
Intensification of Penaeid Shrimp Culture: An Applied Review of Advances in Production Systems, Nutrition and Breeding

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
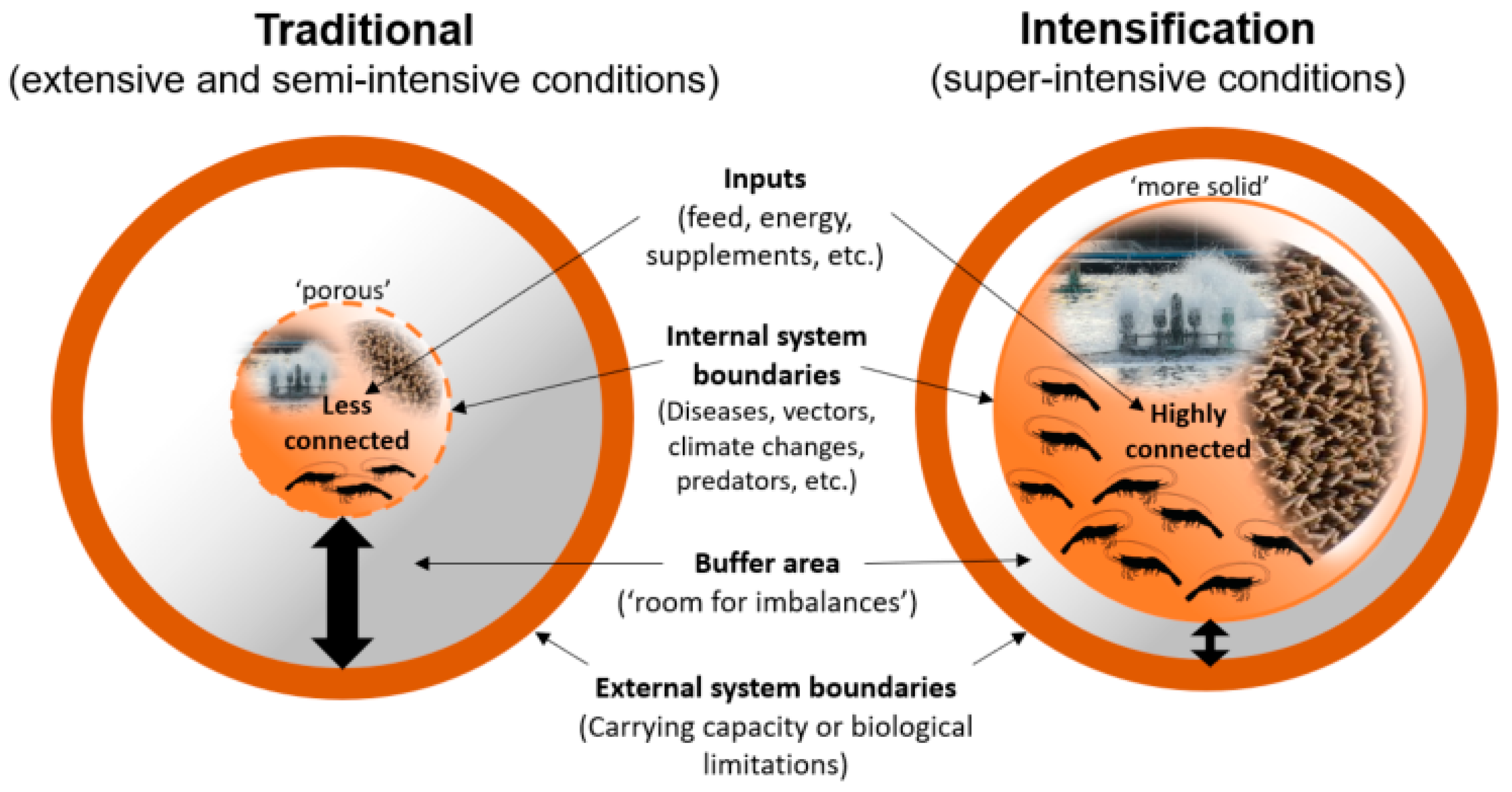
2. Super-Intensive Production Systems and Strategies
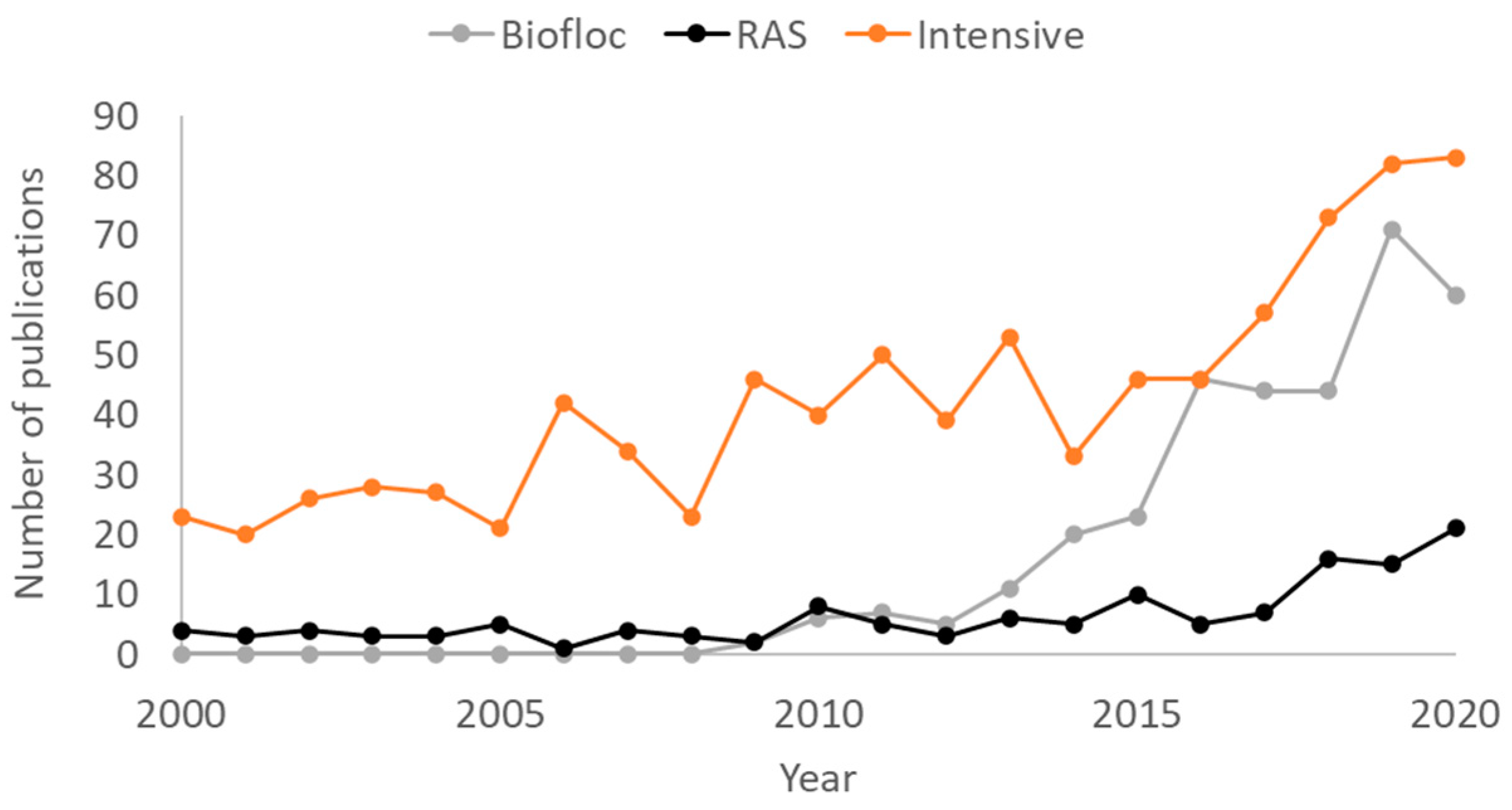
2.1. Biofloc Technology (BFT) and Other Microbial-Based Intensive Systems
2.2. Water Exchange Systems: Flow-Through, RAS and Hybrid Systems
2.3. Nursery Systems

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Production System | Evaluated Aspect | Main Findings | Reference |
|---|---|---|---|
| BFT | Different carbon sources | Lower ammonia concentrations in molasses and starch supplemented systems. | [100] |
| BFT | Stocking density and use of artificial substrates | Substrates increased shrimp yield. | [97] |
| BFT | Different artificial substrates | Needlona® resulted in higher survival and lower concentrations of TSS. | [99] |
| BFT | Stocking densities | Optimum stocking density of 140 post-larvae L−1 | [101] |
| BFT | Different BFT management strategies | Chemoautotrophic strategy reduced Vibrionaceae and improved shrimp performance. | [77] |
| BFT | Feeding frequency | Reducing feeding frequency did not affect shrimp performance. | [102] |
| BFT | Stocking density and water exchange | No water exchange did not affect shrimp growth. | [103] |
| BFT and microalgae-based system | Production system and TSS level | Both systems resulted in similar shrimp performances. | [96] |
| BFT, RAS, and hybrid system | Production system | No significant differences in shrimp performance between the 3 systems. | [85] |
| Hybrid RAS | Stocking density and use of artificial substrate | Higher shrimp yield when using substrates. | [98] |
2.4. Super Intensive Shrimp-Based Integrated Systems
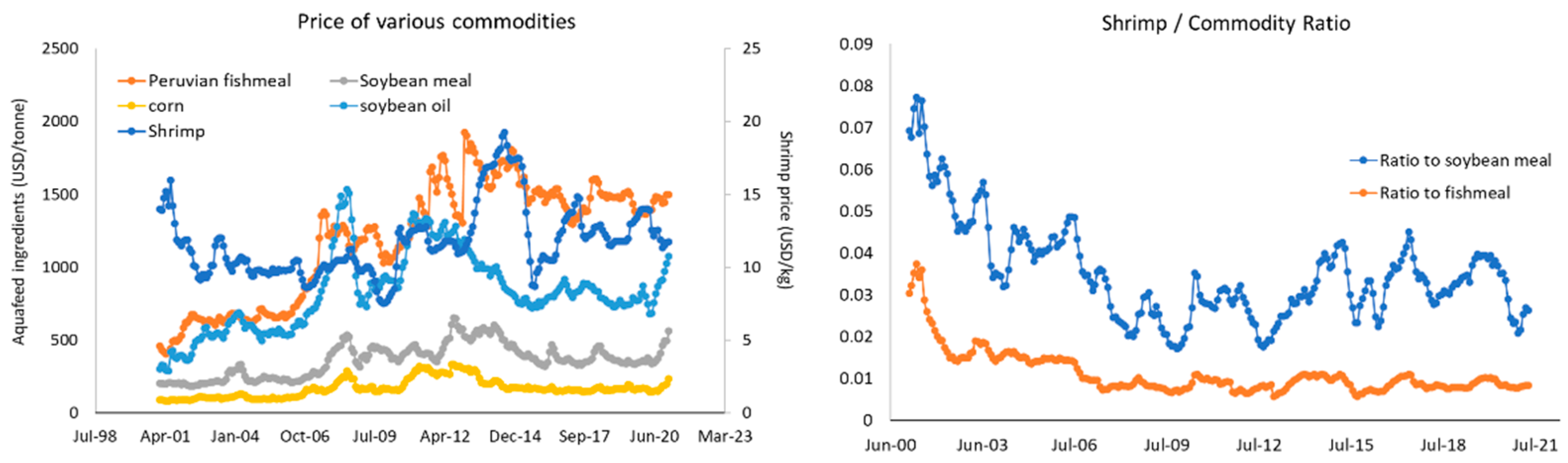
3. The Role of Nutrition in Shrimp Intensification
3.1. Nutrient Requirements in Super-Intensive Systems and Tailored Feeds
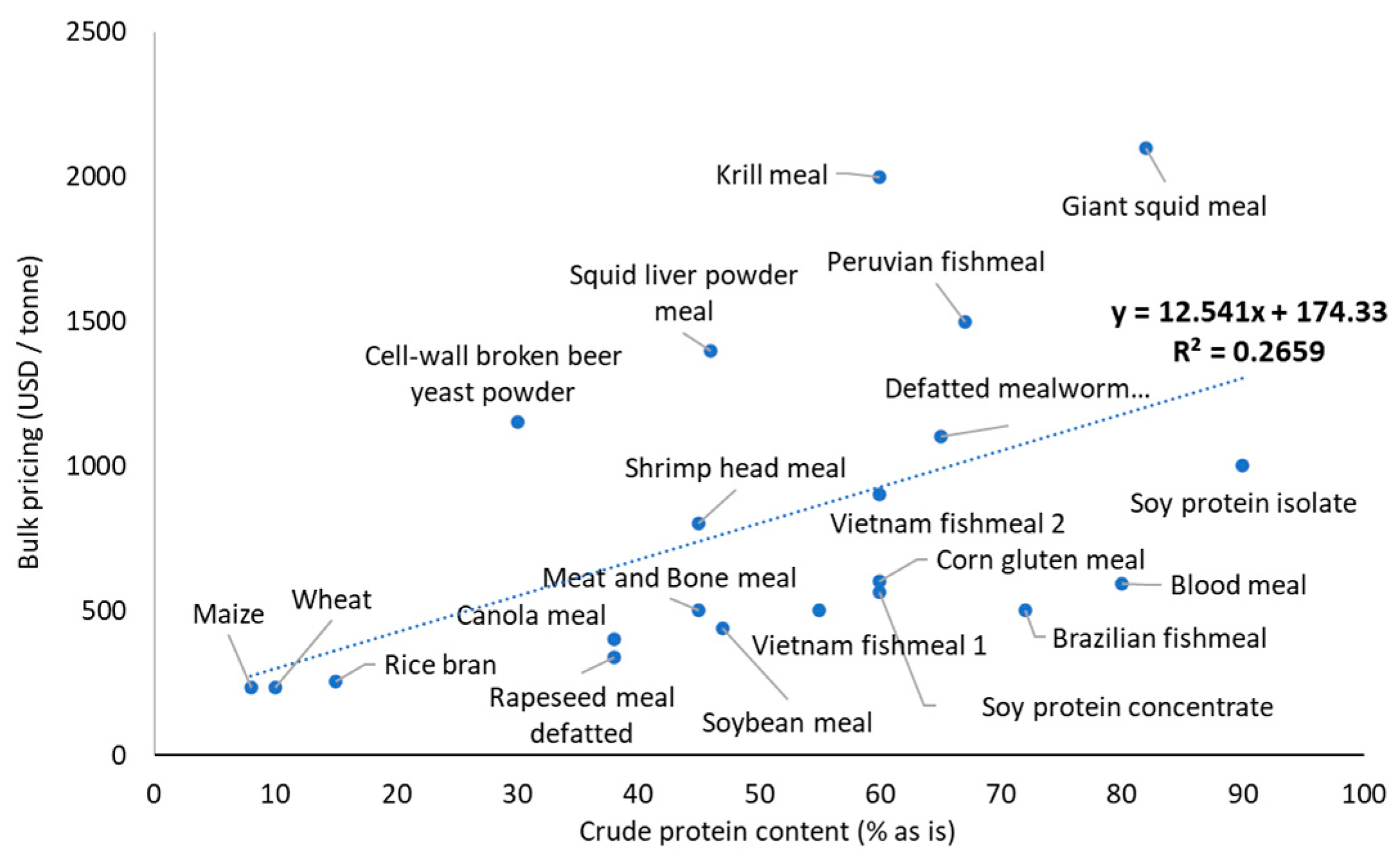
3.2. Digestible Ingredients and Pellet Stability
3.3. Marine and Microbial-Based Growth Promoters
3.4. Feed Additives for Improved Nutrition and Health
3.5. Feed Management in Super-Intensive Systems
4. Breeding and Pathogen Surveillance in Shrimp Intensification
4.1. Domestication and SPF
4.2. Biosecurity and Pathogen Surveillance
4.3. Genetic Improvement
5. Sustainability and Social License
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Fishery Statistical Collection. Global Aquaculture Production. Available online: http://www.fao.org/fishery/statistics/global-aquaculture-production/en (accessed on 2 November 2021).
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action; The State of World Fisheries and Aquaculture (SOFIA); FAO: Rome, Italy, 2020; ISBN 978-92-5-132692-3. [Google Scholar]
- Avnimelech, Y. Biofloc Technology: A Practical Guide Book, 3rd ed.; Avnimelech, Y., Ed.; The World Aquaculture Society: Baton Rouge, LA, USA, 2015; ISBN 978-188880-7226. [Google Scholar]
- Samocha, T. Sustainable Biofloc Systems for Marine Shrimp; Elsevier: Amsterdam, The Netherlands, 2019; ISBN 9780128180402. [Google Scholar]
- Ciji, A.; Akhtar, M.S. Stress management in aquaculture: A review of dietary interventions. Rev. Aquac. 2021, 13, 2190–2247. [Google Scholar] [CrossRef]
- Alday-Sanz, V.; Brock, J.; Flegel, T.W.; McIntosh, R.; Bondad-Reantaso, M.G.; Salazar, M.; Subasinghe, R. Facts, truths and myths about SPF shrimp in Aquaculture. Rev. Aquac. 2020, 12, 76–84. [Google Scholar] [CrossRef]
- Joffre, O.M.; Klerkx, L.; Khoa, T.N.D.D. Aquaculture innovation system analysis of transition to sustainable intensification in shrimp farming. Agron. Sustain. Dev. 2018, 38, 34. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.K.; Guo, Q.; Jang, I.K. Effect of biofloc on the survival and growth of the postlarvae of three penaeids (Litopenaeus vannamei, Fenneropenaeus chinensis, and Marsupenaeus japonicus) and their biofloc feeding efficiencies, as related to the morphological structure of the third maxilliped. J. Crustac. Biol. 2015, 35, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Emerenciano, M.G.C.; Martínez-Córdova, L.R.; Martínez-Porchas, M.; Miranda-Baeza, A. Biofloc Technology (BFT): A Tool for Water Quality Management in Aquaculture. In Water Quality; Intech Open: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Shrimp Insights. SPF L. vannamei Broodstock Report. Available online: https://shrimpinsights.com/report-series (accessed on 9 September 2021).
- Flegel, T.W. Historic emergence, impact and current status of shrimp pathogens in Asia. J. Invertebr. Pathol. 2012, 110, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Lightner, D.V.; Redman, R.M.; Pantoja, C.R.; Tang, K.F.J.; Noble, B.L.; Schofield, P.; Mohney, L.L.; Nunan, L.M.; Navarro, S.A. Historic emergence, impact and current status of shrimp pathogens in the Americas. J. Invertebr. Pathol. 2012, 110, 174–183. [Google Scholar] [CrossRef]
- Flegel, T.W.; Lightner, D.V.; Lo, C.F.; Owens, L. Shrimp Disease Control: Past, Present and Future. In Diseases in Asian Aquaculture VI; Bondad-Reantaso, M.G., Mohan, C.V., Crumlish, M., Subasinghe, R.P., Eds.; Asian Fisheries Society: Manila, Philipines, 2008; p. 505. [Google Scholar]
- Shinn, A.P.; Pratoomyot, J.; Griffiths, D.; Trong, T.Q.; Vu, N.T.; Jiravanichpaisal, P.; Briggs, M. Asian shrimp production and the economic costs of disease. Asian Fish. Sci. 2018, 31, 29–58. [Google Scholar] [CrossRef]
- The World Bank. Reducing Disease Risk in Aquaculture; World Bank Report Number 88257-Glb, Agriculture and Environmental Services Discussion Paper 09; The World Bank: Washington, DC, USA, 2014; 119p. [Google Scholar]
- Moss, S.M.; Moss, D.R.; Arce, S.M.; Lightner, D.V.; Lotz, J.M. The role of selective breeding and biosecurity in the prevention of disease in penaeid shrimp aquaculture. J. Invertebr. Pathol. 2012, 110, 247–250. [Google Scholar] [CrossRef]
- Arnold, S.; Emerenciano, M.G.C.; Cowley, J.A.; Little, B.; Rahman, A.; Perrin, T. Collaboration drives innovations in super-intensive indoor shrimp farming. Global Seafood Alliance, 19 October 2020. [Google Scholar]
- Engle, C.R.; McNevin, A.; Racine, P.; Boyd, C.E.; Paungkaew, D.; Viriyatum, R.; Tinh, H.Q.; Minh, H.N. Economics of Sustainable Intensification of Aquaculture: Evidence from Shrimp Farms in Vietnam and Thailand. J. World Aquac. Soc. 2017, 48, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Aaqillah-Amr, M.A.; Hidir, A.; Azra, M.N.; Ahmad-Ideris, A.R.; Abualreesh, M.H.; Noordiyana, M.N.; Ikhwanuddin, M. Use of pelleted diets in commercially farmed decapods during juvenile stages: A review. Animals 2021, 11, 1761. [Google Scholar] [CrossRef]
- Tacon, A.G.J. Shrimp Farming and the Environment. Thematic Review of Feeds and Feed Management Practices in Shrimp Aquaculture. Consortium Program on Shrimp Farming and the Environment. 2002. Available online: http://library.enaca.org/Shrimp/Case/Thematic/FinalFeed.pdf (accessed on 23 September 2021).
- Castillo-Juárez, H.; Campos-Montes, G.R.; Caballero-Zamora, A.; Montaldo, H.H. Genetic improvement of Pacific white shrimp [Penaeus (Litopenaeus) vannamei]: Perspectives for genomic selection. Front. Genet. 2015, 6, 93. [Google Scholar] [CrossRef] [Green Version]
- Krummenauer, D.; Samocha, T.; Poersch, L.; Lara, G.; Wasielesky, W. The reuse of water on the culture of pacific white shrimp, Litopenaeus vannamei, in BFT system. J. World Aquac. Soc. 2014, 45, 3–14. [Google Scholar] [CrossRef]
- Euroshrimp. #14 European Shrimp Production. Available online: https://www.euroshrimp.net/wp-content/uploads/2021/02/Issue-14-Euroshrimp-Newsletter-European-Shrimp-Prodution.pdf (accessed on 22 February 2021).
- Martinez-Porchas, M.; Martinez-Cordova, L.R. World aquaculture: Environmental impacts and troubleshooting alternatives. Sci. World J. 2012, 2012, 389623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piñeros, J.; Gutiérrez, M.; Coelho, M.; Lapa, M. Aireación en la tecnología Biofloc (BTF): Principios básicos, aplicaciones y perspectivas. Rev. Politécnica 2020, 16, 29–40. [Google Scholar] [CrossRef]
- Krummenauer, D.; Peixoto, S.; Cavalli, R.O.; Poersch, L.H.; Wasielesky, W. Superintensive culture of white shrimp, Litopenaeus vannamei, in a biofloc technology system in Southern Brazil at different stocking densities. J. World Aquac. Soc. 2011, 42, 726–733. [Google Scholar] [CrossRef]
- da Silveira, L.G.P.; Krummenauer, D.; Poersch, L.H.; Rosas, V.T.; Wasielesky, W. Hyperintensive stocking densities for Litopenaeus vannamei grow-out in biofloc technology culture system. J. World Aquac. Soc. 2020, 51, 1290–1300. [Google Scholar] [CrossRef]
- Emerenciano, M.G.C. Conexão Tailândia, presente e futuro! Aquac. Bras. 2018, 14, 60–61. [Google Scholar]
- Ulloa Walker, D.A.; Morales Suazo, M.C.; Emerenciano, M.G.C. Biofloc technology: Principles focused on potential species and the case study of Chilean river shrimp Cryphiops caementarius. Rev. Aquac. 2020, 12, 1759–1782. [Google Scholar] [CrossRef]
- Aguilera-Rivera, D.; Prieto-Davó, A.; Escalante, K.; Chávez, C.; Cuzon, G.; Gaxiola, G. Probiotic effect of FLOC on Vibrios in the pacific white shrimp Litopenaeus vannamei. Aquaculture 2014, 424–425, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Lukwambe, B.; Nicholaus, R.; Zhang, D.; Yang, W.; Zhu, J.; Zheng, Z. Successional changes of microalgae community in response to commercial probiotics in the intensive shrimp (Litopenaeus vannamei Boone) culture systems. Aquaculture 2019, 511, 734257. [Google Scholar] [CrossRef]
- Khademzade, O.; Zakeri, M.; Haghi, M.; Mousavi, S.M. The effects of water additive Bacillus cereus and Pediococcus acidilactici on water quality, growth performances, economic benefits, immunohematology and bacterial flora of whiteleg shrimp (Penaeus vannamei Boone, 1931) reared in earthen ponds. Aquac. Res. 2020, 51, 1759–1770. [Google Scholar] [CrossRef]
- Furtado, P.S.; Poersch, L.H.; Wasielesky, W. Effect of calcium hydroxide, carbonate and sodium bicarbonate on water quality and zootechnical performance of shrimp Litopenaeus vannamei reared in bio-flocs technology (BFT) systems. Aquaculture 2011, 321, 130–135. [Google Scholar] [CrossRef]
- Galkanda-Arachchige, H.; Roy, L.A.; Dahl, S.; James, J.; Kelly, A.M.; Allen Davis, D. Laboratory and on-farm evaluation of low-cost salt mixtures for use during salinity acclimation and the nursery phase of Pacific white shrimp (Litopenaeus vannamei). Aquac. Res. 2020, 51, 3460–3471. [Google Scholar] [CrossRef]
- Galkanda-Arachchige, H.S.C.; Roy, L.A.; Davis, D.A. Evaluation of an alternative salt mixture to culture Pacific white shrimp (Litopenaeus vannamei) in inland aquaculture. Aquac. Res. 2020, 51, 3540–3550. [Google Scholar] [CrossRef]
- Pinto, P.H.O.; Rocha, J.L.; do Vale Figueiredo, J.P.; Carneiro, R.F.S.; Damian, C.; de Oliveira, L.; Seiffert, W.Q. Culture of marine shrimp (Litopenaeus vannamei) in biofloc technology system using artificially salinized freshwater: Zootechnical performance, economics and nutritional quality. Aquaculture 2020, 520, 734960. [Google Scholar] [CrossRef]
- Khanjani, M.H.; Alizadeh, M.; Sharifinia, M. Rearing of the Pacific white shrimp, Litopenaeus vannamei in a biofloc system: The effects of different food sources and salinity levels. Aquac. Nutr. 2020, 26, 328–337. [Google Scholar] [CrossRef]
- Emerenciano, M.; Gaxiola, G.; Cuzo, G. Biofloc Technology (BFT): A Review for Aquaculture Application and Animal Food Industry. In Biomass Now—Cultivation and Utilization; Intech Open: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- De Schryver, P.; Crab, R.; Defoirdt, T.; Boon, N.; Verstraete, W. The basics of bio-flocs technology: The added value for aquaculture. Aquaculture 2008, 277, 125–137. [Google Scholar] [CrossRef]
- Crab, R.; Defoirdt, T.; Bossier, P.; Verstraete, W. Biofloc technology in aquaculture: Beneficial effects and future challenges. Aquaculture 2012, 356–357, 351–356. [Google Scholar] [CrossRef]
- Ray, A.J.; Lewis, B.L.; Browdy, C.L.; Leffler, J.W. Suspended solids removal to improve shrimp (Litopenaeus vannamei) production and an evaluation of a plant-based feed in minimal-exchange, superintensive culture systems. Aquaculture 2010, 299, 89–98. [Google Scholar] [CrossRef]
- Martínez-Córdova, L.R.; Emerenciano, M.; Miranda-Baeza, A.; Martínez-Porchas, M. Microbial-based systems for aquaculture of fish and shrimp: An updated review. Rev. Aquac. 2015, 7, 131–148. [Google Scholar] [CrossRef]
- Panigrahi, A.; Esakkiraj, P.; Jayashree, S.; Saranya, C.; Das, R.R.; Sundaram, M. Colonization of enzymatic bacterial flora in biofloc grown shrimp Penaeus vannamei and evaluation of their beneficial effect. Aquac. Int. 2019, 27, 1835–1846. [Google Scholar] [CrossRef]
- Xu, W.J.; Pan, L.Q.; Sun, X.H.; Huang, J. Effects of bioflocs on water quality, and survival, growth and digestive enzyme activities of Litopenaeus vannamei (Boone) in zero-water exchange culture tanks. Aquac. Res. 2013, 44, 1093–1102. [Google Scholar] [CrossRef]
- Cardona, E.; Lorgeoux, B.; Chim, L.; Goguenheim, J.; Le Delliou, H.; Cahu, C. Biofloc contribution to antioxidant defence status, lipid nutrition and reproductive performance of broodstock of the shrimp Litopenaeus stylirostris: Consequences for the quality of eggs and larvae. Aquaculture 2016, 452, 252–262. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Wille, M.; Lourenço, T.M.; Bossier, P. Biofloc-Based Enhanced Survival of Litopenaeus vannamei Upon AHPND-Causing Vibrio parahaemolyticus Challenge Is Partially Mediated by Reduced Expression of Its Virulence Genes. Front. Microbiol. 2020, 11, 1270. [Google Scholar] [CrossRef] [PubMed]
- Hostins, B.; Wasielesky, W.; Decamp, O.; Bossier, P.; De Schryver, P. Managing input C/N ratio to reduce the risk of Acute Hepatopancreatic Necrosis Disease (AHPND) outbreaks in biofloc systems—A laboratory study. Aquaculture 2019, 508, 60–65. [Google Scholar] [CrossRef]
- Azam, F.; Fenchel, T.; Field, J.G.; Gray, J.S.; Meyer-Reil, L.A.; Thingstad, F. The Ecological Role of Water-Column Microbes in the Sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Nevejan, N.; De Schryver, P.; Wille, M.; Dierckens, K.; Baruah, K.; Van Stappen, G. Bacteria as food in aquaculture: Do they make a difference? Rev. Aquac. 2018, 10, 180–212. [Google Scholar] [CrossRef]
- Pomeroy, L.R. The Ocean’s Food Web, A Changing Paradigm. Bioscience 1974, 24, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Avnimelech, Y. Carbon/nitrogen ratio as a control element in aquaculture systems. Aquaculture 1999, 176, 227–235. [Google Scholar] [CrossRef]
- Hargreaves, J.A. Biofloc Production Systems for Aquaculture Southern Regional Aquaculture Center; SRAC Publication No. 4503; National Institute of Food and Agriculture, US Department of Agriculture: Washington, DC, USA, 2013.
- Ebeling, J.M.; Timmons, M.B.; Bisogni, J.J. Engineering analysis of the stoichiometry of photoautotrophic, autotrophic, and heterotrophic removal of ammonia-nitrogen in aquaculture systems. Aquaculture 2006, 257, 346–358. [Google Scholar] [CrossRef] [Green Version]
- Wasielesky, W.; Atwood, H.; Stokes, A.; Browdy, C.L. Effect of natural production in a zero exchange suspended microbial floc based super-intensive culture system for white shrimp Litopenaeus vannamei. Aquaculture 2006, 258, 396–403. [Google Scholar] [CrossRef]
- Megahed, M.E.; Mohamed, K. Sustainable Growth of Shrimp Aquaculture Through Biofloc Production as Alternative to Fishmeal in Shrimp Feeds. J. Agric. Sci. 2014, 6, 176–188. [Google Scholar] [CrossRef] [Green Version]
- Ballester, E.L.C.; Abreu, P.C.; Cavalli, R.O.; Emerenciano, M.; de Abreu, L.; Wasielesky, W. Effect of practical diets with different protein levels on the performance of Farfantepenaeus paulensis juveniles nursed in a zero exchange suspended microbial flocs intensive system. Aquac. Nutr. 2010, 16, 163–172. [Google Scholar] [CrossRef]
- Jatobá, A.; Da Silva, B.C.; Da Silva, J.S.; Vieira, F.d.N.; Mouriño, J.L.P.; Seiffert, W.Q.; Toledo, T.M. Protein levels for Litopenaeus vannamei in semi-intensive and biofloc systems. Aquaculture 2014, 432, 365–371. [Google Scholar] [CrossRef]
- Scopel, B.R.; Schveitzer, R.; Seiffert, W.Q.; Pierri, V.; Arantes, R.D.F.; Vieira, F.D.N.; Vinatea, L.A. Substituição da farinha de peixe em dietas para camarões marinhos cultivados em sistema bioflocos. Pesqui. Agropecuária Bras. 2011, 46, 928–934. [Google Scholar] [CrossRef]
- Bauer, W.; Prentice-Hernandez, C.; Tesser, M.B.; Wasielesky, W.; Poersch, L.H.S. Substitution of fishmeal with microbial floc meal and soy protein concentrate in diets for the pacific white shrimp Litopenaeus vannamei. Aquaculture 2012, 342–343, 112–116. [Google Scholar] [CrossRef]
- Emerenciano, M.; Cuzon, G.; Goguenheim, J.; Gaxiola, G. Floc contribution on spawning performance of blue shrimp Litopenaeus stylirostris. Aquac. Res. 2012, 44, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Emerenciano, M.; Cuzon, G.; Arévalo, M.; Mascaró Miquelajauregui, M.; Gaxiola, G. Effect of short-term fresh food supplementation on reproductive performance, biochemical composition, and fatty acid profile of Litopenaeus vannamei (Boone) reared under biofloc conditions. Aquac. Int. 2013, 21, 987–1007. [Google Scholar] [CrossRef]
- Emerenciano, M.; Cuzon, G.; Arévalo, M.; Gaxiola, G. Biofloc technology in intensive broodstock farming of the pink shrimp Farfantepenaeus duorarum: Spawning performance, biochemical composition and fatty acid profile of eggs. Aquac. Res. 2014, 45, 1713–1726. [Google Scholar] [CrossRef]
- Braga, A.; Lopes, D.L.A.; Magalhães, V.; Poersch, L.H.; Wasielesky, W. Use of biofloc technology during the pre-maturation period of Litopenaeus vannamei males: Effect of feeds with different protein levels on the spermatophore and sperm quality. Aquac. Res. 2015, 46, 1965–1973. [Google Scholar] [CrossRef]
- De Lorenzo, M.A.; Candia, E.W.S.; Schleder, D.D.; Rezende, P.C.; Seiffert, W.Q.; do Nascimento Vieira, F. Intensive hatchery performance of Pacific white shrimp in the biofloc system under three different fertilization levels. Aquac. Eng. 2016, 72–73, 40–44. [Google Scholar] [CrossRef]
- Xu, W.J.; Pan, L.Q. Enhancement of immune response and antioxidant status of Litopenaeus vannamei juvenile in biofloc-based culture tanks manipulating high C/N ratio of feed input. Aquaculture 2013, 412–413, 117–124. [Google Scholar] [CrossRef]
- Huda, A.S.; Ispinanto, J.; Bahri, F.; Decamp, O. Successful production in semi-biofloc in Indonesia. Aqua Cult. Asia Pac. 2013, 9, 8–12. [Google Scholar]
- Effendy, I.; Al Deen, S.; Chithambaran, S. Semi Intensive and Semi Biofloc Methods for the Culture of Indian White Prawn, Fenneropenaeus indicus in High-density Polyethylene Liner Ponds. HAYATI J. Biosci. 2016, 23, 106–110. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.O.B.; Silva, A.E.M.; Silva, D.A.; Santos, E.P.; Lima, P.C.M.; Andrade, R.J.V.; Silva, S.B.C.; Galvez, A.O. Utilização do sistema simbiótico em berçário de camarões marinhos. Aquac. Bras. 2020, 18, 10–15. [Google Scholar]
- Hussain, A.S.; Mohammad, D.A.; Sallam, W.S.; Shoukry, N.M.; Davis, D.A. Effects of culturing the Pacific white shrimp Penaeus vannamei in “biofloc” vs “synbiotic” systems on the growth and immune system. Aquaculture 2021, 542, 736905. [Google Scholar] [CrossRef]
- Zeng, S.; Khoruamkid, S.; Kongpakdee, W.; Wei, D.; Yu, L.; Wang, H.; Deng, Z.; Weng, S.; Huang, Z.; He, J.; et al. Dissimilarity of microbial diversity of pond water, shrimp intestine and sediment in Aquamimicry system. AMB Express 2020, 10, 180. [Google Scholar] [CrossRef]
- Wright, J. Brazilian Shrimp Producer Wins Global Aquaculture Innovation & Leadership Award. Global Seafood Alliance, 19 October 2015. [Google Scholar]
- Natrah, F.M.I.; Bossier, P.; Sorgeloos, P.; Yusoff, F.M.; Defoirdt, T. Significance of microalgal-bacterial interactions for aquaculture. Rev. Aquac. 2014, 6, 48–61. [Google Scholar] [CrossRef]
- Olier, B.S.; Tubin, J.S.B.; de Mello, G.L.; Martínez-Porchas, M.; Emerenciano, M.G.C. Does vertical substrate could influence the dietary protein level and zootechnical performance of the Pacific white shrimp Litopenaeus vannamei reared in a biofloc system? Aquac. Int. 2020, 28, 1227–1241. [Google Scholar] [CrossRef]
- Ferreira, G.S.; Santos, D.; Schmachtl, F.; Machado, C.; Fernandes, V.; Bögner, M.; Schleder, D.D.; Seiffert, W.Q.; Vieira, F.N. Heterotrophic, chemoautotrophic and mature approaches in biofloc system for Pacific white shrimp. Aquaculture 2021, 533, 736099. [Google Scholar] [CrossRef]
- Furtado, P.S.; Poersch, L.H.; Wasielesky, W. The effect of different alkalinity levels on Litopenaeus vannamei reared with biofloc technology (BFT). Aquac. Int. 2015, 23, 345–358. [Google Scholar] [CrossRef]
- Panigrahi, A.; Das, R.R.; Sivakumar, M.R.; Saravanan, A.; Saranya, C.; Sudheer, N.S.; Kumaraguru Vasagam, K.P.; Mahalakshmi, P.; Kannappan, S.; Gopikrishna, G. Bio-augmentation of heterotrophic bacteria in biofloc system improves growth, survival, and immunity of Indian white shrimp Penaeus indicus. Fish Shellfish Immunol. 2020, 98, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.S.; Silva, V.F.; Martins, M.A.; da Silva, A.C.C.P.; Machado, C.; Seiffert, W.Q.; do Nascimento Vieira, F. Strategies for ammonium and nitrite control in Litopenaeus vannamei nursery systems with bioflocs. Aquac. Eng. 2020, 88, 102040. [Google Scholar] [CrossRef]
- El-Sayed, A.F.M. Use of biofloc technology in shrimp aquaculture: A comprehensive review, with emphasis on the last decade. Rev. Aquac. 2021, 13, 676–705. [Google Scholar] [CrossRef]
- Dalsgaard, J.; Lund, I.; Thorarinsdottir, R.; Drengstig, A.; Arvonen, K.; Pedersen, P.B. Farming different species in RAS in Nordic countries: Current status and future perspectives. Aquac. Eng. 2013, 53, 2–13. [Google Scholar] [CrossRef] [Green Version]
- Taw, N. A look at various intensive shrimp farming systems in Asia. Commercial implementation of biofloc and RAS production systems help control shrimp farming diseases. Global Seafood Alliance, 24 July 2017. [Google Scholar]
- Ray, A.J.; Drury, T.H.; Cecil, A. Comparing clear-water RAS and biofloc systems: Shrimp (Litopenaeus vannamei) production, water quality, and biofloc nutritional contributions estimated using stable isotopes. Aquac. Eng. 2017, 77, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Ley, C. Dutch Company Completes RAS Unit for Shrimp Research. Available online: https://www.hatcheryinternational.com/dutch-company-completes-ras-unit-for-shrimp-research-1359/ (accessed on 15 October 2021).
- Xu, W.; Xu, Y.; Su, H.; Hu, X.; Yang, K.; Wen, G.; Cao, Y. Characteristics of ammonia removal and nitrifying microbial communities in a hybrid biofloc-ras for intensive Litopenaeus vannamei culture: A pilot-scale study. Water 2020, 12, 3000. [Google Scholar] [CrossRef]
- Suantika, G.; Situmorang, M.L.; Kurniawan, J.B.; Pratiwi, S.A.; Aditiawati, P.; Astuti, D.I.; Azizah, F.F.N.; Djohan, Y.A.; Zuhri, U.; Simatupang, T.M. Development of a zero water discharge (ZWD)—Recirculating aquaculture system (RAS) hybrid system for super intensive white shrimp (Litopenaeus vannamei) culture under low salinity conditions and its industrial trial in commercial shrimp urban farming in G. Aquac. Eng. 2018, 82, 12–24. [Google Scholar] [CrossRef]
- Tierney, T.W.; Ray, A.J. Comparing biofloc, clear-water, and hybrid nursery systems (Part I): Shrimp (Litopenaeus vannamei) production, water quality, and stable isotope dynamics. Aquac. Eng. 2018, 82, 73–79. [Google Scholar] [CrossRef]
- Mello, G.L.; (Santa Catarina State University, Laguna, SC, Brazil). Private communication.
- Kaya, D.; Genc, E.; Genc, M.A.; Aktas, M.; Eroldogan, O.T.; Guroy, D. Biofloc technology in recirculating aquaculture system as a culture model for green tiger shrimp, Penaeus semisulcatus: Effects of different feeding rates and stocking densities. Aquaculture 2020, 528, 735526. [Google Scholar] [CrossRef]
- Samocha, T.M.; Prangnell, D.I.; Hanson, T.R.; Treece, G.D.; Morris, T.C.; Castro, L.F.; Staresinic, N. Design and Operation of Super Intensive, Biofloc-Dominated Systems for Indoor Production of the Pacific White Shrimp, Litopenaeus vannamei; The Texas A&M Agrilife Research Experience: Corpus Christi, TX, USA, 2017. [Google Scholar]
- Boyd, C.E.; Jory, D.E.; Chamberlain, G.W. Operating Procedures for Shrimp Farming; Global Aqauculture Alliance: St. Louis, MO, USA, 2006. [Google Scholar]
- Kumaran, M.; Anand, P.R.; Kumar, J.A.; Ravisankar, T.; Paul, J.; Vasagam, K.P.K.; Vimala, D.D.; Raja, K.A. Is Pacific white shrimp (Penaeus vannamei) farming in India is technically efficient?—A comprehensive study. Aquaculture 2017, 468, 262–270. [Google Scholar] [CrossRef]
- Ali, H.; Rahman, M.M.; Jaman, A.; Eltholth, M.; Murray, F.J. Assessment of the efficacy of prophylactic health products on water quality and shrimp (Penaeus monodon) performance at the nursery phase. Aquac. Nutr. 2021, 27, 1173–1180. [Google Scholar] [CrossRef]
- Arnold, S.J.; Coman, F.E.; Jackson, C.J.; Groves, S.A. High-intensity, zero water-exchange production of juvenile tiger shrimp, Penaeus monodon: An evaluation of artificial substrates and stocking density. Aquaculture 2009, 293, 42–48. [Google Scholar] [CrossRef]
- Emerenciano, M.G.C.; Wasielesky Junior, W.; Soares, R.B.; Ballester, E.C.; Izeppi, E.M.; Cavalli, R.O. Crescimento e sobrevivência do camarão-rosa (Farfantepenaeus paulensis) na fase de berçário em meio heterotrófico. Acta Sci. Biol. Sci. 2007, 29, 1–7. [Google Scholar] [CrossRef]
- Emerenciano, M.G.C.; Vinatea, L.; Gálvez, A.; Shule, A.; Stokes, A.; Venero, J.; Haveman, J.; Richardson, J.; Thomas, B.; Leffler, J. Effect of two different diets fish meal based and “organic” plant based diets in Litopenaeus setiferus earlier postlarvae culture under bio-floc, green-water and clear-water conditions. In Proceedings of the CD of abstracts World Aquaculture Society Meeting 2009, Veracruz, Mexico, 5–29 September 2009. [Google Scholar]
- Emerenciano, M.; Ballester, E.L.C.; Cavalli, R.O.; Wasielesky, W. Biofloc technology application as a food source in a limited water exchange nursery system for pink shrimp Farfantepenaeus brasiliensis (Latreille, 1817). Aquac. Res. 2012, 43, 447–457. [Google Scholar] [CrossRef]
- Schveitzer, R.; de Lorenzo, M.A.; do Nascimento Vieira, F.; Pereira, S.A.; Mouriño, J.L.P.; Seiffert, W.Q.; Andreatta, E.R. Nursery of young Litopenaeus vannamei post-larvae reared in biofloc- and microalgae-based systems. Aquac. Eng. 2017, 78, 140–145. [Google Scholar] [CrossRef]
- Legarda, E.C.; Barcelos, S.S.; Redig, J.C.; Ramírez, N.C.B.; Guimarães, A.M.; Santo, C.M.d.E.; Seiffert, W.Q.; Vieira, F.d.N. Effects of stocking density and artificial substrates on yield and water quality in a biofloc shrimp nursery culture. Rev. Bras. Zootec. 2018, 47, e20170060. [Google Scholar] [CrossRef] [Green Version]
- Tierney, T.W.; Fleckenstein, L.J.; Ray, A.J. The effects of density and artificial substrate on intensive shrimp Litopenaeus vannamei nursery production. Aquac. Eng. 2020, 89, 102063. [Google Scholar] [CrossRef]
- Costa Rezende, P.; Dias Schleder, D.; Ventura da Silva, H.; Morais Henriques, F.; de Lorenzo, M.A.; Quadros Seiffert, W.; Roberto Andreatta, E.; Nascimento Vieira, F. do Prenursery of the Pacific white shrimp in a biofloc system using different artificial substrates. Aquac. Eng. 2018, 82, 25–30. [Google Scholar] [CrossRef]
- Khanjani, M.H.; Sajjadi, M.M.; Alizadeh, M.; Sourinejad, I. Nursery performance of Pacific white shrimp (Litopenaeus vannamei Boone, 1931) cultivated in a biofloc system: The effect of adding different carbon sources. Aquac. Res. 2017, 48, 1491–1501. [Google Scholar] [CrossRef]
- Rezende, P.C.; Schleder, D.D.; Seiffert, W.Q.; Andreatta, E.R.; Vieira, F.D.N. Pre-nursery of shrimp post-larvae reared in biofloc system under different stocking densities. Bol. Inst. Pesca 2019, 45, e533. [Google Scholar] [CrossRef]
- Wasielesky, W.; Bezerra, A.; Poersch, L.; Hoffling, F.B.; Krummenauer, D. Effect of feeding frequency on the white shrimp Litopenaeus vannamei during the pilot-scale nursery phase of a superintensive culture in a biofloc system. J. World Aquac. Soc. 2020, 51, 1175–1191. [Google Scholar] [CrossRef]
- Esparza-Leal, H.M.; Ponce-Palafox, J.T.; Álvarez-Ruiz, P.; López-Álvarez, E.S.; Vázquez-Montoya, N.; López-Espinoza, M.; Mejia, M.M.; Gómez-Peraza, R.L.; Nava-Perez, E. Effect of stocking density and water exchange on performance and stress tolerance to low and high salinity by Litopenaeus vannamei postlarvae reared with biofloc in intensive nursery phase. Aquac. Int. 2020, 28, 1473–1483. [Google Scholar] [CrossRef]
- Thakur, D.P.; Lin, C.K. Water quality and nutrient budget in closed shrimp (Penaeus monodon) culture systems. Aquac. Eng. 2003, 27, 159–176. [Google Scholar] [CrossRef]
- Chopin, T.; Robinson, S.M.C.; Troell, M.; Neori, A.; Buschmann, A.H.; Fang, J. Multitrophic Integration for Sustainable Marine Aquaculture. In Encyclopedia of Ecology; Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2008; pp. 2463–2475. [Google Scholar] [CrossRef]
- Dash, P.; Avunje, S.; Tandel, R.S.; Sandeep, K.P.; Panigrahi, A. Biocontrol of Luminous Vibriosis in Shrimp Aquaculture: A Review of Current Approaches and Future Perspectives. Rev. Fish. Sci. Aquac. 2017, 25, 245–255. [Google Scholar] [CrossRef]
- Pinheiro, I.; Arantes, R.; do Espírito Santo, C.M.; do Nascimento Vieira, F.; Lapa, K.R.; Gonzaga, L.V.; Fett, R.; Barcelos-Oliveira, J.L.; Seiffert, W.Q. Production of the halophyte Sarcocornia ambigua and Pacific white shrimp in an aquaponic system with biofloc technology. Ecol. Eng. 2017, 100, 261–267. [Google Scholar] [CrossRef]
- Fierro-Sañudo, J.F.; De Oca, G.A.R.M.; León-Cañedo, J.A.; Alarcón-Silvas, S.G.; Mariscal-Lagarda, M.M.; Díaz-Valdés, T.; Páez-Osuna, F. Production and management of shrimp (Penaeus vannamei) in co-culture with basil (Ocimum basilicum) using two sources of low-salinity water. Lat. Am. J. Aquat. Res. 2018, 46, 63–71. [Google Scholar] [CrossRef]
- Fleurence, J.; Morançais, M.; Dumay, J.; Decottignies, P.; Turpin, V.; Munier, M.; Garcia-Bueno, N.; Jaouen, P. What are the prospects for using seaweed in human nutrition and for marine animals raised through aquaculture? Trends Food Sci. Technol. 2012, 27, 57–61. [Google Scholar] [CrossRef]
- Martins, M.A.; da SILVA, V.F.; Tarapuez, P.R.; Hayashi, L.; Vieira, F.D.N. Cultivation of the seaweed Ulva spp. With effluent from a shrimp biofloc rearing system: Different species and stocking density. Bol. Inst. Pesca 2020, 46, e602. [Google Scholar] [CrossRef]
- Elizondo-González, R.; Quiroz-Guzmán, E.; Escobedo-Fregoso, C.; Magallón-Servín, P.; Peña-Rodríguez, A. Use of seaweed Ulva lactuca for water bioremediation and as feed additive for white shrimp Litopenaeus vannamei. PeerJ 2018, 2018, e4459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poli, M.A.; Legarda, E.C.; de Lorenzo, M.A.; Pinheiro, I.; Martins, M.A.; Seiffert, W.Q.; do Nascimento Vieira, F. Integrated multitrophic aquaculture applied to shrimp rearing in a biofloc system. Aquaculture 2019, 511, 734274. [Google Scholar] [CrossRef]
- Holanda, M.; Santana, G.; Furtado, P.; Rodrigues, R.V.; Cerqueira, V.R.; Sampaio, L.A.; Wasielesky, W.; Poersch, L.H. Evidence of total suspended solids control by Mugil liza reared in an integrated system with pacific white shrimp Litopenaeus vannamei using biofloc technology. Aquac. Rep. 2020, 18, 100479. [Google Scholar] [CrossRef]
- Pinheiro, I.; Carneiro, R.F.S.; Vieira, F.d.N.; Gonzaga, L.V.; Fett, R.; Costa, A.C.d.O.; Magallón-Barajas, F.J.; Seiffert, W.Q. Aquaponic production of Sarcocornia ambigua and Pacific white shrimp in biofloc system at different salinities. Aquaculture 2020, 519, 734918. [Google Scholar] [CrossRef]
- Ge, H.; Ni, Q.; Li, J.; Li, J.-T.; Chen, Z.; Zhao, F.Z. Integration of white shrimp (Litopenaeus vannamei) and green seaweed (Ulva prolifera) in minimum-water exchange aquaculture system. J. Appl. Phycol. 2019, 31, 1425–1432. [Google Scholar] [CrossRef]
- Poli, M.A.; Legarda, E.C.; de Lorenzo, M.A.; Martins, M.A.; do Nascimento Vieira, F. Pacific white shrimp and Nile tilapia integrated in a biofloc system under different fish-stocking densities. Aquaculture 2019, 498, 83–89. [Google Scholar] [CrossRef]
- Legarda, E.C.; Poli, M.A.; Martins, M.A.; Pereira, S.A.; Martins, M.L.; Machado, C.; de Lorenzo, M.A.; do Nascimento Vieira, F. Integrated recirculating aquaculture system for mullet and shrimp using biofloc technology. Aquaculture 2019, 512, 734308. [Google Scholar] [CrossRef]
- Chamorro Legarda, E.; Aranha Martins, M.; Moreira Pereira, P.K.; Siqueira Carneiro, R.F.; Pinheiro, I.C.; Seiffert, W.Q.; Machado, C.; de Lorenzo, M.A.; Vieira, F.d.N. Shrimp rearing in biofloc integrated with different mullet stocking densities. Aquac. Res. 2020, 51, 3571–3581. [Google Scholar] [CrossRef]
- Martins, M.A.; Poli, M.A.; Legarda, E.C.; Pinheiro, I.C.; Carneiro, R.F.S.; Pereira, S.A.; Martins, M.L.; Gonçalves, P.; Schleder, D.D.; do Nascimento Vieira, F. Heterotrophic and mature biofloc systems in the integrated culture of Pacific white shrimp and Nile tilapia. Aquaculture 2020, 514, 734517. [Google Scholar] [CrossRef]
- Omont, A.; Elizondo-González, R.; Quiroz-Guzmán, E.; Escobedo-Fregoso, C.; Hernández-Herrera, R.; Peña-Rodríguez, A. Digestive microbiota of shrimp Penaeus vannamei and oyster Crassostrea gigas co-cultured in integrated multi-trophic aquaculture system. Aquaculture 2020, 521, 735059. [Google Scholar] [CrossRef]
- Poli, M.A.; Martins, M.A.; Pereira, S.A.; Jesus, G.F.A.; Martins, M.L.; Mouriño, J.L.P.; do Nascimento Vieira, F. Increasing stocking densities affect hemato-immunological parameters of Nile tilapia reared in an integrated system with Pacific white shrimp using biofloc technology. Aquaculture 2021, 536, 736497. [Google Scholar] [CrossRef]
- Legarda, E.C.; da Silva, D.; Miranda, C.S.; Pereira, P.K.M.; Martins, M.A.; Machado, C.; de Lorenzo, M.A.; Hayashi, L.; do Nascimento Vieira, F. Sea lettuce integrated with Pacific white shrimp and mullet cultivation in biofloc impact system performance and the sea lettuce nutritional composition. Aquaculture 2021, 534, 736265. [Google Scholar] [CrossRef]
- Burford, M.A.; Sellars, M.J.; Arnold, S.J.; Keys, S.J.; Crocos, P.J.; Preston, N.P. Contribution of the natural biota associated with substrates to the nutritional requirements of the post-larval shrimp, Penaeus esculentus (Haswell), in high-density rearing systems. Aquac. Res. 2004, 35, 508–515. [Google Scholar] [CrossRef] [Green Version]
- Krummenauer, D.; Abreu, P.C.; Poersch, L.; Reis, P.A.C.P.; Suita, S.M.; dos Reis, W.G.; Wasielesky, W. The relationship between shrimp (Litopenaeus vannamei) size and biofloc consumption determined by the stable isotope technique. Aquaculture 2020, 529, 735635. [Google Scholar] [CrossRef]
- IndexMundi. Shrimp Monthly Price—US Dollars per Kilogram. Available online: https://www.indexmundi.com/commodities/?commodity=shrimp&months=60 (accessed on 15 June 2021).
- National Research Council. Nutrient Requirements of Fish and Shrimp; The National Academies Press: Washington, DC, USA, 2011; ISBN 978-0-309-47322-4.
- Martínez-Antonio, E.M.; Racotta, I.S.; Ruvalcaba-Márquez, J.C.; Magallón-Barajas, F. Modulation of stress response and productive performance of Litopenaeus vannamei through diet. PeerJ 2019, 2019, e6850. [Google Scholar] [CrossRef] [Green Version]
- Façanha, F.N.; Oliveira-Neto, A.R.; Figueiredo-Silva, C.; Nunes, A.J.P. Effect of shrimp stocking density and graded levels of dietary methionine over the growth performance of Litopenaeus vannamei reared in a green-water system. Aquaculture 2016, 463, 16–21. [Google Scholar] [CrossRef]
- Xia, S.; Li, Y.; Wang, W.; Rajkumar, M.; Kumaraguru Vasagam, K.P.; Wang, H. Influence of dietary protein levels on growth, digestibility, digestive enzyme activity and stress tolerance in white-leg shrimp, Litopenaeus vannamei (Boone, 1931), reared in high-density tank trials. Aquac. Res. 2010, 41, 1845–1854. [Google Scholar] [CrossRef]
- Cuzon, G.; Lawrence, A.; Gaxiola, G.; Rosas, C.; Guillaume, J. Nutrition of Litopenaeus vannamei reared in tanks or in ponds. Aquaculture 2004, 235, 513–551. [Google Scholar] [CrossRef]
- Ruvalcaba-Márquez, J.C.; Álvarez-Ruíz, P.; Zenteno-Savín, T.; Martínez-Antonio, E.; Goytortúa-Bores, E.; Casillas-Hernández, R.; Mejía-Ruíz, H.; Magallón-Barajas, F.J. Performance, immune response, and oxidative stress parameters of Litopenaeus vannamei fed diets containing varying carbohydrate/protein, lipid/protein, and energy/protein ratios. Aquac. Rep. 2021, 21, 100771. [Google Scholar] [CrossRef]
- International Aquaculture Feed Formulation Database Aquaculture Species Nutritional Specifications Database (ASNS). 2021. Available online: https://www.iaffd.com/ (accessed on 2 November 2021).
- Braga, A.; Magalhães, V.; Hanson, T.; Morris, T.C.; Samocha, T.M. The effects of feeding commercial feed formulated for semi-intensive systems on Litopenaeus vannamei production and its profitability in a hyper-intensive biofloc-dominated system. Aquac. Rep. 2016, 3, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.J.; Morris, T.C.; Samocha, T.M. Effects of two commercial feeds for semi-intensive and hyper-intensive culture and four C/N ratios on water quality and performance of Litopenaeus vannamei juveniles at high density in biofloc-based, zero-exchange outdoor tanks. Aquaculture 2018, 490, 194–202. [Google Scholar] [CrossRef]
- Ju, Z.Y.; Forster, I.; Conquest, L.; Dominy, W. Enhanced growth effects on shrimp (Litopenaeus vannamei) from inclusion of whole shrimp floc or floc fractions to a formulated diet. Aquac. Nutr. 2008, 14, 533–543. [Google Scholar] [CrossRef]
- Xu, W.J.; Pan, L.Q. Dietary protein level and C/N ratio manipulation in zero-exchange culture of Litopenaeus vannamei: Evaluation of inorganic nitrogen control, biofloc composition and shrimp performance. Aquac. Res. 2014, 45, 1842–1851. [Google Scholar] [CrossRef]
- Panjaitan, P. Field and Laboratory Study of Penaeus monodon Culture with Zero Water Exchange and Limited Water Exchange Model Using Molasses as a Carbon Source; Charles Darwin University: Darwin, NT, Australia, 2004; Available online: https://ris.cdu.edu.au/ws/portalfiles/portal/22706397/Thesis_CDU_6467_Panjaitan_P.pdf (accessed on 29 October 2021).
- Pinho, S.M.; Emerenciano, M.G.C. Sensorial attributes and growth performance of whiteleg shrimp (Litopenaeus vannamei) cultured in biofloc technology with varying water salinity and dietary protein content. Aquaculture 2021, 540, 736727. [Google Scholar] [CrossRef]
- Jatobá, A.; Vieira, F.d.N.; da Silva, B.C.; Soares, M.; Mouriño, J.L.P.; Seiffert, W.Q. Replacement of fishmeal for soy protein concentrate in diets for juvenile Litopenaeus vannamei in biofloc-based rearing system. Rev. Bras. Zootec. 2017, 46, 705–713. [Google Scholar] [CrossRef] [Green Version]
- Shao, J.; Wang, B.; Liu, M.; Jiang, K.; Wang, L.; Wang, M. Replacement of fishmeal by fermented soybean meal could enhance the growth performance but not significantly influence the intestinal microbiota of white shrimp Litopenaeus vannamei. Aquaculture 2019, 504, 354–360. [Google Scholar] [CrossRef]
- Naylor, R.L.; Hardy, R.W.; Bureau, D.P.; Chiu, A.; Elliott, M.; Farrell, A.P.; Forster, I.; Gatlin, D.M.; Goldburg, R.J.; Hua, K.; et al. Feeding aquaculture in an era of finite resources. Proc. Natl. Acad. Sci. USA 2009, 106, 18040. [Google Scholar] [CrossRef] [Green Version]
- Turchini, G.M.; Trushenski, J.T.; Glencross, B.D. Thoughts for the Future of Aquaculture Nutrition: Realigning Perspectives to Reflect Contemporary Issues Related to Judicious Use of Marine Resources in Aquafeeds. N. Am. J. Aquac. 2019, 81, 13–39. [Google Scholar] [CrossRef]
- Francis, G.; Makkar, H.P.S.; Becker, K. Antinutritional factors present in plant-derived alternate fish feed ingredients and their effects in fish. Agriculture 2001, 199, 197–227. [Google Scholar] [CrossRef]
- Richard, L.; Surget, A.; Rigolet, V.; Kaushik, S.J.; Geurden, I. Availability of essential amino acids, nutrient utilisation and growth in juvenile black tiger shrimp, Penaeus monodon, following fishmeal replacement by plant protein. Aquaculture 2011, 322–323, 109–116. [Google Scholar] [CrossRef]
- Lim, C.; Dominy, W. Evaluation of soybean meal as a replacement for marine animal protein in diets for shrimp (Penaeus vannamei). Aquaculture 1990, 87, 53–63. [Google Scholar] [CrossRef]
- Amaya, E.; Davis, D.A.; Rouse, D.B. Alternative diets for the Pacific white shrimp Litopenaeus vannamei. Aquaculture 2007, 262, 419–425. [Google Scholar] [CrossRef]
- Van Nguyen, N.; Hoang, L.; Van Khanh, T.; Duy Hai, P.; Hung, L.T. Utilization of fermented soybean meal for fishmeal substitution in diets of Pacific white shrimp (Litopenaeus vannamei). Aquac. Nutr. 2018, 24, 1092–1100. [Google Scholar] [CrossRef]
- McLean, E.; Barrows, F.T.; Craig, S.R.; Alfrey, K.; Tran, L. Complete replacement of fishmeal by soybean and poultry meals in Pacific whiteleg shrimp feeds: Growth and tolerance to EMS/AHPND and WSSV challenge. Aquaculture 2020, 527, 735383. [Google Scholar] [CrossRef]
- Carvalho, R.A.P.L.F.d.; Ota, R.H.; Kadry, V.O.; Tacon, A.G.J.; Lemos, D. Apparent digestibility of protein, energy and amino acids of six protein sources included at three levels in diets for juvenile white shrimp Litopenaeus vannamei reared in high performance conditions. Aquaculture 2016, 465, 223–234. [Google Scholar] [CrossRef]
- Liu, X.H.; Ye, J.D.; Kong, J.H.; Wang, K.; Wang, A.L. Apparent digestibility of 12 protein-origin ingredients for pacific white shrimp Litopenaeus vannamei. N. Am. J. Aquac. 2012, 75, 90–98. [Google Scholar] [CrossRef]
- Lemos, D.; Lawrence, A.L.; Siccardi, A.J. Prediction of apparent protein digestibility of ingredients and diets by in vitro pH-stat degree of protein hydrolysis with species-specific enzymes for juvenile Pacific white shrimp Litopenaeus vannamei. Aquaculture 2009, 295, 89–98. [Google Scholar] [CrossRef]
- Qiu, X.; Nguyen, L.; Davis, D.A. Apparent digestibility of animal, plant and microbial ingredients for Pacific white shrimp Litopenaeus vannamei. Aquac. Nutr. 2018, 24, 930–939. [Google Scholar] [CrossRef]
- Yang, Q.; Zhou, X.; Zhou, Q.; Tan, B.; Chi, S.; Dong, X. Apparent digestibility of selected feed ingredients for white shrimp Litopenaeus vannamei, Boone. Aquac. Res. 2009, 41, 78–86. [Google Scholar] [CrossRef]
- Cruz-Suárez, L.E.; Tapia-Salazar, M.; Villarreal-Cavazos, D.; Beltran-Rocha, J.; Nieto-López, M.G.; Lemme, A.; Ricque-Marie, D. Apparent dry matter, energy, protein and amino acid digestibility of four soybean ingredients in white shrimp Litopenaeus vannamei juveniles. Aquaculture 2009, 292, 87–94. [Google Scholar] [CrossRef]
- Davis, D.A.; Arnold, C.R. Evaluation of five carbohydrate sources for Penaeus vannamei. Aquaculture 1993, 114, 285–292. [Google Scholar] [CrossRef]
- Nunes, A.J.P.; Sá, M.V.C.; Browdy, C.L.; Vazquez-Anon, M. Practical supplementation of shrimp and fish feeds with crystalline amino acids. Aquaculture 2014, 431, 20–27. [Google Scholar] [CrossRef] [Green Version]
- Huai, M.Y.; Liu, Y.J.; Tian, L.X.; Deng, S.X.; Xu, A.L.; Gao, W.; Yang, H.J. Effect of dietary protein reduction with synthetic amino acids supplementation on growth performance, digestibility, and body composition of juvenile Pacific white shrimp, Litopenaeus vannamei. Aquac. Int. 2010, 18, 255–269. [Google Scholar] [CrossRef]
- Sabry-Neto, H.; Lemos, D.; Raggi, T.; Nunes, A.J.P. Effects of soy protein ratio, lipid content and minimum level of krill meal in plant-based diets over the growth and digestibility of the white shrimp, Litopenaeus vannamei. Aquac. Nutr. 2017, 23, 293–303. [Google Scholar] [CrossRef]
- Sookying, D.; Davis, D.A.; Soller Dias da Silva, F. A review of the development and application of soybean-based diets for Pacific white shrimp Litopenaeus vannamei. Aquac. Nutr. 2013, 19, 441–448. [Google Scholar] [CrossRef]
- Davis, D.A.; Arnold, C.R.; McCallum, I. Nutritional value of feed peas (Pisum sativum) in practical diet formulations for Litopenaeus vannamei. Aquac. Nutr. 2002, 8, 87–94. [Google Scholar] [CrossRef]
- Simon, C.J.; Truong, H.; Habilay, N.; Hines, B. Feeding behaviour and bioavailability of essential amino acids in shrimp Penaeus monodon fed fresh and leached fishmeal and fishmeal-free diets. Animals 2021, 11, 847. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, X.; Xu, H.; Sun, W.; Leng, X. Substitute of soy protein concentrate for fish meal in diets of white shrimp (Litopenaeus vannamei Boone). Aquac. Int. 2017, 25, 1303–1315. [Google Scholar] [CrossRef]
- Bulbul, M.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Abdul Kader, M. Growth performance of juvenile kuruma shrimp, Marsupenaeus japonicus (Bate) fed diets replacing fishmeal with soybean meal. Aquac. Res. 2015, 46, 572–580. [Google Scholar] [CrossRef]
- Terrazas-Fierro, M.; Civera-Cerecedo, R.; Ibarra-Martínez, L.; Goytortúa-Bores, E.; Herrera-Andrade, M.; Reyes-Becerra, A. Apparent digestibility of dry matter, protein, and essential amino acid in marine feedstuffs for juvenile whiteleg shrimp Litopenaeus vannamei. Aquaculture 2010, 308, 166–173. [Google Scholar] [CrossRef]
- Fleckenstein, L.J.; Tierney, T.W.; Fisk, J.C.; Ray, A.J. The effects of different solids and biological filters in intensive pacific white shrimp (Litopenaeus vannamei) production systems. Aquac. Eng. 2020, 91, 102120. [Google Scholar] [CrossRef]
- Dominy, W.G.; Cody, J.J.; Terpstra, J.H.; Obaldo, L.G.; Chai, M.K.; Takamori, T.I.; Larsen, B.; Forster, I.P. A comparative study of the physical and biological properties of commercially-available binders for shrimp feeds. J. Appl. Aquac. 2003, 14, 81–99. [Google Scholar] [CrossRef]
- Simon, C.J. The effect of carbohydrate source, inclusion level of gelatinised starch, feed binder and fishmeal particle size on the apparent digestibility of formulated diets for spiny lobster juveniles, Jasus edwardsii. Aquaculture 2009, 296, 329–336. [Google Scholar] [CrossRef]
- Cruz-Ricque, L.E.; Guillaume, J.; Cuzon, G. Squid Protein Effect on Growth of Four Penaeid Shrimp. J. World Aquac. Soc. 1987, 18, 209–217. [Google Scholar] [CrossRef]
- Kuhn, D.D.; Lawrence, A.L.; Boardman, G.D.; Patnaik, S.; Marsh, L.; Flick, G.J. Evaluation of two types of bioflocs derived from biological treatment of fish effluent as feed ingredients for Pacific white shrimp, Litopenaeus vannamei. Aquaculture 2010, 303, 28–33. [Google Scholar] [CrossRef]
- Qiu, X.; Davis, D.A. Evaluation of dried fermented biomass as a feed ingredient in plant-based practical diets for juvenile Pacific white shrimp Litopenaeus vannamei. Aquac. Nutr. 2018, 24, 383–391. [Google Scholar] [CrossRef]
- Qiu, X.; Tian, H.Y.; Davis, D.A. Evaluation of a novel bacterial biomass as a substitution for soybean meal in plant-based practical diets for Pacific white shrimp Litopenaeus vannamei. Aquac. Nutr. 2018, 24, 872–885. [Google Scholar] [CrossRef]
- Sudaryono, A.; Sukardi, P.; Yudiarti, E.; Hardi, E.H.; Hastuti, S.; Susilowati, T. Potential of using tropical brown macroalgae Sargassum cristaefolium meal in the diets for juvenile white shrimp (Litopenaeus vannamei). IOP Conf. Ser. Earth Environ. Sci. 2018, 144, 012049. [Google Scholar] [CrossRef] [Green Version]
- Simon, C.J.; Truong, H.H.; Noble, T.H.; Osborne, S.A.; Wynne, J.W.; Wade, N.M. Microbial biomass, marine invertebrate meals and feed restriction influence the biological and gut microbiota response of shrimp Penaeus monodon. Aquaculture 2020, 520, 734679. [Google Scholar] [CrossRef]
- Cruz-Ricque, L.E.; Guillaume, J.; Van Wormhoudt, A. Effect of squid extracts on time course appearance of glucose and free amino acids in haemolymph in Penaeus japonicus after feeding: Preliminary results. Aquaculture 1989, 76, 57–65. [Google Scholar] [CrossRef]
- Córdova-Murueta, J.H.; García-Carreño, F.L. Nutritive value of squid and hydrolyzed protein supplement in shrimp feed. Aquaculture 2002, 210, 371–384. [Google Scholar] [CrossRef]
- Smith, D.M.; Tabrett, S.J.; Barclay, M.C.; Irvin, S.J. The efficacy of ingredients included in shrimp feeds to stimulate intake. Aquac. Nutr. 2005, 11, 263–272. [Google Scholar] [CrossRef]
- Williams, K.C.; Smith, D.M.; Barclay, M.C.; Tabrett, S.J.; Riding, G. Evidence of a growth factor in some crustacean-based feed ingredients in diets for the giant tiger shrimp Penaeus monodon. Aquaculture 2005, 250, 377–390. [Google Scholar] [CrossRef]
- Anand, P.S.S.; Kohli, M.P.S.; Kumar, S.; Sundaray, J.K.; Roy, S.D.; Venkateshwarlu, G.; Sinha, A.; Pailan, G.H. Effect of dietary supplementation of biofloc on growth performance and digestive enzyme activities in Penaeus monodon. Aquaculture 2014, 418–419, 108–115. [Google Scholar] [CrossRef]
- Shyne Anand, P.S.; Kohli, M.P.S.; Dam Roy, S.; Sundaray, J.K.; Kumar, S.; Sinha, A.; Pailan, G.H.; Sukham, M. kumar Effect of dietary supplementation of periphyton on growth, immune response and metabolic enzyme activities in Penaeus monodon. Aquac. Res. 2015, 46, 2277–2288. [Google Scholar] [CrossRef]
- Glencross, B.; Arnold, S.; Irvin, S. Bioactive factors in microbial biomass have the capacity to offset reductions in the level of protein in the diet of black tiger shrimp, Penaeus monodon. Aquaculture 2015, 446, 74–79. [Google Scholar] [CrossRef]
- Ju, Z.Y.; Forster, I.P.; Dominy, W.G. Effects of supplementing two species of marine algae or their fractions to a formulated diet on growth, survival and composition of shrimp (Litopenaeus vannamei). Aquaculture 2009, 292, 237–243. [Google Scholar] [CrossRef]
- Derby, C.D.; Elsayed, F.H.; Williams, S.A.; González, C.; Choe, M.; Bharadwaj, A.S.; Chamberlain, G.W. Krill meal enhances performance of feed pellets through concentration-dependent prolongation of consumption by Pacific white shrimp, Litopenaeus vannamei. Aquaculture 2016, 458, 13–20. [Google Scholar] [CrossRef]
- Truong, H.H.; Hines, B.M.; Rombenso, A.N.; Simon, C.J. Feed intake, gastro-intestinal transit and haemolymph free amino acids in the shrimp Penaeus monodon are influenced by marine meal supplementation. Aquaculture 2021, 533, 736171. [Google Scholar] [CrossRef]
- Arnold, S.; Smullen, R.; Briggs, M.; West, M.; Glencross, B. The combined effect of feed frequency and ration size of diets with and without microbial biomass on the growth and feed conversion of juvenile Penaeus monodon. Aquac. Nutr. 2016, 22, 1340–1347. [Google Scholar] [CrossRef]
- Rombenso, A.N.; Duong, M.H.; Hines, B.M.; Mã, T.; Simon, C.J. The marine microbial biomass, NovacqTM, a useful feed additive for postlarvae and juvenile Litopenaeus vannamei. Aquaculture 2021, 530, 735959. [Google Scholar] [CrossRef]
- Nunes, A.J.P.; Sá, M.V.C.; Sabry-Neto, H. Growth performance of the white shrimp, Litopenaeus vannamei, fed on practical diets with increasing levels of the Antarctic krill meal, Euphausia superba, reared in clear- versus green-water culture tanks. Aquac. Nutr. 2011, 17, e511–e520. [Google Scholar] [CrossRef]
- Modor Intelligence. Global Fish, Fish Feed, Fish Feed Additives, Shrimp, Shrimp Feed, and Shrimp Feed Additives Market—Growth, Trends, Covid-19 Impact, and Forecasts (2021–2026). Available online: https://www.mordorintelligence.com/industry-reports/global-fish-shrimp-feed-additives-market-industry (accessed on 15 November 2021).
- Zhang, M.; Sun, Y.; Chen, K.; Yu, N.; Zhou, Z.; Chen, L.; Du, Z.; Li, E. Characterization of the intestinal microbiota in Pacific white shrimp, Litopenaeus vannamei, fed diets with different lipid sources. Aquaculture 2014, 434, 449–455. [Google Scholar] [CrossRef]
- Barman, D.; Nen, P. Immunostimulants for Aquaculture Health Management. J. Mar. Sci. Res. Dev. 2013, 3, 134. [Google Scholar] [CrossRef] [Green Version]
- Bachère, E. Shrimp immunity and disease control. Aquaculture 2000, 191, 3–11. [Google Scholar] [CrossRef]
- Nugroho, R.A.; Fotedar, R. Dietary organic selenium improves growth, survival and resistance to Vibrio mimicus in cultured marron, Cherax cainii (Austin, 2002). Fish Shellfish Immunol. 2013, 35, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Pan, L.; Huang, F.; Wang, C.; Xu, W. Effects of Different Carbon Sources on Bioactive Compound Production of Biofloc, Immune Response, Antioxidant Level, and Growth Performance of Litopenaeus vannamei in Zero-water Exchange Culture Tanks. J. World Aquac. Soc. 2016, 47, 566–576. [Google Scholar] [CrossRef]
- Zokaeifar, H.; Balcázar, J.L.; Saad, C.R.; Kamarudin, M.S.; Sijam, K.; Arshad, A.; Nejat, N. Effects of Bacillus subtilis on the growth performance, digestive enzymes, immune gene expression and disease resistance of white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2012, 33, 683–689. [Google Scholar] [CrossRef] [Green Version]
- Nimrat, S.; Tanutpongpalin, P.; Sritunyalucksana, K.; Boonthai, T.; Vuthiphandchai, V. Enhancement of growth performance, digestive enzyme activities and disease resistance in black tiger shrimp (Penaeus monodon) postlarvae by potential probiotics. Aquac. Int. 2013, 21, 655–666. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Esteban, M.Á. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Sirirustananun, N.; Chen, J.C.; Lin, Y.C.; Yeh, S.T.; Liou, C.H.; Chen, L.L.; Sim, S.S.; Chiew, S.L. Dietary administration of a Gracilaria tenuistipitata extract enhances the immune response and resistance against Vibrio alginolyticus and white spot syndrome virus in the white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2011, 31, 848–855. [Google Scholar] [CrossRef]
- Wongprasert, K.; Rudtanatip, T.; Praiboon, J. Immunostimulatory activity of sulfated galactans isolated from the red seaweed Gracilaria fisheri and development of resistance against white spot syndrome virus (WSSV) in shrimp. Fish Shellfish Immunol. 2014, 36, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Baruah, K.; Nguyen, D.V.; Smagghe, G.; Vossen, E.; Bossier, P. Phloroglucinol-mediated Hsp70 production in crustaceans: Protection against Vibrio parahaemolyticus in Artemia franciscana and Macrobrachium rosenbergii. Front. Immunol. 2018, 9, 1091. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.Y.; Loo, K.W.; Wong, W.L. Synergistic Antimicrobial Effect of a Seaweed-Probiotic Blend Against Acute Hepatopancreatic Necrosis Disease (AHPND)-Causing Vibrio parahaemolyticus. Probiotics Antimicrob. Proteins 2020, 12, 906–917. [Google Scholar] [CrossRef]
- Rungrassamee, W.; Maibunkaew, S.; Karoonuthaisiri, N.; Jiravanichpaisal, P. Application of bacterial lipopolysaccharide to improve survival of the black tiger shrimp after Vibrio harveyi exposure. Dev. Comp. Immunol. 2013, 41, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Anuta, J.D.; Buentello, A.; Patnaik, S.; Lawrence, A.L.; Mustafa, A.; Hume, M.E.; Gatlin, D.M.; Kemp, M.C. Effect of Dietary Supplementation of Acidic Calcium Sulfate (Vitoxal) on Growth, Survival, Immune Response and Gut Microbiota of the Pacific White Shrimp, Litopenaeus vannamei. J. World Aquac. Soc. 2011, 42, 834–844. [Google Scholar] [CrossRef]
- He, W.; Rahimnejad, S.; Wang, L.; Song, K.; Lu, K.; Zhang, C. Effects of organic acids and essential oils blend on growth, gut microbiota, immune response and disease resistance of Pacific white shrimp (Litopenaeus vannamei) against Vibrio parahaemolyticus. Fish Shellfish Immunol. 2017, 70, 164–173. [Google Scholar] [CrossRef]
- Rombenso, A.N.; Truong, H.; Simon, C. Dietary butyrate alone or in combination with succinate and fumarate improved survival, feed intake, growth and nutrient retention efficiency of juvenile Penaeus monodon. Aquaculture 2020, 528, 735492. [Google Scholar] [CrossRef]
- Ng, W.-K.; Koh, C.-B.; Teoh, C.-Y.; Romano, N. Farm-raised tiger shrimp, Penaeus monodon, fed commercial feeds with added organic acids showed enhanced nutrient utilization, immune response and resistance to Vibrio harveyi challenge. Aquaculture 2015, 449, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Holt, C.C.; Bass, D.; Stentiford, G.D.; van der Giezen, M. Understanding the role of the shrimp gut microbiome in health and disease. J. Invertebr. Pathol. 2020, 186, 107387. [Google Scholar] [CrossRef]
- Noble, T.H.; Truong, H.H.; Rombenso, A.; Simon, C.J.; Wade, N.M.; Wynne, J.W. Effect of sample type and the use of high or low fishmeal diets on bacterial communities in the gastrointestinal tract of Penaeus monodon. Appl. Microbiol. Biotechnol. 2021, 105, 1301–1313. [Google Scholar] [CrossRef]
- Gainza, O.; Romero, J. Effect of mannan oligosaccharides on the microbiota and productivity parameters of Litopenaeus vannamei shrimp under intensive cultivation in Ecuador. Sci. Rep. 2020, 10, 2719. [Google Scholar] [CrossRef]
- Hauton, C.; Hudspith, M.; Gunton, L. Future prospects for prophylactic immune stimulation in crustacean aquaculture—The need for improved metadata to address immune system complexity. Dev. Comp. Immunol. 2015, 48, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Ullman, C.; Rhodes, M.; Hanson, T.; Cline, D.; Davis, D.A. Effects of Four Different Feeding Techniques on the Pond Culture of Pacific White Shrimp, Litopenaeus vannamei. J. World Aquac. Soc. 2019, 50, 54–64. [Google Scholar] [CrossRef] [Green Version]
- Nunes, A.J.P.; Sabry-Neto, H.; Oliveira-Neto, S.; Burri, L. Feed preference and growth response of juvenile Litopenaeus vannamei to supplementation of marine chemoattractants in a fishmeal-challenged diet. J. World Aquac. Soc. 2019, 50, 1048–1063. [Google Scholar] [CrossRef]
- Wade, N.M.; Bourne, N.; Simon, C.J. Influence of marker particle size on nutrient digestibility measurements and particle movement through the digestive system of shrimp. Aquaculture 2018, 491, 273–280. [Google Scholar] [CrossRef]
- Ullman, C.; Rhodes, M.A.; Davis, D.A. The effects of feed leaching on the growth of Pacific white shrimp Litopenaeus vannamei in a green-water tank system. Aquac. Res. 2019, 50, 3074–3077. [Google Scholar] [CrossRef]
- Reis, J.; Novriadi, R.; Swanepoel, A.; Jingping, G.; Rhodes, M.; Davis, D.A. Optimizing feed automation: Improving timer-feeders and on demand systems in semi-intensive pond culture of shrimp Litopenaeus vannamei. Aquaculture 2020, 519, 734759. [Google Scholar] [CrossRef]
- Darodes de Tailly, J.; Keitel, J.; Owen, M.A.G.; Alcaraz-Calero, J.M.; Alexander, M.E.; Sloman, K.A. Monitoring methods of feeding behaviour to answer key questions in penaeid shrimp feeding. Rev. Aquac. 2021, 13, 1828–1843. [Google Scholar] [CrossRef]
- Guo, J.; Duan, M.; Qiu, X.; Masagounder, K.; Davis, D.A. Characterization of methionine uptake and clearance in the hemolymph of Pacific white shrimp Litopenaeus vannamei. Aquaculture 2020, 526, 735351. [Google Scholar] [CrossRef]
- Van, T.P.T.H.; Rhodes, M.A.; Zhou, Y.; Davis, D.A. Feed management for Pacific white shrimp Litopenaeus vannamei under semi-intensive conditions in tanks and ponds. Aquac. Res. 2017, 48, 5346–5355. [Google Scholar] [CrossRef]
- Venero, J.A.; Davis, D.A.; Rouse, D.B. Variable feed allowance with constant protein input for the pacific white shrimp Litopenaeus vannamei reared under semi-intensive conditions in tanks and ponds. Aquaculture 2007, 269, 490–503. [Google Scholar] [CrossRef]
- Glencross, B.; Tabrett, S.; Irvin, S.; Wade, N.; Anderson, M.; Blyth, D.; Smith, D.; Coman, G.; Preston, N. An analysis of the effect of diet and genotype on protein and energy utilization by the black tiger shrimp, Penaeus monodon—Why do genetically selected shrimp grow faster? Aquac. Nutr. 2013, 19, 128–138. [Google Scholar] [CrossRef]
- Walker, P.J.; Mohan, C.V. Viral disease emergence in shrimp aquaculture: Origins, impact and the effectiveness of health management strategies. Rev. Aquac. 2009, 1, 125–154. [Google Scholar] [CrossRef]
- Wyban, J. Selective Breeding of Penaeus vannamei: Impact on World Aquaculture and Lessons for Future. J. Coast. Res. 2019, 86, 1–5. [Google Scholar] [CrossRef]
- Lightner, D.V. Biosecurity in Shrimp Farming: Pathogen Exclusion through Use of SPF Stock and Routine Surveillance. J. World Aquac. Soc. 2005, 36, 229–248. [Google Scholar] [CrossRef]
- Moss, S.M.; Arce, S.M.; Moss, D.R.; Otoshi, C.A. Disease prevention strategies for penaeid shrimp culture. In Proceedings of the 32nd US–Japan Cooperative Program in Natural Resources Aquaculture Panel Symposium, Santa Barbara, CA, USA, 17, 18 and 20 November 2003. [Google Scholar]
- Moss, S.M.; Moss, D.R. Selective breeding of penaeid shrimp. In Shellfish Safety and Quality; Shumway, S.E., Rodrick, G.E., Eds.; Woodhead Publishing: Sawston, UK, 2009; pp. 425–452. [Google Scholar]
- Vazquez, L.; Alpuche, J.; Maldonado, G.; Agundis, C.; Pereyra-Morales, A.; Zenteno, E. Immunity mechanisms in crustaceans. Innate Immun. 2009, 15, 179–188. [Google Scholar] [CrossRef]
- Noble, T.H. Selective Breeding for Tolerance to Gill-Associated Virus in the Black Tiger Shrimp, Penaeus monodon. Ph.D. Thesis, James Cook University, Townsville, QSL, Australia, 2019; p. 149. [Google Scholar]
- Lillehammer, M.; Bangera, R.; Salazar, M.; Vela, S.; Erazo, E.C.; Suarez, A.; Cock, J.; Rye, M.; Robinson, N.A. Genomic selection for white spot syndrome virus resistance in whiteleg shrimp boosts survival under an experimental challenge test. Sci. Rep. 2020, 10, 20571. [Google Scholar] [CrossRef] [PubMed]
- Robalino, J.; Browdy, C.L.; Prior, S.; Metz, A.; Parnell, P.; Gross, P.; Warr, G. Induction of antiviral immunity by double-stranded RNA in a marine invertebrate. J. Virol. 2004, 78, 10442–10448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robalino, J.; Bartlett, T.; Shepard, E.; Prior, S.; Jaramillo, G.; Scura, E.; Chapman, R.W.; Gross, P.S.; Browdy, C.L.; Warr, G.W. Double-Stranded RNA Induces Sequence-Specific Antiviral Silencing in Addition to Nonspecific Immunity in a Marine Shrimp: Convergence of RNA Interference and Innate Immunity in the Invertebrate Antiviral Response? J. Virol. 2005, 79, 13561–13571. [Google Scholar] [CrossRef] [Green Version]
- Robalino, J.; Bartlett, T.C.; Chapman, R.W.; Gross, P.S.; Browdy, C.L.; Warr, G.W. Double-stranded RNA and antiviral immunity in marine shrimp: Inducible host mechanisms and evidence for the evolution of viral counter-responses. Dev. Comp. Immunol. 2007, 31, 539–547. [Google Scholar] [CrossRef]
- Westenberg, M.; Heinhuis, B.; Zuidema, D.; Vlak, J.M. siRNA injection induces sequence-independent protection in Penaeus monodon against white spot syndrome virus. Virus Res. 2005, 114, 133–139. [Google Scholar] [CrossRef]
- Tirasophon, W.; Roshorm, Y.; Panyim, S. Silencing of yellow head virus replication in penaeid shrimp cells by dsRNA. Biochem. Biophys. Res. Commun. 2005, 334, 102–107. [Google Scholar] [CrossRef]
- Assavalapsakul, W.; Smith, D.R.; Panyim, S. Identification and characterization of a Penaeus monodon lymphoid cell-expressed receptor for the yellow head virus. J. Virol. 2006, 80, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Yodmuang, S.; Tirasophon, W.; Roshorm, Y.; Chinnirunvong, W.; Panyim, S. YHV-protease dsRNA inhibits YHV replication in Penaeus monodon and prevents mortality. Biochem. Biophys. Res. Commun. 2006, 341, 351–356. [Google Scholar] [CrossRef]
- Sellars, M.; Rao, M.; Arnold, S.; Wade, N.; Cowley, J. Penaeus monodon is protected against gill-associated virus by muscle injection but not oral delivery of bacterially expressed dsRNAs. Dis. Aquat. Organ. 2011, 95, 19–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Han, F.; Zhang, X. Silencing shrimp white spot syndrome virus (WSSV) genes by siRNA. Antiviral Res. 2007, 73, 126–131. [Google Scholar] [CrossRef]
- Flegel, T.W. New circular DNA vaccines and heritable, anti-viral immunity possible for shrimp. Two decades of study on shrimp, insect viral defense mechanisms open the way for vaccine production and heritable anti-viral immunity. Global Seafood Alliance, 21 June 2021. [Google Scholar]
- Roy, S.; Bossier, P.; Norouzitallab, P.; Vanrompay, D. Trained immunity and perspectives for shrimp aquaculture. Rev. Aquac. 2020, 12, 2351–2370. [Google Scholar] [CrossRef]
- Cock, J.; Gitterle, T.; Salazar, M.; Rye, M. Breeding for disease resistance of Penaeid shrimps. Aquaculture 2009, 286, 1–11. [Google Scholar] [CrossRef]
- Doyle, R.W. An approach to the quantitative analysis of domestication selection in aquaculture. Aquaculture 1983, 33, 167–185. [Google Scholar] [CrossRef]
- Coman, G.; Sellars, M.; Henshall, J.; Preston, N. Penaeus monodon Breeding: Genetic Improvement using Novel Technologies and Techniques. In Proceedings of the World Aquaculture Society Meeting 2013, Nashville, TN, USA, 21–25 February 2013. [Google Scholar]
- Coman, G.J.; Argue, B.J. Domestication and selective breeding of Penaeus monodon. In Proceedings of the World Aquaculture Society Meeting, Las Vegas, NV, USA, 22–26 February 2016. [Google Scholar]
- Flegel, T.W. A future vision for disease control in shrimp aquaculture. J. World Aquac. Soc. 2019, 50, 249–266. [Google Scholar] [CrossRef]
- Sithigorngul, P.; Rukpratanporn, S.; Chaivisuthangkura, P.; Sridulyakul, P.; Longyant, S. Simultaneous and rapid detection of white spot syndrome virus and yellow head virus infection in shrimp with a dual immunochromatographic strip test. J. Virol. Methods 2011, 173, 85–91. [Google Scholar] [CrossRef]
- Stentiford, G.D.; Sritunyalucksana, K.; Flegel, T.W.; Bryony, A.; Williams, P.; Withyachumnarnkul, B.; Itsathitphaisarn, O.; Bass, D. New Paradigms to Help Solve the Global Aquaculture Disease Crisis. PLoS Pathog. 2017, 13, e1006160. [Google Scholar] [CrossRef]
- Moss, S.M.; Doyle, R.W.; Lightner, D.V. Breeding shrimp for disease resistance: Challenges and opportunities for improvement. In Diseases in Asian Aquaculture V; Walker, P.J., Lester, R.J.G., Doyle, R.W., Eds.; Fish Health Section, Asian Fisheries Society: Manila, Philipines, 2005; pp. 379–393. [Google Scholar]
- Argue, B.J.; Arce, S.M.; Lotz, J.M.; Moss, S.M. Selective breeding of Pacific white shrimp (Litopenaeus vannamei) for growth and resistance to Taura Syndrome Virus. Aquaculture 2002, 204, 447–460. [Google Scholar] [CrossRef]
- White, B.L.; Schofield, P.J.; Poulos, B.T.; Lightner, D.V. A laboratory challenge method for estimating Taura syndrome virus resistance in selected lines of Pacific white shrimp Litopenaeus vannamei. J. World Aquac. Soc. 2002, 33, 341–348. [Google Scholar] [CrossRef]
- Gitterle, T.; Salte, R.; Gjerde, B.; Cock, J.; Johansen, H.; Salazar, M.; Lozano, C.; Rye, M. Genetic (co)variation in resistance to White Spot Syndrome Virus (WSSV) and harvest weight in Penaeus (Litopenaeus) vannamei. Aquaculture 2005, 246, 139–149. [Google Scholar] [CrossRef]
- Gitterle, T.; Ødegård, J.; Gjerde, B.; Rye, M.; Salte, R. Genetic parameters and accuracy of selection for resistance to White Spot Syndrome Virus (WSSV) in Penaeus (Litopenaeus) vannamei using different statistical models. Aquaculture 2006, 251, 210–218. [Google Scholar] [CrossRef]
- Gitterle, T.; Gjerde, B.; Cock, J.; Salazar, M.; Rye, M.; Vidal, O.; Lozano, C.; Erazo, C.; Salte, R. Optimization of experimental infection protocols for the estimation of genetic parameters of resistance to White Spot Syndrome Virus (WSSV) in Penaeus (Litopenaeus) vannamei. Aquaculture 2006, 261, 501–509. [Google Scholar] [CrossRef]
- Hayes, B.J.; Gitterle, T.; Gopikrishna, G.; Gopal, C.; Krishna, G.; Jahageerdar, S.; Lozano, C.; Alavandi, S.; Paulpandi, S.; Ravichandran, P.; et al. Limited evidence for genetic variation for resistance to the white spot syndrome virus in Indian populations of Penaeus monodon. Aquac. Res. 2010, 41, e872–e877. [Google Scholar] [CrossRef]
- Huang, Y.-C.; Yin, Z.-X.; Ai, H.-S.; Huang, X.-D.; Li, S.-D.; Weng, S.-P.; He, J.-G. Characterization of WSSV resistance in selected families of Litopenaeus vannamei. Aquaculture 2011, 311, 54–60. [Google Scholar] [CrossRef]
- Cuéllar-Anjel, J.; White-Noble, B.; Schofield, P.; Chamorro, R.; Lightner, D.V. Report of significant WSSV-resistance in the Pacific white shrimp, Litopenaeus vannamei, from a Panamanian breeding program. Aquaculture 2012, 368–369, 36–39. [Google Scholar] [CrossRef]
- Gitterle, T.; Rye, M.; Salte, R.; Cock, J.; Johansen, H.; Lozano, C.; Arturo Suárez, J.; Gjerde, B. Genetic (co)variation in harvest body weight and survival in Penaeus (Litopenaeus) vannamei under standard commercial conditions. Aquaculture 2005, 243, 83–92. [Google Scholar] [CrossRef]
- Liu, G.; Zhu, S.; Liu, D.; Guo, X.; Ye, Z. Effects of stocking density of the white shrimp Litopenaeus vannamei (Boone) on immunities, antioxidant status, and resistance against Vibrio harveyi in a biofloc system. Fish Shellfish Immunol. 2017, 67, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Benchmarck Insights. Technologies shaping the future of shrimp production. Benchmark Insights, 11 November 2019. [Google Scholar]
- Zacarias, S.; Carboni, S.; Davie, A.; Little, D.C. Reproductive performance and offspring quality of non-ablated Pacific white shrimp (Litopenaeus vannamei) under intensive commercial scale conditions. Aquaculture 2019, 503, 460–466. [Google Scholar] [CrossRef]
- Treerattrakool, S.; Panyim, S.; Udomkit, A. Induction of Ovarian Maturation and Spawning in Penaeus monodon Broodstock by Double-Stranded RNA. Mar. Biotechnol. 2011, 13, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Treerattrakool, S.; Chartthai, C.; Phromma-in, N.; Panyim, S.; Udomkit, A. Silencing of gonad-inhibiting hormone gene expression in Penaeus monodon by feeding with GIH dsRNA-enriched Artemia. Aquaculture 2013, 404–405, 116–121. [Google Scholar] [CrossRef]
- Feijó, R.G.; Braga, A.L.; Lanes, C.F.C.; Figueiredo, M.A.; Romano, L.A.; Klosterhoff, M.C.; Nery, L.E.M.; Maggioni, R.; Wasielesky, W.; Marins, L.F. Silencing of Gonad-Inhibiting Hormone Transcripts in Litopenaeus vannamei Females by use of the RNA Interference Technology. Mar. Biotechnol. 2016, 18, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.J.; Bae, S.H.; Suzuki, T.; Niitsu, S.; Wilder, M.N. Transcriptional silencing of vitellogenesis-inhibiting hormone (VIH) subtype-I in the whiteleg shrimp, Litopenaeus vannamei. Aquaculture 2019, 506, 119–126. [Google Scholar] [CrossRef]






| System | Main Characteristics | Reference |
|---|---|---|
| 1. Biofloc technology (heterotrophic-based, also known as ‘pure BFT’) | Several scientific studies available High reliance on heterotrophic bacteria to control toxic N-compounds Application of a mature (biofloc-based) inoculum is often observed to speed up the microbial maturation process High C:N ratio (normally 15–20:1) and routine external carbon inputs (normally during the entire cycle) Levels of suspended solids (e.g., settling solids) normally varying from 5 up to more than 10 mL/L Clarifiers, protein skimmers, and/or settling chambers to control suspended solids are often used Low-intermediate water exchange rates to remove the sludge and solids Drop in pH and alkalinity is often observed Routine application of water probiotics, carbonate and bicarbonate sources and other minerals Biofloc particles with low lipid content | [3,4,9,73,74,75,76] |
| 2. Biofloc technology (chemoautotrophic based) | Little scientific information available High reliance on chemoautotrophic (nitrifying) bacteria to control N-compounds Application of a mature chemoautotrophic-based inoculum is often observed to speed up the microbial maturation process Intermediate C:N ratio (~10:1) with low external carbon inputs (normally up to the first 30–50 days) or even none Low levels of suspended solids (e.g., settling solids, normally up to 5 mL/L) Clarifiers, protein skimmers, and/or settling chambers to control suspended solids are often used Drop in pH and alkalinity is often observed Routine application of water probiotics and intensive control of alkalinity levels with routine application of carbonate and bicarbonate and other mineral sources. Low water exchange rates to remove the sludge and solids Biofloc particles with low lipid content | [4,17,74,77] |
| 3. Semi-biofloc (photoautotrophic-based, also known as ‘green-biofloc’) | Little scientific information available High reliance on microalgae to control N-compounds, resulting in low capacity of organic matter digestion by heterotrophic (degrading) bacteria Adoption of a mature inoculum is quite unusual Little control of the C:N ratio (normally < 10:1) with low or no external carbon inputs Low levels of suspended solids (e.g., settling solids normally up to 5 mL/L, but higher levels can be observed with algae blooms) Clarifiers, protein skimmers, and/or settling chambers to control suspended solids are quite unusual Partial/none mesh cover in ponds with some degree of pH fluctuation Routine application of water probiotics. However, less need for carbonate and bicarbonate sources to control alkalinity Higher water exchange rates to remove the sludge (mainly dead algae cells) and suspended solids Biofloc particles with high lipid content | [3,37,78] |
| 4. Semi-biofloc (mixed trophic conditions) | Little scientific information available A mix between algae and bacteria is promoted to control N-compounds, resulting in intermediate capacity of organic matter digestion by heterotrophic (degrading) bacteria Adoption of a mature inoculum is quite unusual Intermediate C:N ratio (10–15:1) with external carbon inputs, especially in the first days of production cycle Low levels of suspended solids (e.g., settling solids normally up to 5 mL/L) Clarifiers, protein skimmers, and/or settling chambers to control suspended solids are quite unusual Ponds are fully or partially mesh covered Large amount of water probiotics is routinely applied into ponds, helping to control algae bloom and pH fluctuations. Less need of carbonate and bicarbonate sources to control alkalinity Intermediate to high water exchange rates to remove the sludge and suspended solids Biofloc particles with intermediate lipid content | [3,66,67] |
| 5. Aquamimicry (without fish) | Little scientific information available A mix between algae and bacteria is promoted to control N-compounds, resulting in intermediate capacity of organic matter digestion by heterotrophic (degrading) bacteria Routine external fermented carbon inputs generate intermediate C:N ratio (10–15:1) Routine application of water probiotics, and carbonate and bicarbonate sources to control alkalinity Normally low levels of suspended solids (e.g., settling solids normally up to 5 mL/L, but higher levels can be observed with algae blooms) Clarifiers, protein skimmers, and/or settling chambers to control suspended solids are quite unusual Intermediate to high water exchange rates to remove the sludge and suspended solids Zooplankton blooms (e.g., insect larvae, copepods, rotifers, etc.) are expected | [70] |
| 6. Aquamimicry (integrated with fish) | Little scientific information available Water continuously circulates from shrimp ponds, to fish ponds and water treatment ponds with shrimp sludge been directed into fish ponds Other characteristics are the same as per ‘Aquamimicry without fish’. | [28] |
| 7. AquaScience® (integrated with tilapia) | Little scientific information is available A mix between algae and bacteria is promoted to control N-compounds. Shrimp sludge is drained (shrimp toilet) to fish and nitrification pounds. Decantation, heterotrophic bacteria, and microalgae are used to treat the water. After treatment, the water returns to shrimp pounds. Water exchange is minimal and is reused for consecutive production cycles. Relatively small and lined pounds are used for shrimp production (0.4 ha). | [71] |
| 8. Synbiotics | Little scientific information available A mix between algae and bacteria is promoted to control N-compounds. High capacity of organic matter digestion by heterotrophic (degrading) bacteria Routine addition of external fermented (aerobic, anaerobic or both) carbon inputs, with or without the addition of exogenous enzymes, normally representing an intermediate C:N ratio (10–15:1) Low levels of suspended solids are expected (e.g., settling solids normally up to 5 mL/L) Clarifiers, protein skimmers and/or settling chambers to control suspended solids are unusual Intermediate water exchange rates to remove the sludge and suspended solids Routine application of water probiotics, carbonate, and bicarbonate sources to control alkalinity (pH is normally stable) High bacteria loads, intense zooplankton bloom might not occur | [68,69] |
| System | Main Characteristics | Reference |
|---|---|---|
| 1. Flow-through | Little scientific information available Water exchange varying from 10 to more than 100%/day is promoted to control N-compounds Relatively low natural productivity (e.g., bacteria, phytoplankton, and zooplankton) Low inputs (e.g., bioremediators, carbon sources, etc.) compared to microbial-based systems pH fluctuations and low levels of solids | [28,80] |
| 2. RAS (recirculating aquaculture systems) | Little scientific information available Incorporates conventional RAS equipment and filtering devices (normally in indoor conditions, ‘boutique type’ farms, improving carrying capacity, e.g., >5 kg m−3) Relatively low natural productivity (e.g., bacteria, phytoplankton, and zooplankton) Routine application of carbonate and bicarbonate sources (use of water probiotics are quite unusual) Low water exchange rates | [80,81,82,85] |
| 3. Green-water RAS (photoautotrophic-based) | No scientific information available Incorporates RAS equipment and filtering devices (indoor conditions) Phytoplankton and chemoautotrophic bacteria dominance over heterotrophic bacteria (less light control). Low capacity of organic matter digestion by heterotrophic (degrading) bacteria Little control of the C:N ratio (normally < 10:1) with low or no external carbon inputs Higher water circulation rates to remove the sludge (mainly dead algae cells) and suspended solids (e.g., settling solids normally up to 5 mL/L, but higher levels can be observed with algae blooms) Routine application of carbonate and bicarbonate sources, and usual application of water probiotics Low water exchange rates | [86] |
| 4. BioRAS (heterotrophic based) | Little scientific information available Incorporates RAS equipment and filtering devices (indoor conditions) Heterotrophic and chemoautotrophic bacteria dominance over phytoplankton (higher light control, pH stable) Intermediate C:N ratio (~10:1) with low external carbon inputs (normally up to the first 30–50 days) Intermediate capacity of organic matter digestion by heterotrophic (degrading) bacteria Higher water circulation rates to remove the sludge and suspended solids (e.g., settling solids normally up to 5 mL/L) Routine application of bioremediators, carbonate and bicarbonate sources Low water exchange rates | [83,85,87] |
| Integrated Species | Shrimp Production System | Evaluated Aspect | Main Findings | Reference |
|---|---|---|---|---|
| Aquaponics | ||||
| Sarcocornia ambigua | BFT | Aquaponics vs. shrimp monoculture | N use was 25% more efficient, 2 kg of plants produced for each kg of shrimp | [107] |
| Ocimum basilicum | RAS | Water source and aquaponic system | Low-salinity groundwater resulted in greater shrimp and basil yields | [108] |
| S. ambigua | BFT | Different salinities | Optimal salinity between 16 and 24 g L−1 | [114] |
| Shrimp and Macroalgae | ||||
| Ulva lactuca | Recirculation system | Integration vs. shrimp monoculture | Integrated system maintained adequate water quality, improved growth for shrimp fed seaweed | [111] |
| U. prolifera | Minimum water exchange | Water exchange rate and algae density | 10% water exchange and 800 mg L−1 of stocked algae improved shrimp growth and survival | [115] |
| U. fasciata and U. ohnoi | BFT | Algae species and density | Best performance for U. ohnoi under 2 g L−1 | [110] |
| Shrimp and fish or shellfish | ||||
| O. niloticus | BFT | Fish stocking densities | Recovery of N and P and overall yield increased linearly | [116] |
| M. curema | BFT | Integration vs. shrimp monoculture | Increases in overall yield and P recovery | [117] |
| M. liza | BFT | Integration vs. shrimp monoculture | Lower TSS concentrations in integrated systems | [113] |
| M. curema | BFT | Fish stocking densities | Integration of mullet and shrimp increased biofloc system yield by 11.9% | [118] |
| O. niloticus | BFT | Heterotrophic and mature BFT systems | Higher fish and overall yields in heterotrophic BFT | [119] |
| Crassostrea gigas | Water exchange-based | Integration vs. shrimp monoculture | Improved shrimp growth performance and water quality by oyster presence | [120] |
| O. niloticus | BFT | Fish stocking densities | Increasing stocking densities affected fish physiology | [121] |
| Multitrophic | ||||
| O. niloticus and S. ambigua | BFT | Three-species integration vs. shrimp+fish | Total yield increased by 21.5%, reduction in water nitrate concentration | [112] |
| M. liza and U. fasciata | BFT | Three-species integration vs. shrimp+fish | Yield increase of 12.2%, improved N and P recovery, improved sea lettuce biochemical composition | [122] |
| Nutrient Requirements (%) | L. vannamei | ||
|---|---|---|---|
| RAS | Semi-Intensive | Intensive | |
| Crude protein | 38–44 | 33–42 | 40–46 |
| Crude lipid | 9–11 | 7 | 8 |
| Dig. energy (kJ/kg) | 15,820–16,292 | 14,033–15,380 | 15,079–15,874 |
| Amino acids (%) | |||
| Arg | 2.56–2.94 | 2.58–2.92 | 2.69–2.99 |
| His | 0.73–0.83 | 0.73–0.82 | 0.77–0.84 |
| Ile | 1.51–1.71 | 1.52–1.70 | 1.59–1.73 |
| Leu | 2.52–2.99 | 2.53–2.98 | 2.64–3.06 |
| Lys | 2.76–3.18 | 2.72–3.14 | 2.83–3.22 |
| Met | 0.97–1.11 | 0.98–1.11 | 1.01–1.13 |
| Phe | 1.74–1.97 | 1.76–1.96 | 1.83–2.00 |
| Thr | 1.31–1.56 | 1.31–1.54 | 1.37–1.58 |
| Trp | 0.34–0.39 | 0.34–0.39 | 0.36–0.39 |
| Val | 1.7–2.01 | 1.72–2.00 | 1.79–2.04 |
| Fatty acids (%) | |||
| Sum n-3 | 0.89 | 0.83 | 0.87 |
| Sum n-6 | 0.6 | 0.6 | 0.6 |
| EPA + DHA | 0.71–1.01 | 0.67–0.94 | 0.69–0.98 |
| Cholesterol | 667–834 | 521–727 | 540–752 |
| Phospholipids | 1.1–1.5 | 1–1.4 | 1.1–1.4 |
| Ingredient | APD (%) | Reference |
|---|---|---|
| Fishmeal | 83.7–91.6 | [150,151,153,164] |
| Krill meal | 80.5 | [151] |
| Meat and bone meal | 73.9–82.2 | [150,153] |
| Hydrolyzed feather meal | 63.9 | [151] |
| Poultry meal | 75.0–78.7 | [150,151,153] |
| Soybean meal | 89.0–96.9 | [150,151,153,154] |
| Soy protein isolate | 93.7–96.2 | [151,154] |
| Canola meal | 78.3 | [150] |
| Wheat (gross energy digestibility) | 87.0 | [155] |
| Supplier | Adisseo | Biomin | Alltech | Lallemand | Evonik | DSM | Cargill | BASF | DuPont (Danisco Animal Nutrition) | ADM | Kemin |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Prebiotic, immunostimulants and immunomodulators | Nutri®-Farm Stim S | Levabon® Aquagrow E | Bio-MOS®, Actigen® | Agrimos, Yang, M-glucan, Fibosel | BalanGut® LS | CitriStim® | AquastemTM | ||||
| Probiotic | Nutri®-Farm P/PW/L/FE | AquaStar® | Acid-Pak 4 way Lacto-Sacc | Bactocell | Ecobiol®, Fecinor®, Gutcare® | ||||||
| Phytobiotic | Sanacore® GM | Natustat | Xtract (Pancosma) | ||||||||
| Amino acids | Rhodimet® For Aqua | MetAMINO®, AQUAVI® Met-Met, Biolys®, ThreAMINO®, TrypAMINO® | Betafin® | Proplex®, L-lysine, L-threonine | |||||||
| Nucleotides | Nupro® | Laltide® | Rovimax, Rovimax NX Plus | ||||||||
| Enzymes and digestion enhancers | Aquagest® S, Aqualyso®, Lipogest® | Digestarom® | Aquate, Allzyme SSF® Allzyme VegPro | Phytase, xylanase and protease WX RONOZYME® WX, HiPhos RONOZYME® ProAct, PRoAct 360TM | Xylanase, phytase | AquatriaTM | |||||
| Vitamins | Aquavit® C stable | OVN, ROVIMIX® STAY-C®35 | Lutavit® A, Lutavit® B2, Lutavit® E, Vitamin A palmitate, propionate, acetate | VibrellTM C | |||||||
| Minerals | Selisseo® | Bioplex®, Sel-plex® | Copper-glycinate, Iron-glycinate, manganese-glycinate, Zinc-glycinate | B-traxim (Pancosma) | |||||||
| Acidifiers (organic acids) | Bacti-Nil® | Biotronic® | Acid-Balance AcidPak 4 Way | Amasil®, Lupro-Cid®, Lupro-Grain®, Lupro-Mix®NA, | DaaFIT® (Pancosma) | ||||||
| Antioxidants | Oxy-Nil® Aqua Zero, | Banox Antiox-Av Antiox-RC Nature-Ban Vitalix | Alkosel 2000, Melofeed | Carophyll Pink Rovimix E Stay C | ProvioxTM | Lucanthin® Pink | OxiviaTM C | ||||
| Attractants | Aquabite® | ||||||||||
| Binders | Nutribind® | AllBind |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emerenciano, M.G.C.; Rombenso, A.N.; Vieira, F.d.N.; Martins, M.A.; Coman, G.J.; Truong, H.H.; Noble, T.H.; Simon, C.J. Intensification of Penaeid Shrimp Culture: An Applied Review of Advances in Production Systems, Nutrition and Breeding. Animals 2022, 12, 236. https://doi.org/10.3390/ani12030236
Emerenciano MGC, Rombenso AN, Vieira FdN, Martins MA, Coman GJ, Truong HH, Noble TH, Simon CJ. Intensification of Penaeid Shrimp Culture: An Applied Review of Advances in Production Systems, Nutrition and Breeding. Animals. 2022; 12(3):236. https://doi.org/10.3390/ani12030236
Chicago/Turabian StyleEmerenciano, Maurício G. C., Artur N. Rombenso, Felipe d. N. Vieira, Mateus A. Martins, Greg J. Coman, Ha H. Truong, Tansyn H. Noble, and Cedric J. Simon. 2022. "Intensification of Penaeid Shrimp Culture: An Applied Review of Advances in Production Systems, Nutrition and Breeding" Animals 12, no. 3: 236. https://doi.org/10.3390/ani12030236