062253.en (accessed on 12 May 2021).
- BirdLife International Species Factsheet: Alectoris graeca. Available online: http://datazone.birdlife.org/species/factsheet/rock-partridge-alectoris-graeca (accessed on 13 October 2021).
- BirdLife International. European Birds of Conservation Concern Populations, Trends and National Responsibilities; BirdLife International: Cambridge, UK, 2017; ISBN 978-1-912086-00-9. [Google Scholar]
- Budinski, I.; Čulina, A.; Mikulić, K.; Jurinović, L. Bird species that have significantly changed breeding range on Croatian coastal area: Comparison of 30 years old data and recent knowledge. Bird Census News 2010, 23, 49–58. [Google Scholar]
- Rippa, D.; Maselli, V.; Soppelsa, O.; Fulgione, D. The impact of agro-pastoral abandonment on the Rock Partridge Alectoris graeca in the Apennines. Ibis 2011, 153, 721–734. [Google Scholar] [CrossRef]
- Knaus, P.; Antoniazza, S.; Wechsler, S.; Guélat, J.; Kéry, M.; Strebel, N.; Sattler, T. Swiss Breeding Bird Atlas 2013–2016. Distribution and Population Trends of Birds in Switzerland and Liechtenstein; Swiss Ornithological Institute: Sempach, Switzerland, 2018; ISBN 9783859490130. [Google Scholar]
- Brusaferro, A.; Iesari, V.; Tardella, F.M.; Scocco, P.; Catorci, A. Effect of grassland mosaic structure and dynamism on the reproductive habitat suitability for Alectoris graeca in central Apennines. J. Mt. Sci. 2019, 16, 2783–2792. [Google Scholar] [CrossRef]
- Cattadori, I.M.; Ranci-Ortigosa, G.; Gatto, M.; Hudson, P.J. Is the rock partridge Alectoris graeca saxatilis threatened in the Dolomitic Alps? Anim. Conserv. 2003, 6, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Negri, A.; Pellegrino, I.; Mucci, N.; Randi, E.; Tizzani, P.; Meneguz, P.G.; Malacarne, G. Mitochondrial DNA and microsatellite markers evidence a different pattern of hybridization in red-legged partridge (Alectoris rufa) populations from NW Italy. Eur. J. Wildl. Res. 2013, 59, 407–419. [Google Scholar] [CrossRef] [Green Version]
- Trocchi, V.; Riga, F.; Sorace, A. Piano D’Azione Nazionale per la Coturnice (Alectoris graeca); MATTM—ISPRA: Rome, Italy, 2016. [Google Scholar]
- Bao, X.; Liu, N.; Qu, J.; Wang, X.; An, B.; Wen, L.; Song, S. The phylogenetic position and speciation dynamics of the genus Perdix (Phasianidae, Galliformes). Mol. Phylogenet. Evol. 2010, 56, 840–847. [Google Scholar] [CrossRef]
- Trocchi, V.; Riga, F.; Sorace, A. Piano D’Azione Nazionale per la Starna (Perdix perdix); MATTM—ISPRA: Rome, Italy, 2016. [Google Scholar]
- Pizzirani, C.; Viola, P.; Gabbianelli, F.; Fagotti, A.; Simoncelli, F.; Di Rosa, I.; Salvi, P.; Amici, A.; Lucentini, L. First evidence of heteroplasmy in Grey Partridge (Perdix perdix). Avian Res. 2020, 11, 27. [Google Scholar] [CrossRef]
- Randi, E. Detecting hybridization between wild species and their domesticated relatives. Mol. Ecol. 2008, 17, 285–293. [Google Scholar] [CrossRef]
- BirdLife International Perdix perdix. The IUCN Red List of Threatened Species. Available online: https://doi.org/10.2305/IUCN.UK.2016-3.RLTS.T22678911A85929015.en (accessed on 10 August 2021).
- BirdLife International Species Factsheet: Perdix perdix. Available online: http://datazone.birdlife.org/species/factsheet/grey-partridge-perdix-perdix (accessed on 13 October 2021).
- Liukkonen-Anttila, T.; Uimaniemi, L.; Orell, M.; Lumme, J. Mitochondrial DNA variation and the phylogeography of the grey partridge (Perdix perdix) in Europe: From Pleistocene history to present day populations. J. Evol. Biol. 2002, 15, 971–982. [Google Scholar] [CrossRef] [Green Version]
- Hewitt, G. Post-glacial re-colonization of European biota. Biol. J. Linn. Soc. 1999, 68, 87–112. [Google Scholar] [CrossRef]
- Hewitt, G. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef]
- Randi, E.; Meriggi, A.; Lorenzini, R.; Fusco, G.; Alkon, P.U. Biochemical Analysis of Relationships of Mediterranean Alectoris Partridges. Auk 1992, 109, 358–367. [Google Scholar] [CrossRef]
- Beaumont, M.A.; Rannala, B. The Bayesian revolution in genetics. Nat. Rev. Genet. 2004, 5, 251–261. [Google Scholar] [CrossRef]
- Fumihito, A.; Miyake, T.; Takada, M.; Ohno, S.; Kondo, N. The genetic link between the Chinese bamboo partridge (Bambusicola thoracica) and the chicken and junglefowls of the genus Gallus. Proc. Natl. Acad. Sci. USA 1995, 92, 11053–11056. [Google Scholar] [CrossRef] [Green Version]
- Kerr, K.C.R.; Stoeckle, M.Y.; Dove, C.J.; Weigt, L.A.; Francis, C.M.; Hebert, P.D.N. Comprehensive DNA barcode coverage of North American birds. Mol. Ecol. Notes 2007, 5, 535–543. [Google Scholar] [CrossRef]
- Sorenson, M.D.; Quinn, T.W. A challenge for avian systematics and population biology. Auk 1998, 115, 214–221. [Google Scholar] [CrossRef]
- Xiang, H.; Gao, J.; Yu, B.; Zhou, H.; Cai, D.; Zhang, Y.; Chen, X.; Wang, X.; Hofreiter, M.; Zhao, X. Early Holocene chicken domestication in northern China. Proc. Natl. Acad. Sci. USA 2014, 111, 17564–17569. [Google Scholar] [CrossRef] [Green Version]
- Williams, H.C.; Ormerod, S.J.; Bruford, M.W. Molecular systematics and phylogeography of the cryptic species complex Baetis rhodani (Ephemeroptera, Baetidae). Mol. Phylogenet. Evol. 2006, 40, 370–382. [Google Scholar] [CrossRef]
- Sun, J.; Feng, Z.; Liu, Y. Nucleotide Substitution Pattern in Mitochondrial Cytochrome b Pseudogenes of Ten Species in Galliformes. In Information Technology and Agricultural Engineering; Zhu, E., Sambath, S., Eds.; Springer: Berlin, Germany, 2012; pp. 269–277. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Lethiec, F.; Duroux, P.; Gascuel, O. PHYML Online--a web server for fast maximum likelihood-based phylogenetic inference. Nucleic Acids Res. 2005, 33, W557–W559. [Google Scholar] [CrossRef] [Green Version]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Bech, N.; Novoa, C.; Allienne, J.-F.; Boissier, J.; Bro, E. Quantifying genetic distance between wild and captive strains of the grey partridge Perdix perdix in France: Conservation implications. Biodivers. Conserv. 2020, 29, 609–624. [Google Scholar] [CrossRef]
- Liukkonen, T. Finnish native grey partridge (Perdix perdix) population differs clearly in mitochondrial DNA from the farm stock used for releases. Ann. Zool. Fennici 2006, 43, 271–279. [Google Scholar]
- Strzała, T.; Kowalczyk, A.; Łukaszewicz, E. Reintroduction of the European Capercaillie from the Capercaillie breeding centre in wisła forest district: Genetic assessments of captive and reintroduced populations. PLoS ONE 2015, 10, e0145433. [Google Scholar] [CrossRef]
- Puccioni, M. Cacce e Cacciatori di Toscana: Ricordi e Confessioni; Vallecchi: Firenze, Italy, 1934. [Google Scholar]
- Kan, X.-Z.; Yang, J.-K.; Li, X.-F.; Chen, L.; Lei, Z.-P.; Wang, M.; Qian, C.-J.; Gao, H.; Yang, Z.-Y. Phylogeny of major lineages of galliform birds (Aves: Galliformes) based on complete mitochondrial genomes. Genet. Mol. Res. 2010, 9, 1625–1633. [Google Scholar] [CrossRef] [PubMed]
- Joly, S.; McLenachan, P.A.; Lockhart, P.J. A statistical approach for distinguishing hybridization and incomplete lineage sorting. Am. Nat. 2009, 174, E54–E70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottenburghs, J. Multispecies hybridization in birds. Avian Res. 2019, 10, 20. [Google Scholar] [CrossRef] [Green Version]
- Barr, C.M.; Neiman, M.; Taylor, D.R. Inheritance and recombination of mitochondrial genomes in plants, fungi and animals. New Phytol. 2005, 168, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Matteucci, C.; Toso, F. Note sulla distribuzione e lo status della Starna, Perdix perdix in Italia. Proceedings of Biologia dei Galliformi, Arcavata di Rende, Italy, 16–20 March 1984; Dessì-Fulgheri, F., Mingozzi, T., Eds.; University of Calabria, Department of Ecology: Arcavacata di Rende, Italy, 1986; pp. 29–34. [Google Scholar]
- Priolo, A.; Bocca, M. Coturnice. In Fauna d’Italia. Uccelli.I.; Brichetti, P., De Franceschi, P., Baccetti, N., Eds.; Calderini Edizioni: Bologna, Italy, 1992; pp. 766–777. [Google Scholar]
- Amici, A.; University of Tuscia, Viterbo, Italy; Viola, P.; University of Tuscia, Viterbo, Italy; Rossetti, A.; Sibillini Mountains Nationa Park, Visso, Italy; Morandi, F.; Sibillini Mountains Nationa Park, Visso, Italy. Personal Communication, 2016.
- Zhang, Z.; Chen, J.; Li, L.; Tao, M.; Zhang, C.; Qin, Q.; Xiao, J.; Liu, Y.; Liu, S. Research advances in animal distant hybridization. Sci. China Life Sci. 2014, 57, 889–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, A.; Jain, A.; Sahu, J.; Dubey, A. Interspecies hybridization in animals: An overview. Ann. Anim. Resour. Sci. 2019, 6, 36–42. [Google Scholar]
- Dubost, G.; Royère, J.-Y. Hybridization between ocelot (Felis pardalis) and puma (Felis concolor). Zoo Biol. 1993, 12, 277–283. [Google Scholar] [CrossRef]
- Viola, P.; Primi, R.; Gabbianelli, F.; Danieli, P.P.; Lucentini, L.; Amici, A. Effect of intensive rearing on functional morphology: A case study with the rock partridge (Alectoris graeca graeca). For. Ideas 2019, 25, 275–288. [Google Scholar] [CrossRef]
- Todesco, M.; Pascual, M.A.; Owens, G.L.; Ostevik, K.L.; Moyers, B.T.; Hübner, S.; Heredia, S.M.; Hahn, M.A.; Caseys, C.; Bock, D.G.; et al. Hybridization and extinction. Evol. Appl. 2016, 9, 892–908. [Google Scholar] [CrossRef] [PubMed]
- Wayne, R.K.; Shaffer, H.B. Hybridization and endangered species protection in the molecular era. Mol. Ecol. 2016, 25, 2680–2689. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CMOS | COI | DLOOP | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Groups | Groups | Groups | |||||||||||||
| Morphological species | A A | B P | A A.c. | B A.g. | C A.m. | D A.p. | E A.r. | F P.d. | G P.p. | A A.c. | B A.g | C A.g | D A.r. | E A.r. | F P.p. |
| Alectoris chukar | 5 | 16 | 41 | ||||||||||||
| Alectoris graeca | 57 | 8 | 130 | 20 | 2 | 147 | 1 | 2 | |||||||
| Alectoris magna | 1 | ||||||||||||||
| Alectoris melanocephala | 3 | ||||||||||||||
| Alectoris philbyi | 2 | ||||||||||||||
| Alectoris rufa | 3 | 6 | 40 | 1 | |||||||||||
| Perdix dauurica | 4 | 2 | |||||||||||||
| Perdix hodgsoniae | 3 | ||||||||||||||
| Perdix perdix | 3 | 50 | 6 | 6 | 1 | 195 | 4 | 22 | 720 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fontaneto, D.; Viola, P.; Pizzirani, C.; Chiesa, S.; Rossetti, A.; Amici, A.; Lucentini, L. Mismatches between Morphology and DNA in Italian Partridges May Not Be Explained Only by Recent Artificial Release of Farm-Reared Birds. Animals 2022, 12, 541. https://doi.org/10.3390/ani12050541
Fontaneto D, Viola P, Pizzirani C, Chiesa S, Rossetti A, Amici A, Lucentini L. Mismatches between Morphology and DNA in Italian Partridges May Not Be Explained Only by Recent Artificial Release of Farm-Reared Birds. Animals. 2022; 12(5):541. https://doi.org/10.3390/ani12050541
Chicago/Turabian StyleFontaneto, Diego, Paolo Viola, Claudia Pizzirani, Stefania Chiesa, Alessandro Rossetti, Andrea Amici, and Livia Lucentini. 2022. "Mismatches between Morphology and DNA in Italian Partridges May Not Be Explained Only by Recent Artificial Release of Farm-Reared Birds" Animals 12, no. 5: 541. https://doi.org/10.3390/ani12050541