
Is There a Link between Vomeronasalitis and Aggression in Stable Social Groups of Female Pigs?

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing Conditions
2.2. Skin Lesions
2.3. Histopathological Analysis
2.4. Statistical Analysis
3. Results
3.1. Skin Lesions
3.2. Histopathological Analysis
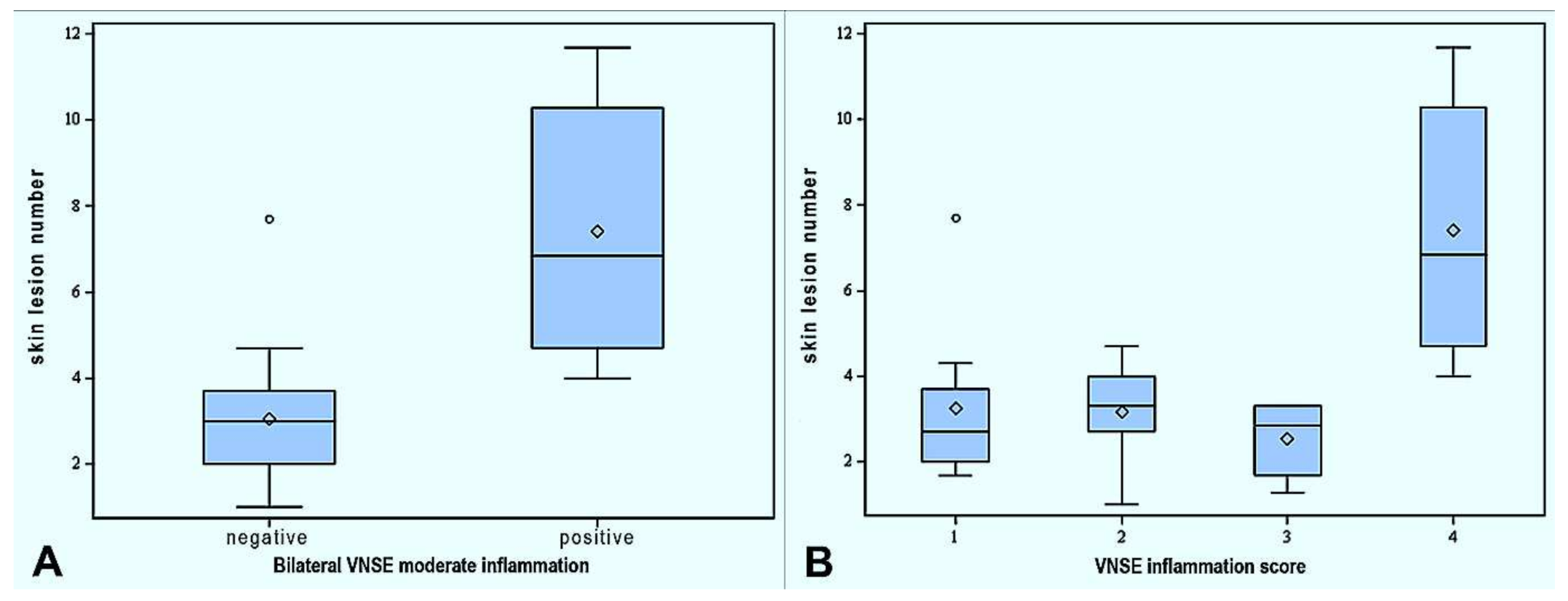
3.3. Association between Skin Lesions and Vomeronasalitis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez-Gómez, A.; Stein, B.; Leinders-Zufall, T.; Chamero, P. Signaling mechanisms and behavioral function of the mouse basal vomeronasal neuroepithelium. Front. Neuroanat. 2014, 26, 8–135. [Google Scholar]
- Salazar, I.; Sanchez-Quinteiro, P.; Lombardero, M.; Cifuentes, J.M. A descriptive and comparative lectin histochemical study of the vomeronasal system in pigs and sheep. J. Anat. 2000, 196, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Guiraudie, G.; Pageat, P.; Cain, A.H.; Madec, I.; Nagnan-Le Meillour, P. Functional characterization of olfactory binding proteins for appeasing compounds and molecular cloning in the vomeronasal organ of pre-pubertal pigs. Chem. Senses 2003, 28, 609–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zancanaro, C. Vomeronasal organ: A short history of discovery and an account of development and morphology in the mouse. In Neurobiology of Chemical Communication, 1st ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 285–296. [Google Scholar]
- Salazar, I.; Sanchez-Quinteiro, P.; Barrios, A.W.; López Amado, M.; Vega, J.A. Anatomy of the olfactory mucosa. Handb Clin. Neurol. 2019, 164, 47–65. [Google Scholar]
- Wysocky, C.; Lepri, J. Consequences of removing the vomeronasal organ. J. Steroid Biochem. Mol. Biol. 1991, 39, 661–669. [Google Scholar] [CrossRef]
- Pankevich, D.; Cherry, J.; Baum, M. Effect of vomeronasal organ removal from male mice on their preference for and neural Fos responses to female urinary odors. Behav. Neurosci. 2006, 120, 925–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiyokawa, Y.; Kikusui, T.; Takeuchi, Y.; Mori, Y. Removal of the vomeronasal organ blocks the stress-induced hyperthermia response to alarm pheromone in male rats. Chem. Senses 2007, 32, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booth, K.K.; Webb, E.C. Effect of blockage of the ducts of the vomeronasal organ on LH plasma levels during the “Whitten Effect” in does. Vet. Med. Int. 2010, 2011, 305468. [Google Scholar] [CrossRef] [Green Version]
- Booth, K.K.; Katz, L.S. Role of the vomeronasal organ in neonatal offspring recognition in sheep. Biol. Reprod. 2000, 63, 953–958. [Google Scholar] [CrossRef] [Green Version]
- Loo, S.K.; Chin, K.N. Lymphoid tissue in the nasal mucosa of primates, with particular reference to intraepithelial lymphocytes. J. Anat. 1974, 117, 249–259. [Google Scholar]
- Torres, M.V.; Ortiz-Leal, I.; Villamayor, P.R.; Ferreiro, A.; Rois, J.L.; Sanchez-Quinteiro, P. Does a third intermediate model for the vomeronasal processing of information exist? Insights from the macropodid neuroanatomy. Brain Struct. Funct. 2021, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.E.; Macri, N.P.; Creasy, D.M. Evaluation of the rabbit nasal cavity in inhalation studies and a comparison with other common laboratory species and man. Toxicol. Pathol. 2011, 39, 893–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babic, N.; Mettenleiter, T.C.; Ugolini, G.; Flamand, A.; Coulon, P. Propagation of pseudorabies virus in the nervous system of the mouse after intranasal inoculation. Virology 1994, 204, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Mechin, V.; Pageat, P.; Teruel, E.; Asproni, P. Histological and Immunohistochemical Characterization of Vomeronasal Organ Aging in Mice. Animals 2021, 11, 1211. [Google Scholar] [CrossRef] [PubMed]
- Asproni, P.; Cozzi, A.; Verin, R.; Lafont-Lecuelle, C.; Bienboire-Frosini, C.; Poli, A.; Pageat, P. Pathology and behaviour in feline medicine: Investigating the link between vomeronasalitis and aggression. J. Feline Med. Surg. 2016, 18, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Arey, D.S.; Edwards, S.A. Factors influencing aggression between sows after mixing and the consequences for welfare and production. Livest. Prod. Sci. 1998, 56, 61–70. [Google Scholar] [CrossRef]
- Stookey, J.M.; Gonyou, H.W. The effects of regrouping on behavioral and production parameters in finishing swine. J. Anim. Sci. 1994, 72, 2804–2811. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.P.; Roehe, R.; D’Eath, R.B.; Ison, S.H.; Farish, M.; Jack, M.C.; Lundeheim, N.; Rydhmer, L.; Lawrence, A.B. Genetic validation of post mixing skin injuries in pigs as an indicator of aggressiveness and the relationship with injuries under more stable social conditions. J. Anim. Sci. 2009, 87, 3076–3082. [Google Scholar] [CrossRef] [Green Version]
- Turner, S.P.; Farnworth, M.J.; White, I.M.S.; Brotherstone, S.; Mendl, M.; Knap, P.; Penny, P.; Lawrence, A.B. The accumulation of skin lesions and their use as a predictor of individual aggressiveness in pigs. Appl. Anim. Behav. Sci. 2006, 96, 245–259. [Google Scholar] [CrossRef]
- Welfare Quality. Welfare Quality® Assessment Protocol for Pigs (Sows and Piglets, Growing and Finishing Pigs); Welfare Quality Consortium: Lelystad, The Netherlands, 2009. [Google Scholar]
- Wyatt, T.D. Pheromones and Animal Behavior: Chemical Signals and Signature Mixes, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Bruce, J.M. Ventilation and Temperature Control Criteria for Pigs. In Environmental Aspects of Housing for Animal Production, 1st ed.; Butterworths: London, UK, 1981; pp. 197–216. [Google Scholar]
- The Humane Society of the United States. An HSUS Report: The Welfare of Animals in the Pig Industry. HSUS Reports: Farm Industry Impacts on Animals; The Humane Society of the United States: Washington, DC, USA, 2010; p. 28. [Google Scholar]
- Dzięcioł, M.; Podgórski, P.; Stańczyk, E.; Szumny, A.; Woszczyło, M.; Pieczewska, B.; Niżański, W.; Nicpoń, J.; Wrzosek, M.A. MRI Features of the Vomeronasal Organ in Dogs (Canis Familiaris). Front Vet Sci. 2020, 7, 159. [Google Scholar] [CrossRef]
- EFSA Panel on Animal Health and Welfare (AHAW). Scientific Opinion on the use of animal-based measures to assess welfare in pigs. EFSA J. 2012, 10, 2512. [Google Scholar] [CrossRef] [Green Version]
- Kern, R.C. Candidate’s Thesis: Chronic sinusitis and anosmia: Pathologic changes in the olfactory mucosa. Laryngoscope 2000, 110, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.P.; Turner, J.; May, L.; Reed, R. A genetic model of chronic rhinosinusitis-associated olfactory inflammation reveals reversible functional impairment and dramatic neuroepithelial reorganization. J. Neurosci. 2010, 30, 2324–2329. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.H.; May, L.; Reed, R.R.; Lane, A.P. Reversible loss of neuronal marker protein expression in a transgenic mouse model for sinusitis-associated olfactory dysfunction. Am. J. Rhinol. Allergy 2011, 24, 192–196. [Google Scholar] [CrossRef] [Green Version]
- Henkin, R.I.; Schmidt, L.; Velicu, I. Interleukin 6 in Hyposmia. JAMA Otolaryngol. Head Neck Surg. 2013, 139, 728–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindberg, C. Animal behaviour and animal welfare. J. Biol. Educ. 1995, 29, 16–22. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Body Zone | Mean | SD |
|---|---|---|
| Ear | 0.3 | 0.36 |
| Front | 1.7 | 1.31 |
| Middle | 1.2 | 1.23 |
| Back | 1.1 | 1.12 |
| Legs | 0.1 | 0.22 |
| All body regions | 4.4 | 2.80 |
| Laterality and Intensity of VNSE Inflammation. (First VNO + Second VNO) | Number of Cases | Percent (%) |
|---|---|---|
| Healthy | 0/38 | 0 |
| Unilateral VNSE chronic inflammation | 13/38 | 34.2 |
| healthy (score 0) + weak inflammation (score 1) | 11/13 | 84.6 |
| healthy (score 0) + moderate inflammation (score 2) | 2/13 | 15.4 |
| Bilateral VNSE chronic inflammation | 25/38 | 65.8 |
| weak (score 1) + weak inflammation (score 1) | 7/25 | 28.0 |
| weak (score 1) + moderate inflammation (score 2) | 6/25 | 24.0 |
| moderate (score 2) + moderate inflammation (score 2) | 12/25 | 48.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asproni, P.; Mainau, E.; Cozzi, A.; Carreras, R.; Bienboire-Frosini, C.; Teruel, E.; Pageat, P. Is There a Link between Vomeronasalitis and Aggression in Stable Social Groups of Female Pigs? Animals 2022, 12, 303. https://doi.org/10.3390/ani12030303
Asproni P, Mainau E, Cozzi A, Carreras R, Bienboire-Frosini C, Teruel E, Pageat P. Is There a Link between Vomeronasalitis and Aggression in Stable Social Groups of Female Pigs? Animals. 2022; 12(3):303. https://doi.org/10.3390/ani12030303
Chicago/Turabian StyleAsproni, Pietro, Eva Mainau, Alessandro Cozzi, Ricard Carreras, Cécile Bienboire-Frosini, Eva Teruel, and Patrick Pageat. 2022. "Is There a Link between Vomeronasalitis and Aggression in Stable Social Groups of Female Pigs?" Animals 12, no. 3: 303. https://doi.org/10.3390/ani12030303