
Comparison of lncRNA Expression in the Uterus between Periods of Embryo Implantation and Labor in Mice

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Animals and Sample Collection
2.2. Total RNA Isolation, Library Preparation, and Sequencing
2.3. Quality Control and Mapping
2.4. lncRNA Identification
2.5. RNA Expression and Functional Analysis
2.6. Target mRNA Prediction and Functional Analysis of lncRNAs
2.7. GO and KEGG Enrichment Analysis
2.8. Similarities and Differences in mRNAs and lncRNAs between the Two Stages
2.9. Murine Uterine Smooth Muscle Cell Isolation and Culture
2.10. P4 Treatment of MSM Cells
2.11. Validation of LncRNA Expression by qRT-PCR
2.12. Statistical Analysis
3. Results
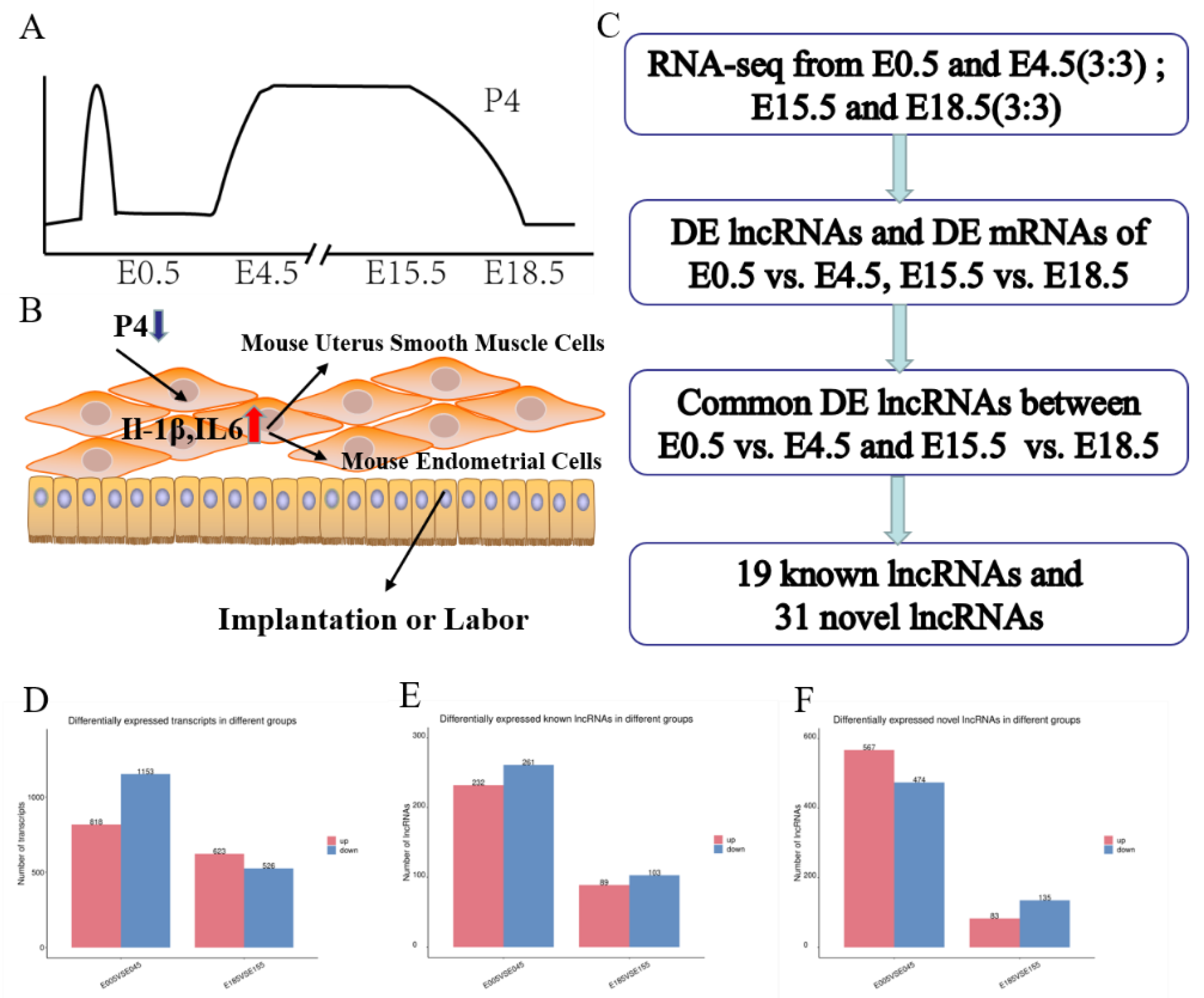
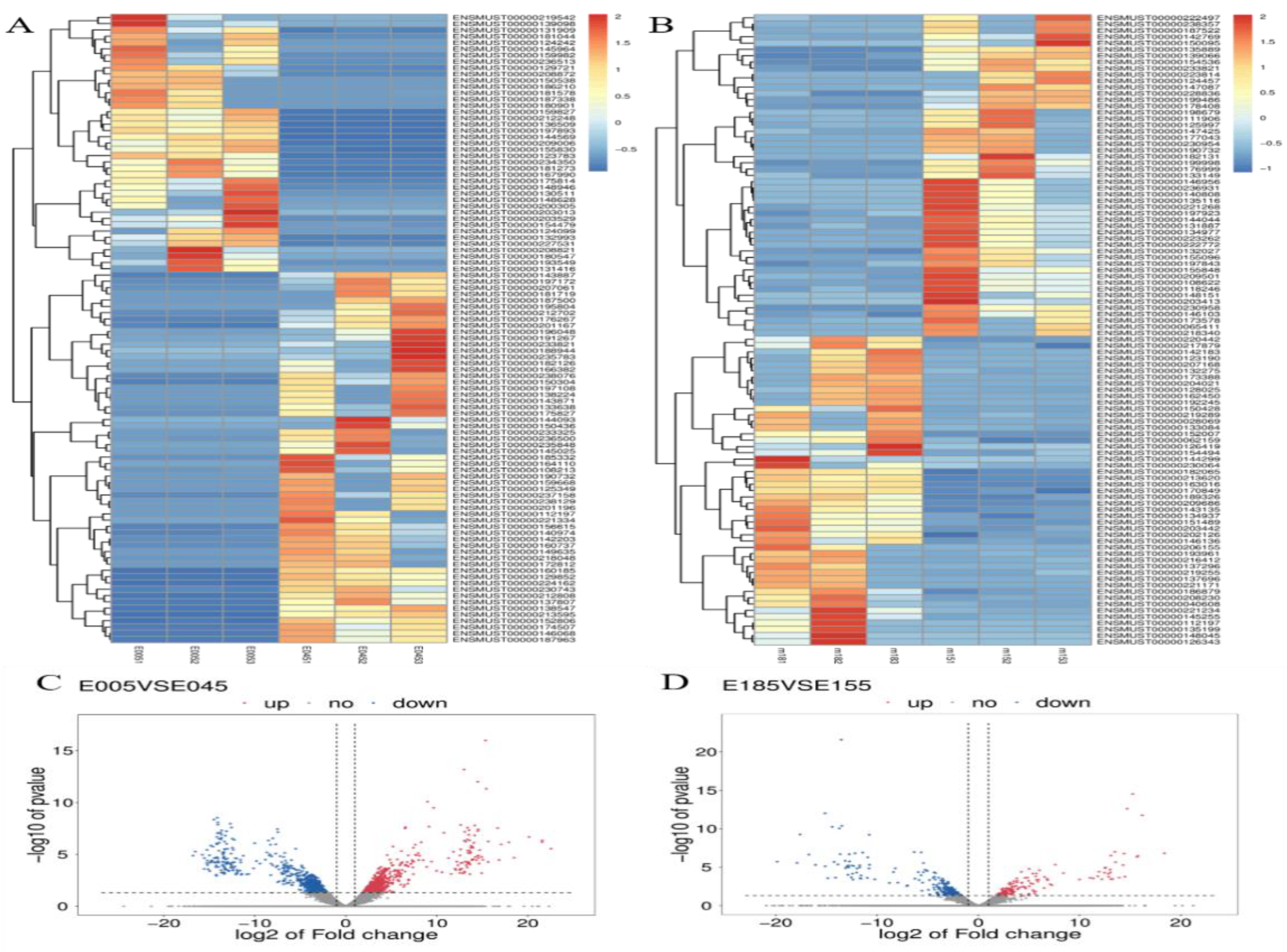
3.1. Sequencing of RNA and Identification of lncRNAs and mRNAs in the Murine Uterus
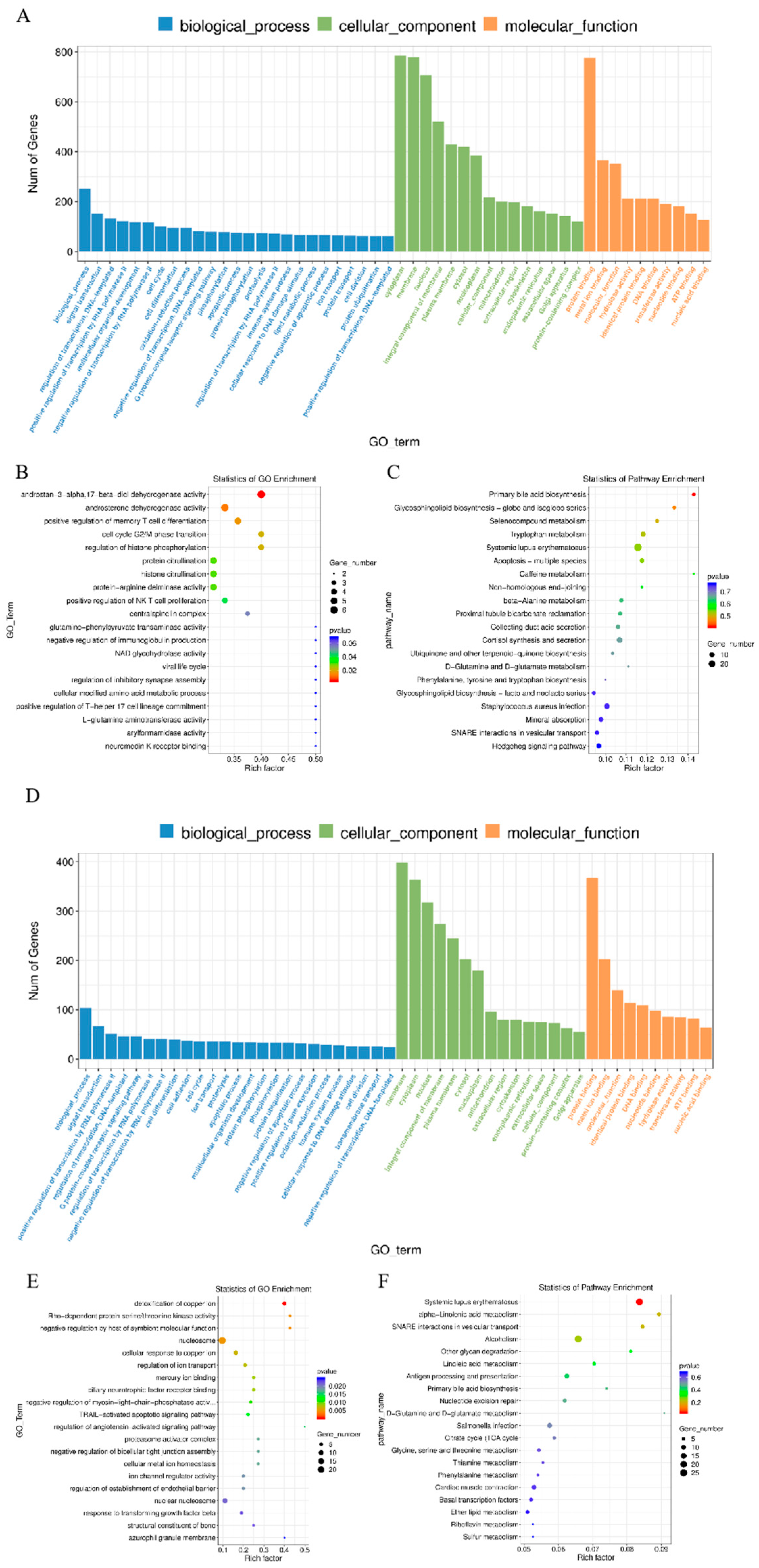
3.2. Enrichment Analysis of DE lncRNAs’ Target mRNAs
3.3. Similar DE mRNAs and DE lncRNAs between the Two Stages
3.4. qRT-PCR Analysis of mRNA Expression in MSM Cells after P4 Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- DeMayo, F.J.; Lydon, J.P. 90 years of progesterone: New insights into progesterone receptor signaling in the endometrium required for embryo implantation. J. Mol. Endocrinol. 2020, 65, T1–T14. [Google Scholar] [CrossRef]
- Sykes, L.; Bennett, P.R. Efficacy of progesterone for prevention of preterm birth. Best Pr. Res. Clin. Obs. Gynaecol. 2018, 52, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Niringiyumukiza, J.D.; Cai, H.; Xiang, W. Prostaglandin E2 involvement in mammalian female fertility: Ovulation, fertilization, embryo development and early implantation. Reprod. Biol. Endocrinol. 2018, 16, 43. [Google Scholar] [CrossRef] [PubMed]
- Bakker, R.; Pierce, S.; Myers, D. The role of prostaglandins E1 and E2, dinoprostone, and misoprostol in cervical ripening and the induction of labor: A mechanistic approach. Arch. Gynecol. Obs. 2017, 296, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Sztachelska, M.; Ponikwicka-Tyszko, D.; Sokolowska, G.; Anisimowicz, S.; Czerniecki, J.; Lebiedzinska, W.; Zbucka-Kretowska, M.; Zygmunt, M.; Wolczynski, S.; Pierzynski, P. Oxytocin antagonism reverses the effects of high oestrogen levels and oxytocin on decidualization and cyclooxygenase activity in endometrial tissues. Reprod. Biomed. Online 2019, 39, 737–744. [Google Scholar] [CrossRef]
- Kernberg, A.; Caughey, A.B. Augmentation of labor: A review of oxytocin augmentation and active management of labor. Obs. Gynecol Clin. North. Am. 2017, 44, 593–600. [Google Scholar] [CrossRef]
- Herington, J.L.; Guo, Y.; Reese, J.; Paria, B.C. Gene profiling the window of implantation: Microarray analyses from human and rodent models. J. Reprod. Health Med. 2016, 2, S19–S25. [Google Scholar] [CrossRef] [Green Version]
- Furcron, A.E.; Romero, R.; Mial, T.N.; Balancio, A.; Panaitescu, B.; Hassan, S.S.; Sahi, A.; Nord, C.; Gomez-Lopez, N. Human Chorionic gonadotropin has anti-inflammatory effects at the maternal-fetal interface and prevents endotoxin-induced preterm birth, but causes dystocia and fetal compromise in mice. Biol. Reprod. 2016, 94, 136. [Google Scholar] [CrossRef] [Green Version]
- Chandran, S.; Cairns, M.T.; O’Brien, M.; O’Connell, E.; Mashayekhi, K.; Smith, T.J. Effects of combined progesterone and 17beta-estradiol treatment on the transcriptome of cultured human myometrial smooth muscle cells. Physiol. Genom. 2016, 48, 50–61. [Google Scholar] [CrossRef] [Green Version]
- Shah, N.M.; Lai, P.F.; Imami, N.; Johnson, M.R. Progesterone-related immune modulation of pregnancy and labor. Front. Endocrinol. 2019, 10, 198. [Google Scholar] [CrossRef] [Green Version]
- Renthal, N.E.; Williams, K.C.; Montalbano, A.P.; Chen, C.C.; Gao, L.; Mendelson, C.R. Molecular regulation of parturition: A myometrial perspective. Cold Spring Harb. Perspect. Med. 2015, 5, a023069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osman, E.K.; Wang, T.; Zhan, Y.; Juneau, C.R.; Morin, S.J.; Seli, E.; Scott, R.T.; Franasiak, J.M. Varying levels of serum estradiol do not alter the timing of the early endometrial secretory transformation. Hum. Reprod. 2020, 35, 1637–1647. [Google Scholar] [CrossRef]
- Amazu, C.; Ma, X.; Henkes, C.; Ferreira, J.J.; Santi, C.M.; England, S.K. Progesterone and estrogen regulate NALCN expression in human myometrial smooth muscle cells. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E441–E452. [Google Scholar] [CrossRef]
- Matsumoto, H. Molecular and cellular events during blastocyst implantation in the receptive uterus: Clues from mouse models. J. Reprod. Dev. 2017, 63, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Massimiani, M.; Lacconi, V.; La Civita, F.; Ticconi, C.; Rago, R.; Campagnolo, L. Molecular signaling regulating endometrium-blastocyst crosstalk. Int. J. Mol. Sci. 2019, 21, 23. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Lopez, N.; StLouis, D.; Lehr, M.A.; Sanchez-Rodriguez, E.N.; Arenas-Hernandez, M. Immune cells in term and preterm labor. Cell Mol. Immunol. 2014, 11, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Shynlova, O.; Nadeem, L.; Zhang, J.; Dunk, C.; Lye, S. Myometrial activation: Novel concepts underlying labor. Placenta 2020, 92, 28–36. [Google Scholar] [CrossRef]
- McCarthy, R.; Martin-Fairey, C.; Sojka, D.K.; Herzog, E.D.; Jungheim, E.S.; Stout, M.J.; Fay, J.C.; Mahendroo, M.; Reese, J.; Herington, J.L.; et al. Mouse models of preterm birth: Suggested assessment and reporting guidelines. Biol. Reprod. 2018, 99, 922–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Dey, S.K. Roadmap to embryo implantation: Clues from mouse models. Nat. Rev. Genet. 2006, 7, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Martin Gonzalez, J.; Morgani, S.M.; Bone, R.A.; Bonderup, K.; Abelchian, S.; Brakebusch, C.; Brickman, J.M. Embryonic stem cell culture conditions support distinct states associated with different developmental stages and potency. Stem Cell Rep. 2016, 7, 177–191. [Google Scholar] [CrossRef] [Green Version]
- Ruane, P.T.; Berneau, S.C.; Koeck, R.; Watts, J.; Kimber, S.J.; Brison, D.R.; Westwood, M.; Aplin, J.D. Apposition to endometrial epithelial cells activates mouse blastocysts for implantation. Mol. Hum. Reprod. 2017, 23, 617–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, B.; Bolumar, D.; Amadoz, A.; Jimenez-Almazan, J.; Valbuena, D.; Vilella, F.; Moreno, I. Identification and characterization of extracellular vesicles and its DNA cargo secreted during murine embryo development. Genes 2020, 11, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edey, L.F.; Georgiou, H.; O’Dea, K.P.; Mesiano, S.; Herbert, B.R.; Lei, K.; Hua, R.; Markovic, D.; Waddington, S.N.; MacIntyre, D.; et al. Progesterone, the maternal immune system and the onset of parturition in the mouse. Biol. Reprod. 2018, 98, 376–395. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.T.; Fu, T.; Zhang, H.Y.; Yang, Z.S.; Zheng, Z.H.; Yang, Z.M. Progesterone-regulated Hsd11b2 as a barrier to balance mouse uterine corticosterone. J. Endocrinol. 2020, 244, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Virgo, B.B.; Bellward, G.D. Serum progesterone levels in the pregnant and postpartum laboratory mouse. Endocrinology 1974, 95, 1486–1490. [Google Scholar] [CrossRef]
- Mendelson, C.R. Minireview: Fetal-maternal hormonal signaling in pregnancy and labor. Mol. Endocrinol. 2009, 23, 947–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, X.H.; Shi, D.Z. Variations of serum estradiol and progesterone levels during consecutive reproductive states in Mongolian gerbils (Meriones unguiculatus). Exp. Anim. 2010, 59, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, H.; Zhou, D.; Shuang, T.; Zhao, H.; Chen, B. Up-regulation of long noncoding RNA SRA promotes cell growth, inhibits cell apoptosis, and induces secretion of estradiol and progesterone in ovarian granular cells of mice. Med. Sci Monit. 2018, 24, 2384–2390. [Google Scholar] [CrossRef] [Green Version]
- Cai, B.; Li, Z.; Ma, M.; Wang, Z.; Han, P.; Abdalla, B.A.; Nie, Q.; Zhang, X. LncRNA-Six1 Encodes a micropeptide to activate Six1 in cis and is involved in cell proliferation and muscle growth. Front. Physiol. 2017, 8, 230. [Google Scholar] [CrossRef]
- Ma, M.; Cai, B.; Jiang, L.; Abdalla, B.A.; Li, Z.; Nie, Q.; Zhang, X. lncRNA-Six1 is a target of miR-1611 that functions as a ceRNA to regulate Six1 protein expression and fiber type switching in chicken myogenesis. Cells 2018, 7, 243. [Google Scholar] [CrossRef] [Green Version]
- Sarropoulos, I.; Marin, R.; Cardoso-Moreira, M.; Kaessmann, H. Developmental dynamics of lncRNAs across mammalian organs and species. Nature 2019, 571, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cho, K.B.; Li, Y.; Tao, G.; Xie, Z.; Guo, B. Long noncoding RNA (lncRNA)-mediated competing endogenous RNA networks provide novel potential biomarkers and therapeutic targets for colorectal cancer. Int. J. Mol. Sci. 2019, 20, 5758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Ou, C. SIRT6/LncRNA-MALAT1: A potential mechanism for treating aging-associated vascular diseases. Int. J. Cardiol. 2020, 319, 139. [Google Scholar] [CrossRef]
- Feng, C.; Shen, J.M.; Lv, P.P.; Jin, M.; Wang, L.Q.; Rao, J.P.; Feng, L. Construction of implantation failure related lncRNA-mRNA network and identification of lncRNA biomarkers for predicting endometrial receptivity. Int. J. Biol. Sci. 2018, 14, 1361–1377. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Elias, M.D.; May, M.; Munuce, M.J.; Bahamondes, L.; Cuasnicu, P.S.; Cohen, D.J. A single post-ovulatory dose of ulipristal acetate impairs post-fertilization events in mice. Mol. Hum. Reprod. 2019, 25, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Yoshinaga, K. A sequence of events in the uterus prior to implantation in the mouse. J. Assist. Reprod. Genet. 2013, 30, 1017–1022. [Google Scholar] [CrossRef] [Green Version]
- Sinawat, S.; Hsaio, W.C.; Flockhart, J.H.; Kaufman, M.H.; Keith, J.; West, J.D. Fetal abnormalities produced after preimplantation exposure of mouse embryos to ammonium chloride. Hum. Reprod. 2003, 18, 2157–2165. [Google Scholar] [CrossRef] [Green Version]
- Van Meer, H.; van Straten, E.M.; Baller, J.F.; van Dijk, T.H.; Plosch, T.; Kuipers, F.; Verkade, H.J. The effects of intrauterine malnutrition on maternal-fetal cholesterol transport and fetal lipid synthesis in mice. Pediatr. Res. 2010, 68, 10–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petropoulos, S.; Gibb, W.; Matthews, S.G. Effect of glucocorticoids on regulation of placental multidrug resistance phosphoglycoprotein (P-gp) in the mouse. Placenta 2010, 31, 803–810. [Google Scholar] [CrossRef]
- Lombardi, A.; Makieva, S.; Rinaldi, S.F.; Arcuri, F.; Petraglia, F.; Norman, J.E. Expression of matrix metalloproteinases in the mouse uterus and human myometrium during pregnancy, labor, and preterm labor. Reprod. Sci. 2018, 25, 938–949. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frazee, A.C.; Pertea, G.; Jaffe, A.E.; Langmead, B.; Salzberg, S.L.; Leek, J.T. Ballgown bridges the gap between transcriptome assembly and expression analysis. Nat. Biotechnol. 2015, 33, 243–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, L.; Zhang, Y.; Ye, Z.Q.; Liu, X.Q.; Zhao, S.Q.; Wei, L.; Gao, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic. Acids Res. 2007, 35, W345–W349. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Luo, H.; Bu, D.; Zhao, G.; Yu, K.; Zhang, C.; Liu, Y.; Chen, R.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic. Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, L.; Li, J.; Zhu, B.; Zhu, H.; Luo, Y.; Wang, Q.; Zuo, J. Analysis of long-non-coding RNAs associated with ethylene in tomato. Gene 2018, 674, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Line, J.; Griffith, K.; Wooldredge, A.; Cruz, Y.P. Progesterone signaling during pregnancy in the lab opossum, Monodelphis domestica. Theriogenology 2019, 136, 101–110. [Google Scholar] [CrossRef]
- Paria, B.C.; Reese, J.; Das, S.K.; Dey, S.K. Deciphering the cross-talk of implantation: Advances and challenges. Science 2002, 296, 2185–2188. [Google Scholar] [CrossRef]
- Egashira, M.; Hirota, Y. Uterine receptivity and embryo-uterine interactions in embryo implantation: Lessons from mice. Reprod. Med. Biol. 2013, 12, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Renthal, N.E.; Chen, C.C.; Williams, K.C.; Gerard, R.D.; Prange-Kiel, J.; Mendelson, C.R. miR-200 family and targets, ZEB1 and ZEB2, modulate uterine quiescence and contractility during pregnancy and labor. Proc. Natl. Acad. Sci. USA 2010, 107, 20828–20833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashiri, A.; Halper, K.I.; Orvieto, R. Recurrent implantation failure-update overview on etiology, diagnosis, treatment and future directions. Reprod. Biol. Endocrinol. 2018, 16, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, R.; Dey, S.K.; Fisher, S.J. Preterm labor: One syndrome, many causes. Science 2014, 345, 760–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamazaki, N.; Uesaka, M.; Nakashima, K.; Agata, K.; Imamura, T. Gene activation-associated long noncoding RNAs function in mouse preimplantation development. Development 2015, 142, 910–920. [Google Scholar] [CrossRef] [Green Version]
- Lv, J.; Liu, H.; Yu, S.; Liu, H.; Cui, W.; Gao, Y.; Zheng, T.; Qin, G.; Guo, J.; Zeng, T.; et al. Identification of 4438 novel lincRNAs involved in mouse pre-implantation embryonic development. Mol. Genet. Genom. 2015, 290, 685–697. [Google Scholar] [CrossRef]
- Taylor, D.H.; Chu, E.T.; Spektor, R.; Soloway, P.D. Long non-coding RNA regulation of reproduction and development. Mol. Reprod. Dev. 2015, 82, 932–956. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Xie, C.; Zhang, Y.; Fan, Z.; Yin, Y.; Blachier, F. Glutamate-glutamine cycle and exchange in the placenta-fetus unit during late pregnancy. Amino Acids 2015, 47, 45–53. [Google Scholar] [CrossRef]
- Xiao, S.; Xu, N.; Ding, Q.; Huang, S.; Zha, Y.; Zhu, H. LncRNA VPS9D1-AS1 promotes cell proliferation in acute lymphoblastic leukemia through modulating GPX1 expression by miR-491-5p and miR-214-3p evasion. Biosci. Rep. 2020, 40, BSR20193461. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, Y.; Li, L.; Liu, Y.; Geng, P.; Li, G.; Song, H. LncRNA SNHG1 enhances cell proliferation, migration, and invasion in cervical cancer. Biochem. Cell Biol. 2018, 96, 38–43. [Google Scholar] [CrossRef]
- Wang, D.W.; Lou, X.Q.; Liu, Z.L.; Zhang, N.; Pang, L. LncRNA SNHG1 protects SH-SY5Y cells from hypoxic injury through miR-140-5p/Bcl-XL axis. Int. J. Neurosci. 2021, 131, 336–345. [Google Scholar] [CrossRef]
- Meng, Q.; Wang, X.; Xue, T.; Zhao, Q.; Wang, W.; Zhao, K. Long noncoding RNA MIR99AHG promotes gastric cancer progression by inducing EMT and inhibiting apoptosis via miR577/FOXP1 axis. Cancer Cell Int. 2020, 20, 414. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Li, H.; Ma, Z.; Dong, G.; Wang, Q.; Wang, S.; Fang, P.; Li, X.; Chen, H.; Liu, T.; et al. MIR99AHG is a noncoding tumor suppressor gene in lung adenocarcinoma. Cell Death Dis. 2021, 12, 424. [Google Scholar] [CrossRef]
- Ai, D.; Yu, F. LncRNA DNM3OS promotes proliferation and inhibits apoptosis through modulating IGF1 expression by sponging MiR-126 in CHON-001 cells. Diagn. Pathol. 2019, 14, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.; Cai, R.; Yu, T.; Zhang, R.; Liu, S.; Guo, X.; Shang, C.; Wang, A.; Jin, Y.; Lin, P. Progesterone-induced RNA Hand2os1 regulates decidualization in mice uteri. Reproduction 2020, 159, 303–314. [Google Scholar] [CrossRef]
- Bork, P. Hundreds of ankyrin-like repeats in functionally diverse proteins: Mobile modules that cross phyla horizontally? Proteins 1993, 17, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.Q.; Liao, S.Y.; Zhuang, X.J.; Han, C.S. Mouse Fem1b interacts with and induces ubiquitin-mediated degradation of Ankrd37. Gene 2011, 485, 153–159. [Google Scholar] [CrossRef]
- Deng, M.; Zhang, W.; Yuan, L.; Tan, J.; Chen, Z. HIF-1a regulates hypoxia-induced autophagy via translocation of ANKRD37 in colon cancer. Exp. Cell Res. 2020, 395, 112175. [Google Scholar] [CrossRef]
- Wu, L.; Duan, Q.; Gao, D.; Wang, Y.; Xue, S.; Li, W.; Lei, M. Zearalenone blocks autophagy flow and induces cell apoptosis during embryo implantation in gilts. Toxicol. Sci. 2020, 175, 126–139. [Google Scholar] [CrossRef]
- Wang, L.; Hu, H.; Morse, A.N.; Han, X.; Bao, J.; Yang, J.; Chen, Y.; Liu, H. Activation of autophagy in human uterine myometrium during labor. Reprod. Sci. 2020, 27, 1665–1672. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Z.; Chen, L.; Cao, M.; Chen, T.; Huang, Y.; Wang, N.; Zhang, B.; Li, F.; Chen, K.; Yuan, C.; et al. Comparison of lncRNA Expression in the Uterus between Periods of Embryo Implantation and Labor in Mice. Animals 2022, 12, 399. https://doi.org/10.3390/ani12030399
Zhao Z, Chen L, Cao M, Chen T, Huang Y, Wang N, Zhang B, Li F, Chen K, Yuan C, et al. Comparison of lncRNA Expression in the Uterus between Periods of Embryo Implantation and Labor in Mice. Animals. 2022; 12(3):399. https://doi.org/10.3390/ani12030399
Chicago/Turabian StyleZhao, Zijiao, Lu Chen, Maosheng Cao, Tong Chen, Yiqiu Huang, Nan Wang, Boqi Zhang, Fangxia Li, Kaimin Chen, Chenfeng Yuan, and et al. 2022. "Comparison of lncRNA Expression in the Uterus between Periods of Embryo Implantation and Labor in Mice" Animals 12, no. 3: 399. https://doi.org/10.3390/ani12030399