
Effects of Low Dietary Cation and Anion Difference on Blood Gas, Renal Electrolyte, and Acid Excretions in Goats in Tropical Conditions

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Animal Care
2.2. Ambient Condition, Behavior, Sample Collections, and Measurements
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saipin, N.; Semsirmboon, S.; Rungsiwiwut, R.; Thammacharoen, S. High ambient temperature directly decreases milk synthesis in the mammary gland in Saanen goats. J. Therm. Biol. 2020, 94, 102783. [Google Scholar] [CrossRef] [PubMed]
- Thammacharoen, S.; Chanpongsang, S.; Chaiyabutr, N.; Teedee, S.; Pornprapai, A.; Insam-ang, A.; Srisaard, C.; Channacoop, N. An analysis of herd-based lactation curve reveals the seasonal effect from dairy cows fed under high ambient temperature. Thai J. Vet. Med. 2020, 50, 169–178. [Google Scholar]
- Thammacharoen, S.; Komolvanich, S.; Chanpongsang, S.; Chaiyabutr, N. Respiratory Hypocapnia at Different Stages of Lactation during Long-term Exogenous Bovine Somatotropin in Crossbred Holstein Cattle in the Tropic. Thai J. Vet. Med. 2011, 41, 245–250. [Google Scholar]
- Semsirmboon, S.; Nguyen, D.K.D.; Chaiyabutr, N.; Poonyachoti, S.; Thammacharoen, S. Natural high ambient temperature-induced respiratory hypocapnia without activation of the hypothalamic-pituitary-adrenal axis in lactating goats. Vet. World 2022, in press. [Google Scholar] [CrossRef]
- Videla, R.; van Amstel, S. Urolithiasis. Vet. Clin. North Am. Food Anim. Pract. 2016, 32, 687–700. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.C.; Sherman, D.M. Urinary System. In Goat Medicine, 2nd ed.; Smith, M.C., Sherman, D.M., Eds.; Willey-Blackwell: Ames, IA, USA, 2009; pp. 537–570. [Google Scholar]
- Ewoldt, J.M.; Anderson, D.E.; Miesner, M.D.; Saville, W.J. Short- and Long-Term Outcome and Factors Predicting Survival After Surgical Tube Cystostomy for Treatment of Obstructive Urolithiasis in Small Ruminants. Vet. Surg. 2006, 35, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Oman, R.E.; Reppert, E.J.; Streeter, R.N.; Jones, M. Outcome and complications in goats treated by perineal urethrostomy for obstructive urolithiasis: 25 cases (2010–2017). J. Vet. Intern. Med. 2018, 33, 292–296. [Google Scholar] [CrossRef] [Green Version]
- Sickinger, M.; Windhorst, A. A systematic review on urolithiasis in small ruminants according to nutrition-dependent prevalence and outcome after surgery. Vet. World 2022, 15, 809–817. [Google Scholar] [CrossRef]
- Block, E. Manipulation of Dietary Cation-Anion Difference on Nutritionally Related Production Diseases, Productivity, and Metabolic Responses of Dairy Cows. J. Dairy Sci. 1994, 77, 1437–1450. [Google Scholar] [CrossRef]
- Stratton-Phelps, M.; House, J.K. Effect of a commercial anion dietary supplement on acid-base balance, urine volume, and urinary ion excretion in male goats fed oat or grass hay diets. Am. J. Vet. Res. 2004, 65, 1391–1397. [Google Scholar] [CrossRef]
- Jones, M.L.; Streeter, R.N.; Goad, C.L. Use of dietary cation anion difference for control of urolithiasis risk factors in goats. Am. J. Vet. Res. 2009, 70, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Chaiyabutr, N.; Chanpongsang, S.; Thammacharoen, S. Dietary cation and anion difference: Effects on milk production and body fluid distribution in lactating dairy goats under tropical conditions. Anim. Sci. J. 2017, 89, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Chanpongsang, S.; Chaiyabutr, N.; Thammacharoen, S. The effect of dietary ions difference on drinking and eating patterns in dairy goats under high ambient temperature. Asian-Australasian J. Anim. Sci. 2018, 32, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Mavangira, V.; Cornish, J.M.; Angelos, J.A. Effect of ammonium chloride supplementation on urine pH and urinary fractional excretion of electrolytes in goats. J. Am. Vet. Med. Assoc. 2010, 237, 1299–1304. [Google Scholar] [CrossRef]
- AOAC. Official Method of Analysis of Official Analytical Chemists, 20th ed.Association of Official Analytical Chemists, Inc.: Arlington, VA, USA, 2016. [Google Scholar]
- Stevens, J.B.; Anderson, K.L.; Correa, M.T.; Stewart, T.; Braselton, W.E., Jr. Hematologic, Blood Gas, Blood Chemistry, and Serum Mineral Values for a Sample of Clinically Healthy Adult Goats. Vet. Clin. Pathol. 1994, 23, 19–24. [Google Scholar] [CrossRef]
- Chan, J. The rapid determination of urinary titratable acid and ammonium and evaluation of freezing as a method of preservation. Clin. Biochem. 1972, 5, 94–98. [Google Scholar] [CrossRef]
- Suwanapaporn, P.; Chaiyabutr, N.; Thammacharoen, S. A low degree of high ambient temperature decreased food intake and activated median preoptic and arcuate nuclei. Physiol. Behav. 2017, 181, 16–22. [Google Scholar] [CrossRef]
- Gomez, D.E.; Bedford, S.; Darby, S.; Palmisano, M.; Mackay, R.J.; Renaud, D.L. Acid-base disorders in sick goats and their association with mortality: A simplified strong ion difference approach. J. Vet. Intern. Med. 2020, 34, 2776–2786. [Google Scholar] [CrossRef]
- Scott, D.; Whitelaw, F.G.; Kay, M. Renal excretion of acid in calves fed either roughage or concentrate diets. Exp. Physiol. 1971, 56, 18–32. [Google Scholar] [CrossRef] [Green Version]
- Mahnensmith, R.; Thier, S.O.; Cooke, C.; Broadus, A.; DeFronzo, R.A. Effect of acute metabolic acidemia on renal electrolyte transport in man. Metabolism 1979, 28, 831–842. [Google Scholar] [CrossRef]
- Augustinsson, O.; Johansson, K. Ammonium chloride induced acidosis and aldosterone secretion in the goat. Acta Physiol. Scand. 1986, 128, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Constable, P.D.; Gelfert, C.-C.; Fürll, M.; Staufenbiel, R.; Stämpfli, H.R. Application of strong ion difference theory to urine and the relationship between urine pH and net acid excretion in cattle. Am. J. Vet. Res. 2009, 70, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Nan, X.; Zhao, P.; Liu, W.; Drackley, J.K.; Liu, S.; Zhang, K.; Bu, D. Effects of low dietary cation-anion difference induced by ruminal ammonium chloride infusion on performance, serum, and urine metabolites of lactating dairy cows. Asian-Australasian J. Anim. Sci. 2018, 31, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Weiner, I.D.; Verlander, J.W. Role of NH3 and NH4+ transporters in renal acid-base transport. Am. J. Physiol. Physiol. 2011, 300, F11–F23. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Chemical Compositions, % DM | Roughage | Control Feed | L-DCAD Feed |
|---|---|---|---|
| Moisture | 7.32 | 8.74 | 8.70 |
| Dry matter | 92.68 | 91.26 | 91.30 |
| Crude protein | 3.87 | 14.67 | 17.44 |
| Fat | 0.95 | 3.57 | 3.83 |
| Ash | 7.15 | 6.50 | 5.51 |
| Calcium | 0.73 | 1.05 | 0.92 |
| Phosphate | 0.12 | 0.50 | 0.37 |
| Sodium | 0.32 | 0.03 | 0.05 |
| Potassium | 1.42 | 1.28 | 1.28 |
| Sulfur | 0.19 | 0.19 | 0.11 |
| Chloride | 0.30 | 0.54 | 1.43 |
| Neutral detergent fiber | 73.12 | ||
| Acid detergent fiber | 44.84 | ||
| DCAD | 30.00 | 7.04 | −12.16 |
| Morning | Afternoon | p-Value | |
|---|---|---|---|
| Ambient condition | |||
| Ta | 25.13 ± 0.3 | 31.00 ± 0.67 | <0.01 |
| RH | 89.25 ± 2.77 | 74.31 ± 2.98 | <0.01 |
| THI | 76.14 ± 0.59 | 83.63 ± 0.89 | <0.01 |
| Biological parameters | |||
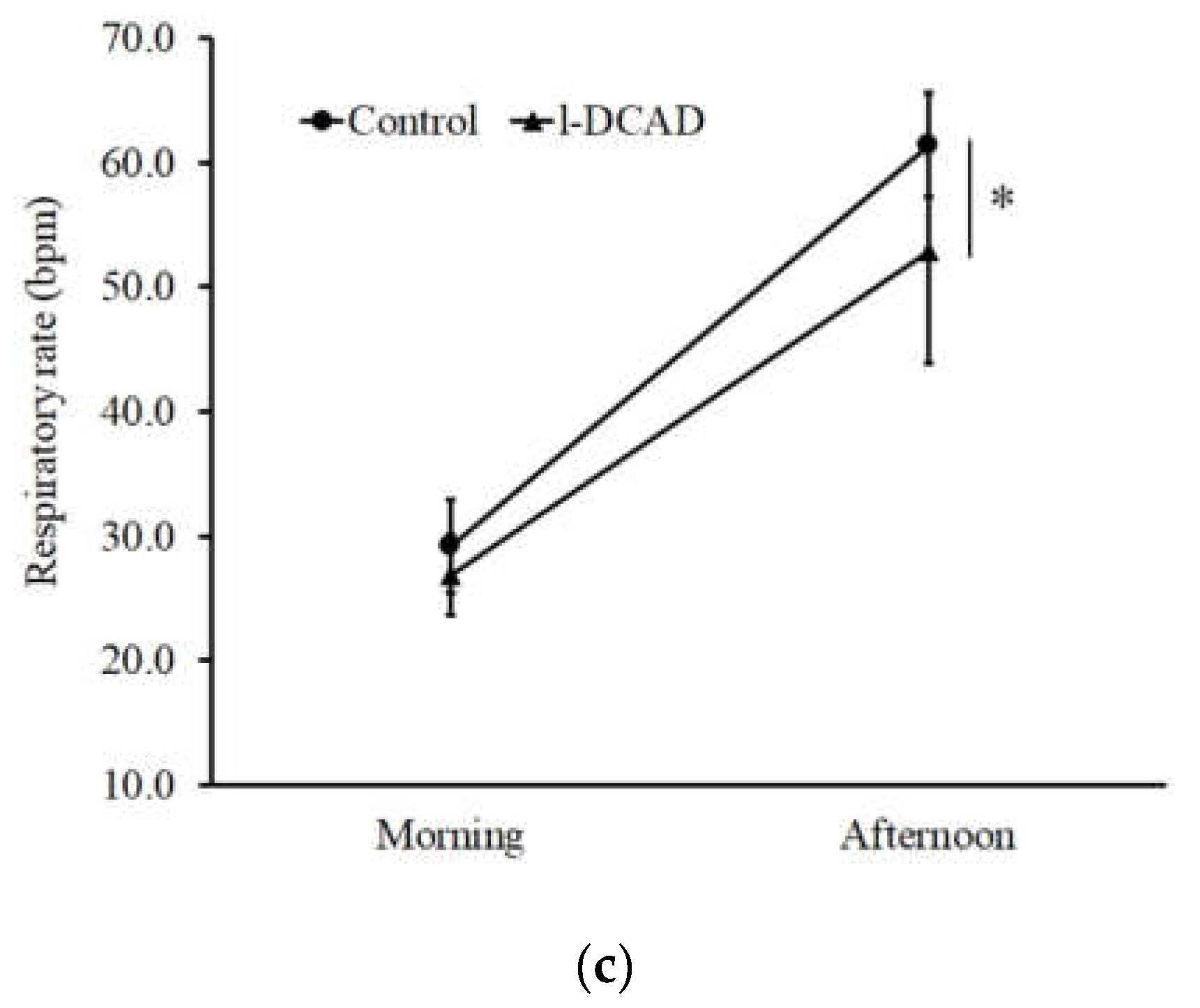
| Respiratory rate (bpm) | 30 ± 3 | 53 ± 4 | <0.01 |
| Rectal temperature (°C) | 38.5 ± 0.1 | 39.5 ± 0.1 | <0.01 |
| Hct (%) | 22.3 ± 0.6 | 22.0 ± 0.6 | 0.60 |
| Glucose (mg/dL) | 55.6 ± 0.8 | 58.8 ± 1.1 | 0.05 |
| BUN (mg/dL) | 19.3 ± 1.0 | 18.6 ± 1.2 | 0.54 |
| Na (mmol/L) | 148.5 ± 1.4 | 150.3 ± 1.4 | 0.38 |
| K (mmol/L) | 3.76 ± 0.10 | 3.82 ± 0.14 | 0.72 |
| Cl (mmol/L) | 103.9 ± 0.7 | 104.8 ± 0.5 | 0.30 |
| Ca (mmol/L) | 1.89 ± 0.05 | 1.95 ± 0.05 | 0.26 |
| Cortisol (ng/mL) | 42.76 ± 9.61 | 37.13 ± 9.18 | 0.33 |
| pH | 7.43 ± 0.01 | 7.44 ± 0.01 | 0.28 |
| PCO2 (mmHg) | 40.47 ± 0.57 | 38.76 ± 0.74 | 0.057 |
| HCO3 (mmol/L) | 26.80 ± 0.68 | 26.28 ± 0.53 | 0.34 |
| Tot CO2 (mmol/L) | 27.92 ± 0.68 | 27.33 ± 0.53 | 0.27 |
| Base excess (mmol/L) | 2.33 ± 0.83 | 2.17 ± 0.56 | 0.80 |
| Anion gap (mmol/L) | 21.5 ± 1.7 | 23.1 ± 1.5 | 0.54 |
| Morning | Afternoon | SEM | Effect (p Value) | ||||
|---|---|---|---|---|---|---|---|
| Control | l-DCAD | Control | l-DCAD | Time | l-DCAD | ||
| Blood chemistries, electrolytes, and hormones | |||||||
| Glucose (mg/dL) | 53.5 | 53.2 | 60.8 | 58.7 | 7.94 | <0.01 | 0.42 |
| BUN (mg/dL) | 20.8 | 19.0 | 20.7 | 20.8 | 1.98 | 0.18 | 0.77 |
| Na (mmol/L) | 150.9 | 149.3 | 150.1 | 153.5 | 18.12 | 0.32 | 0.72 |
| K (mmol/L) | 3.72 | 4.08 | 3.66 | 4.04 | 0.13 | 0.74 | 0.12 |
| Cl (mmol/L) | 102.0 | 107.8 | 101.2 | 109.8 | 4.54 | 0.52 | <0.01 |
| Ca (mmol/L) | 1.76 | 1.75 | 1.93 | 1.89 | 0.01 | <0.01 | 0.64 |
| Cortisol (ng/mL) | 56.91 | 86.30 | 64.92 | 52.56 | 5100 | 0.77 | 0.66 |
| Blood gas parameters | |||||||
| pH | 7.42 | 7.38 | 7.46 | 7.38 | 0.0004 | 0.03 | 0.02 |
| PCO2 (mmHg) | 40.58 | 37.93 | 39.12 | 37.02 | 1.24 | 0.03 | 0.02 |
| HCO3 (mmol/L) | 26.57 | 22.27 | 27.70 | 22.10 | 2.05 | 0.43 | <0.01 |
| Tot CO2 (mmol/L) | 27.83 | 23.33 | 28.83 | 23.33 | 1.90 | 0.40 | <0.01 |
| Base excess (mmol/L) | 2.17 | −3.17 | 3.67 | −2.83 | 2.54 | 0.19 | <0.01 |
| Anion gap (mmol/L) | 26.0 | 23.3 | 24.9 | 25.6 | 17.87 | 0.74 | 0.76 |
| Control | l-DCAD | p-Value | |
|---|---|---|---|
| Endogenous creatinine clearance | |||
| Plasma creatinine (mg/dL) | 0.60 ± 0.05 | 0.53 ± 0.06 | 0.40 |
| Urine creatinine (mg/dL) | 50.88 ± 15.47 | 53.07 ± 19.57 | 0.93 |
| Urine volume (8 h, mL/min) | 1.65 ± 0.31 | 2.85 ± 1.01 | 0.28 |
| Creatinine clearance (mL/min) | 113.03 ± 23.95 | 159.40 ± 20.29 | 0.17 |
| Fractional excretion of electrolytes, % | |||
| Na | 1.05 ± 0.05 | 0.42 ± 0.42 | 0.053 |
| K | 40.91 ± 3.55 | 23.31 ± 3.17 | <0.01 |
| Cl | 0.21 ± 0.04 | 1.38 ± 0.14 | <0.01 |
| Ca | 0.89 ± 0.25 | 2.78 ± 0.45 | <0.01 |
| Acid concentration (mmol/L) | |||
| Net acid | −159.97 ± 28.53 | −5.33 ± 14.87 | <0.01 |
| Titratable acid | −159.97 ± 28.53 | −18.00 ± 8.94 | <0.01 |
| Ammonium | 0.00 ±0.00 | 12.67 ± 7.56 | 0.12 |
| Urine pH | 8.21 ± 0.09 | 6.82 ± 0.44 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Do Nguyen, D.K.; Semsirmboon, S.; Chaiyabutr, N.; Thammacharoen, S. Effects of Low Dietary Cation and Anion Difference on Blood Gas, Renal Electrolyte, and Acid Excretions in Goats in Tropical Conditions. Animals 2022, 12, 3444. https://doi.org/10.3390/ani12233444
Do Nguyen DK, Semsirmboon S, Chaiyabutr N, Thammacharoen S. Effects of Low Dietary Cation and Anion Difference on Blood Gas, Renal Electrolyte, and Acid Excretions in Goats in Tropical Conditions. Animals. 2022; 12(23):3444. https://doi.org/10.3390/ani12233444
Chicago/Turabian StyleDo Nguyen, Dang Khoa, Sapon Semsirmboon, Narongsak Chaiyabutr, and Sumpun Thammacharoen. 2022. "Effects of Low Dietary Cation and Anion Difference on Blood Gas, Renal Electrolyte, and Acid Excretions in Goats in Tropical Conditions" Animals 12, no. 23: 3444. https://doi.org/10.3390/ani12233444