

Ecomorphology and Morphological Disparity of Caquetaia Kraussii (Perciformes: Cichlidae) in Colombia
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
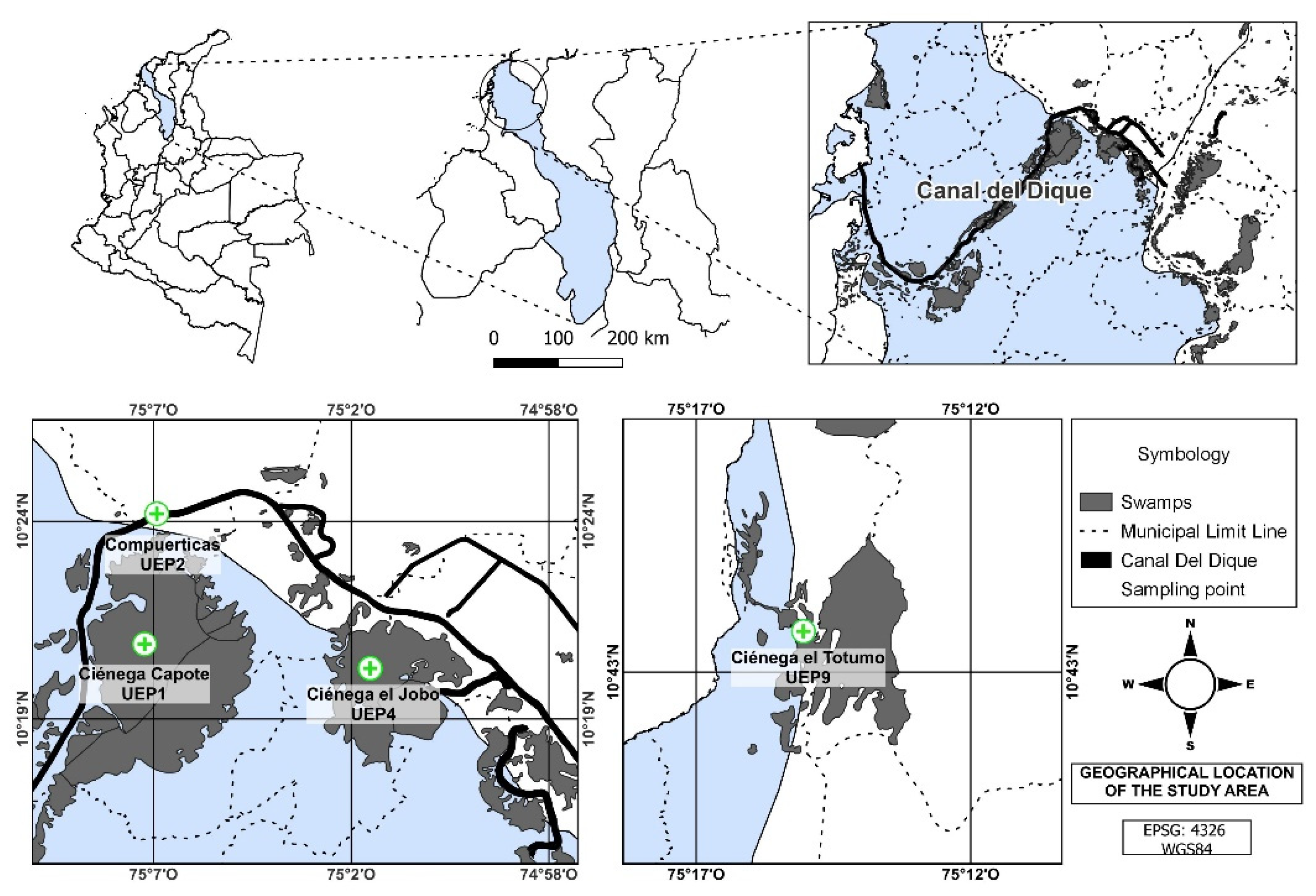
2.1. Field Work
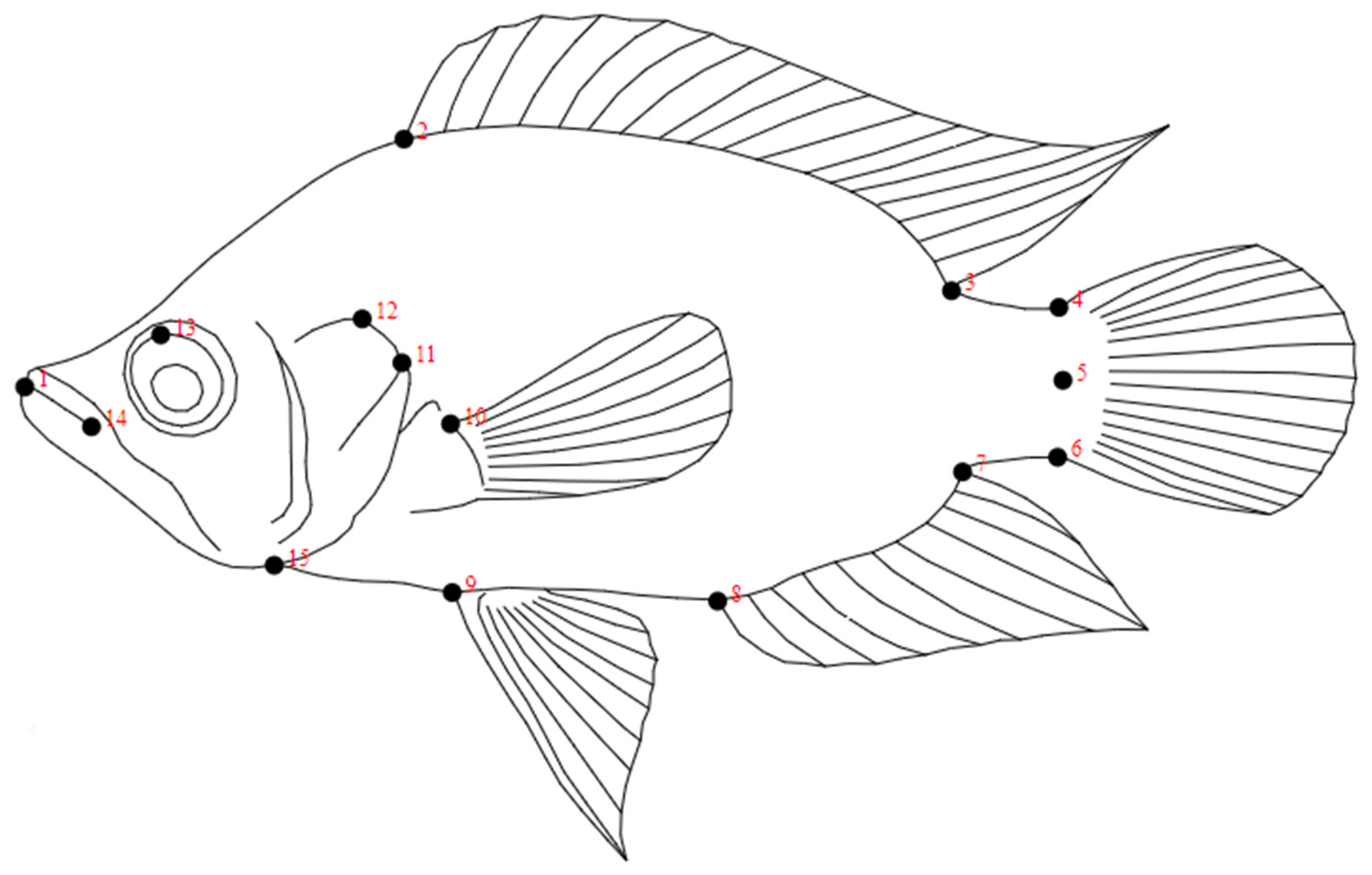
2.2. Geometric Morphometrics
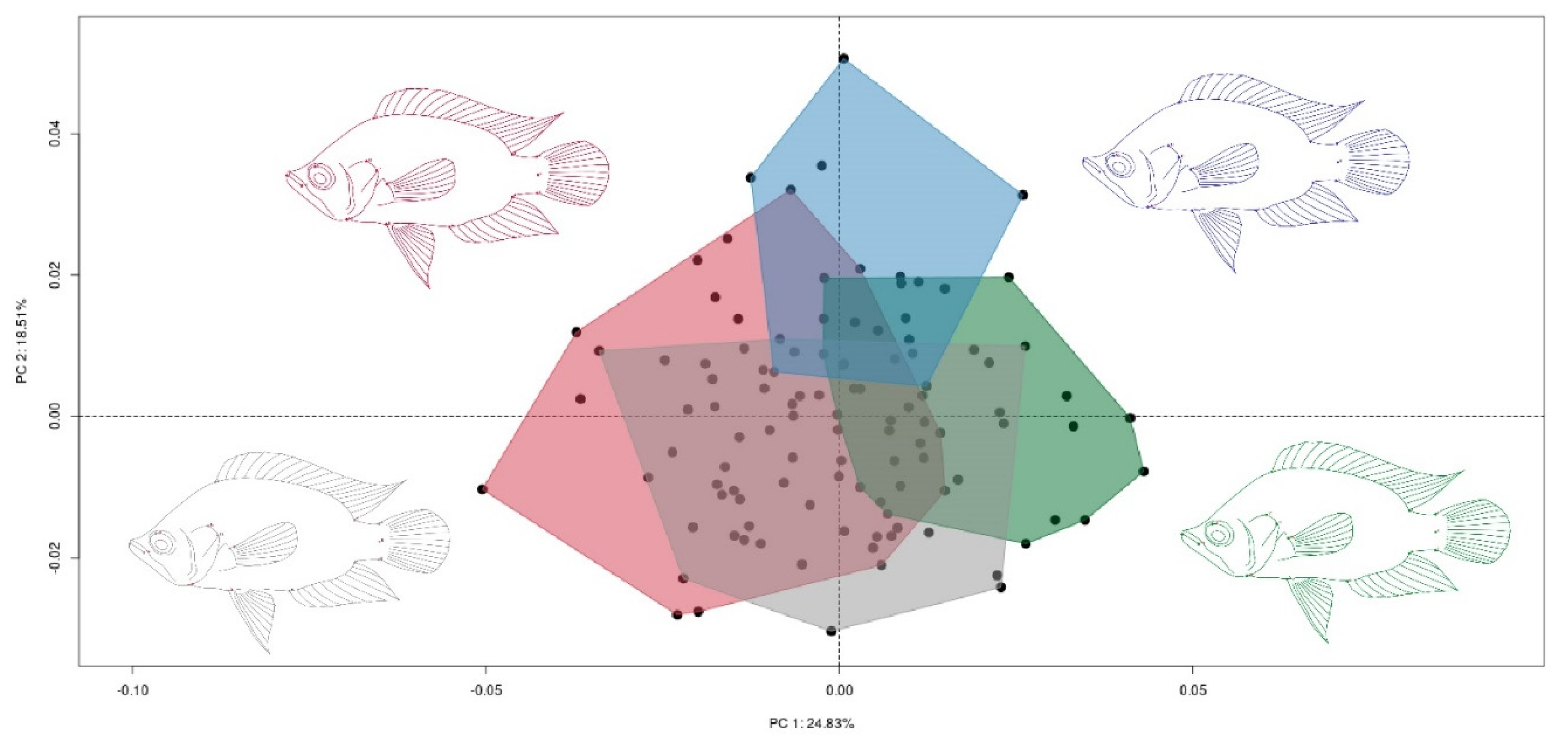
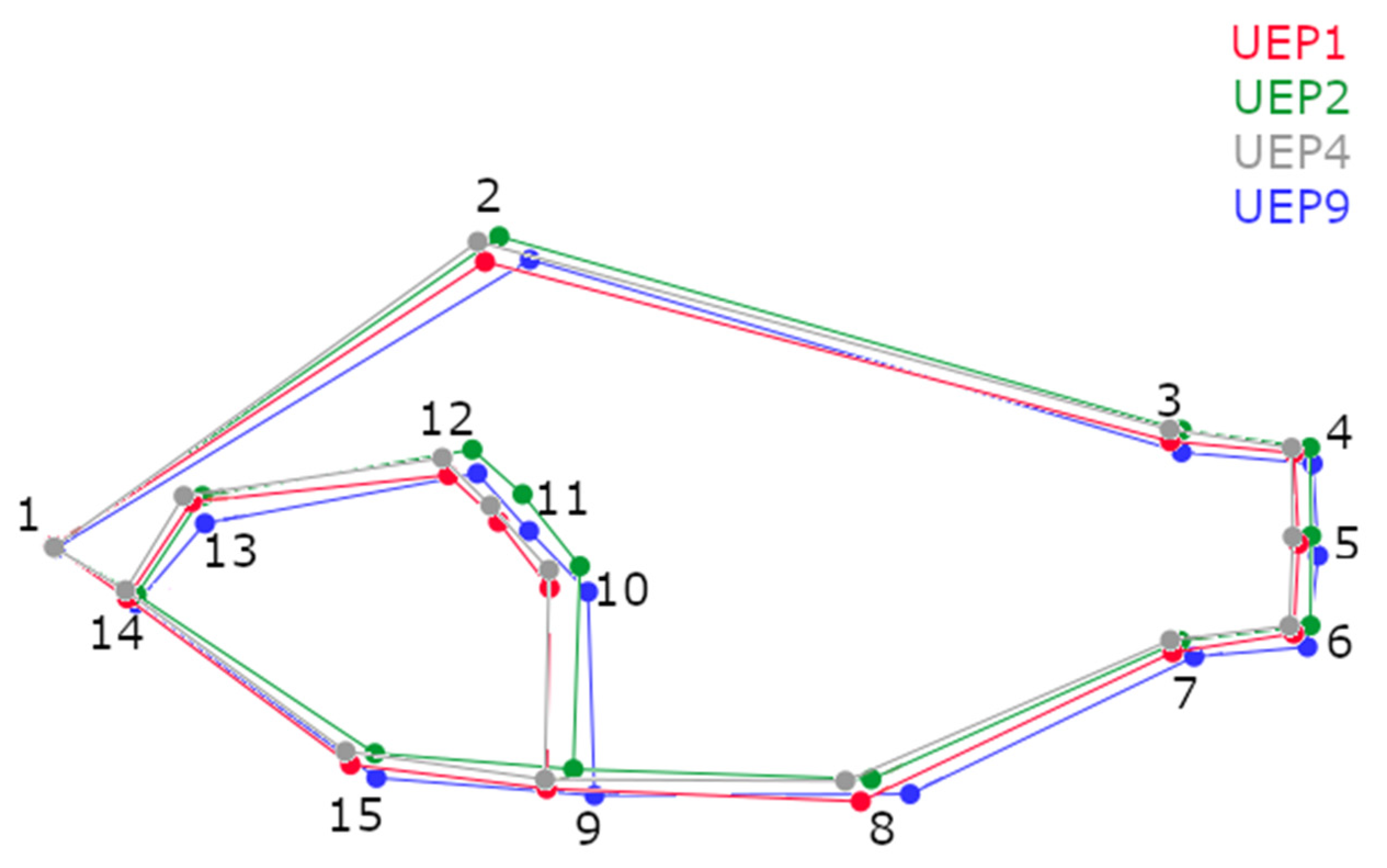
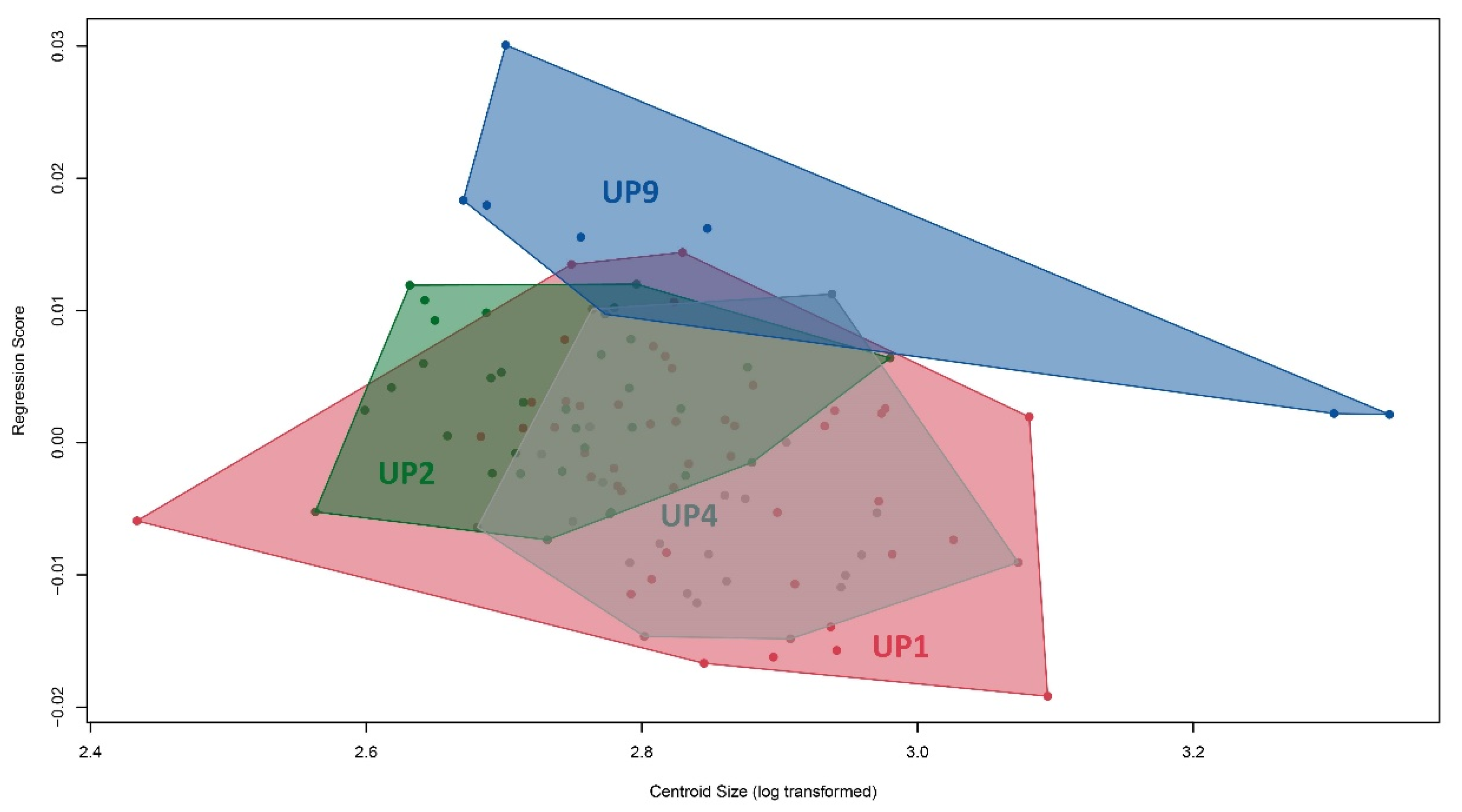
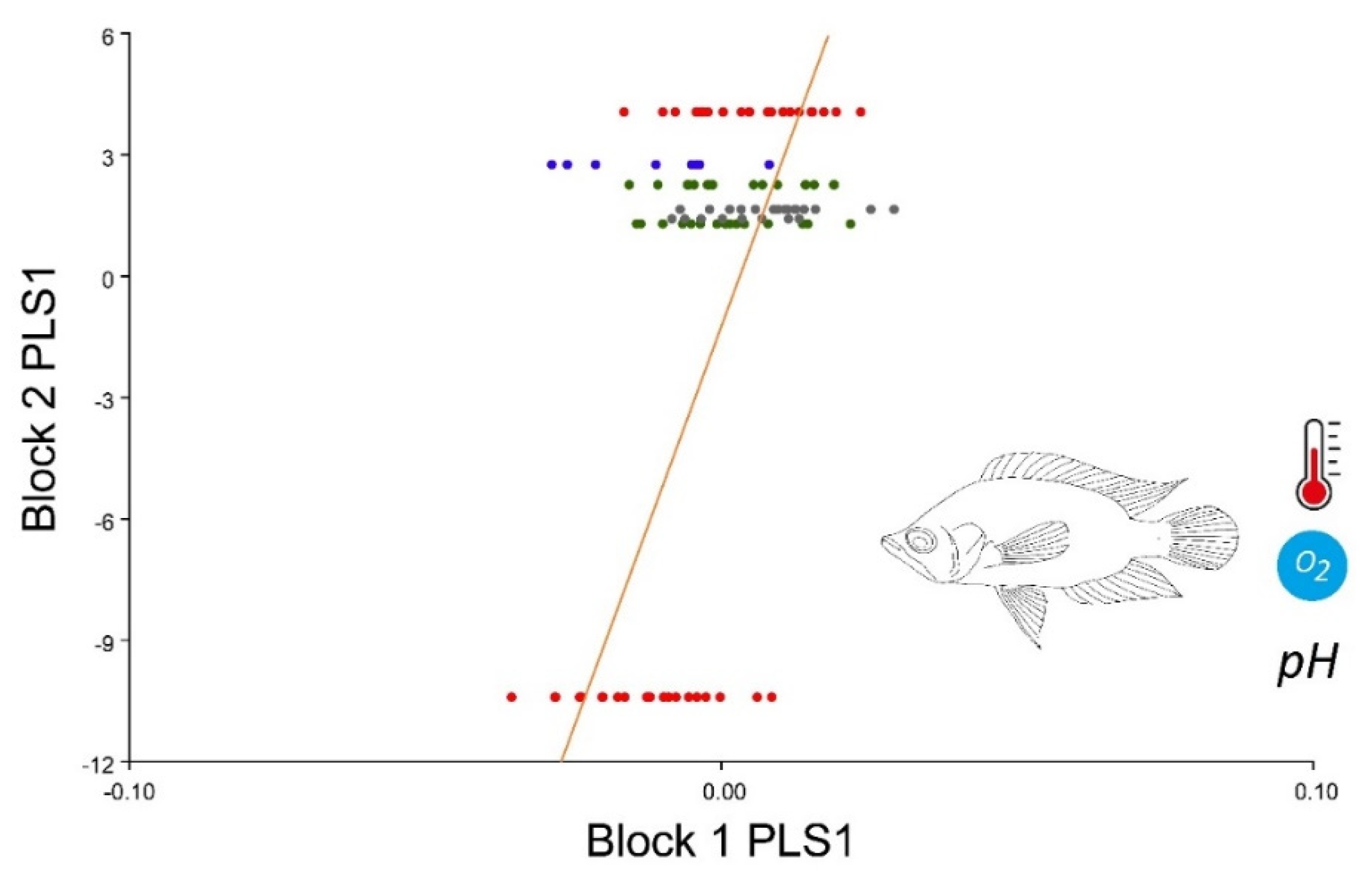
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adams, D.C.; Rohlf, F.J.; Slice, D.E. Geometric morphometrics: Ten years of progress following the ‘revolution’. Ital. J. Zool. 2004, 71, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Miner, B.G.; Sultan, S.E.; Morgan, S.G.; Padilla, D.K.; Relyea, R.A. Ecological consequences of phenotypic plasticity. Trends Ecol. Evol. 2005, 20, 685–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stillwell, R.C.; Davidowitz, G. A developmental perspective on the evolution of sexual size dimorphism of a moth. Proc. R. Soc. B Biol. Sci. 2010, 277, 2069–2074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kingsolver, J.G.; Huey, R.B. Introduction: The evolution of morphology, performance, and fitness. Integr. Comp. Biol. 2003, 43, 361–366. [Google Scholar] [CrossRef]
- Cadrin, S.X. Advances in morphometric identification of fishery stocks. Rev. Fish Biol. Fish. 2000, 10, 91–112. [Google Scholar] [CrossRef]
- Kaouèche, M.; Bahri-Sfar, L.; Hammami, I.; Hassine, O.K.B. Morphometric variations in white seabream Diplodus sargus (Linneus, 1758) populations along the Tunisian coast. Oceanologia 2017, 59, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, G. Evolutionary aspects of fish distribution: Genetic variability and adaptation. J. Fish Biol. 1993, 43, 53–73. [Google Scholar] [CrossRef]
- Turan, C.; Oral, M.; Öztürk, B.; Düzgüneş, E. Morphometric and meristic variation between stocks of Bluefish (Pomatomus saltatrix) in the Black, Marmara, Aegean and northeastern Mediterranean Seas. Fish. Res. 2006, 79, 139–147. [Google Scholar] [CrossRef]
- HammamI, I.; BaHrI-Sfar, L.; Kaoueche, M.; Grenouillet, G.; Lek, S.; Kara, M.; Hassine, O.B. Morphological characterization of striped seabream (Lithognathus mormyrus, Sparidae) in some Mediterranean lagoons. Cybium 2013, 37, 127–139. [Google Scholar]
- Motta, P.J.; Norton, S.F.; Luczkovich, J.J. Perspectives on the ecomorphology of bony fishes. Environ. Biol. Fishes 1995, 44, 11–20. [Google Scholar] [CrossRef]
- Foote, M. The evolution of morphological diversity. Annu. Rev. Ecol. Syst. 1997, 28, 129–152. [Google Scholar] [CrossRef]
- Koehl, M. When does morphology matter? Annu. Rev. Ecol. Syst. 1996, 27, 501–542. [Google Scholar] [CrossRef] [Green Version]
- Betz, O. Ecomorphology: Integration of form, function, and ecology in the analysis of morphological structures. Mitt. Dtsch. Ges. Allg. Angew. Entomol. 2006, 15, 409–416. [Google Scholar]
- Nuñez-Vallecillo, M.; Rivera, A.; Górski, K.; Brante, A.; Benítez, H.A. Ecomorphological analyses reveal impact of land-based stressors on stock structure of two commercially important fish species (Lutjanus synagris and Haemulon plumierii) in the Caribbean. Fish. Res. 2021, 234, 105812. [Google Scholar] [CrossRef]
- Helfman, G.; Collette, B.B.; Facey, D.E.; Bowen, B.W. The Diversity of Fishes: Biology, Evolution, and Ecology; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Hoagstrom, C.W.; Berry, C.R. Morphological diversity among fishes in a Great Plains river drainage. Hydrobiologia 2008, 596, 367–386. [Google Scholar] [CrossRef]
- Junqueira, N.T.; Leal, C.G.; Alves, C.B.M.; Pompeu, P.S. Morphological diversity of fish along the rio das Velhas, Minas Gerais, Brazil. Neotrop. Ichthyol. 2012, 10, 417–424. [Google Scholar] [CrossRef] [Green Version]
- Wainwright, P.C.; Bellwood, D.R.; Westneat, M.W. Ecomorphology of locomotion in labrid fishes. Environ. Biol. Fishes 2002, 65, 47–62. [Google Scholar] [CrossRef]
- Price, S.A.; Holzman, R.; Near, T.J.; Wainwright, P.C. Coral reefs promote the evolution of morphological diversity and ecological novelty in labrid fishes. Ecol. Lett. 2011, 14, 462–469. [Google Scholar] [CrossRef]
- McGee, M.D.; Wainwright, P.C. Convergent evolution as a generator of phenotypic diversity in Threespine Stickleback. Evolution 2013, 67, 1204–1208. [Google Scholar] [CrossRef]
- Bookstein, F.L. Morphometric Tools for Landmark Data: Geometry and Biology; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Adams, D.C.; Rohlf, F.J.; Slice, D.E. A field comes of age: Geometric morphometrics in the 21st century. Hystrix-Ital. J. Mammal. 2013, 24, 7–14. [Google Scholar] [CrossRef]
- Adams, D.C.; Rohlf, F.J. Ecological character displacement in Plethodon: Biomechanical differences found from a geometric morphometric study. Proc. Natl. Acad. Sci. USA 2000, 97, 4106–4111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bookstein, F.L. Size and shape spaces for landmark data in two dimensions. Stat. Sci. 1986, 1, 181–222. [Google Scholar] [CrossRef]
- Bookstein, F.L. Biometrics, biomathematics and the morphometric synthesis. Bull. Math. Biol. 1996, 58, 313–365. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.P.; Monteiro, L.R. Distances and directions in multidimensional shape spaces: Implications for morphometric applications. Syst. Biol. 2005, 54, 678–688. [Google Scholar] [CrossRef] [Green Version]
- Rohlf, F.J.; Slice, D. Extensions of the Procustes methods for the optimal superimposition of landmarks. Syst. Zool. 1990, 39, 40–59. [Google Scholar] [CrossRef] [Green Version]
- Toro-Ibacache, M.V.; Soto, G.M.; Galdames, I.S. Geometric Morphometry and the Biologic Shapes Study: From the Descriptive Morphology to the Quantitative Morphology. Int. J. Morphol. 2010, 28, 977–990. [Google Scholar]
- Benítez, H.A.; Püschel, T.A. Modelando la Varianza de la Forma: Morfometría Geométrica Aplicaciones en Biología Evolutiva. Int. J. Morphol. 2014, 32, 998–1008. [Google Scholar] [CrossRef] [Green Version]
- Villalobos-Leiva, A.; Ha, B. Morfometría geométrica y sus nuevas aplicaciones en ecología y biología evolutiva. Parte 2. Int. J. Morphol. 2020, 38, 1818–1836. [Google Scholar] [CrossRef]
- Cadrin, S.X. Stock identification of marine populations. In Morphometrics for Nonmorphometricians; Springer: Berlin/Heidelberg, Germany, 2010; pp. 219–232. [Google Scholar]
- Jaramillo, N. Morfometría geométrica: Principios teóricos y métodos de empleo. In Fronteras de investigación en enfermedades infecciosas. Modelo Enfermedad de Chagas; Universidad de Antioquia: Antioquia, Colombia, 2011; pp. 69–87. [Google Scholar]
- Alarcón-Durán, I.; Castillo-Rivera, M.A.; Figueroa-Lucero, G.; Arroyo-Cabrales, J.; Barriga-Sosa, I.D.L.Á. Morphological diversity in 6 populations of the Silverside Chirostoma humboldtianum. Rev. Mex. Biodivers. 2017, 88, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Narváez, J.C.; Acero, A.; Blanco, J. Variación morfométrica en poblaciones naturalizadas y domesticadas de la tilapia del Nilo Oreochromis niloticus (Teleostei: Cichlidae) en el norte de Colombia. Rev. Acad. Colomb. Cienc 2005, 29, 383–394. [Google Scholar]
- Olaya-Nieto, C.; Martínez-González, Á.; Díaz-Sánchez, D.; Pérez-Doria, W.; Segura-Guevara, F.; Tordecilla-Petro, G. Relación longitud-peso multianual de la Mojarra amarilla (Caquetaia kraussii Steindachner, 1878) en la Ciénaga de Ayapel, Sistema río San Jorge. Neiva: V Congreso Colombiano de Acuicultura. Rev. Entornos 2011, 186. [Google Scholar]
- Vanega, H.D.G. Algunos aspectos biológicos y pesqueros de Caquetaia kraussii (Steindachner, 1878) en la cuenca media y baja del río Atrato, Chocó. Rev. Biodivers. Neotrop. 2017, 7, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Solano-Peña, D.; Segura-Guevara, F.; Olaya-Nieto, C. Crecimiento y reproducción de la mojarra amarilla (Caquetaia kraussii Steindachner, 1878) en el embalse de Urrá, Colombia. Rev. MVZ Córdoba 2013, 18, 3525–3533. [Google Scholar] [CrossRef] [Green Version]
- Olaya-Nieto, C.W.; Ubarnes-Coronado, G.M.; Ensuncho-Morales, J.E. Crecimiento y mortalidad de mojarra amarilla Caquetaia kraussii en la ciénaga Grande de Lorica, Colombia. Rev. Logos Cienc. Tecnol. 2014, 5, 202–212. [Google Scholar] [CrossRef]
- Hernandez, J.; Villalobos-Leiva, A.; Bermúdez, A.; Ahumada-Cabarcas, D.; Suazo, M.J.; Benítez, H.A. An Overview of Interlocation Sexual Shape Dimorphism in Caquetaia kraussi (Perciformes: Cichlidae) A Geometric Morphometric Approach. Fishes 2022, 7, 146. [Google Scholar] [CrossRef]
- Lagares, G.A.A.; Martínez, L.D.P. Caracterización socioeconómica de la subregión del Canal del Dique. Econ. Y Soc. 2014, 37–61. Available online: https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwj-6tSPueT7AhVMjqQKHae_B5AQFnoECAwQAQ&url=https%3A%2F%2Ffondodeaguacartagena.org%2Fwp-content%2Fuploads%2F2017%2F10%2Fpub-002-ahumada-y-penso-2014.pdf&usg=AOvVaw10LRJVoWmsPHoEDfaeT2Ir (accessed on 5 October 2022).
- Aguilera-Díaz, M.M. El Canal del Dique y su subregión: Una economía basada en la riqueza hídrica. Doc. Trab. Sobre Econ. Reg. Urbana 2006. [Google Scholar] [CrossRef] [Green Version]
- Maldonado-Ocampo, J.; Ortega-Lara, A.; Usma-O, J.; Galvis-V, G.; Villa-Navarro, F.; Vásquez-G, L.; Prada-Pedreros, S.; Ardila-Rodríguez, C. Peces de los Andes de Colombia; Instituto de Investigación de Recursos Biológicos “Alexander Von Humboldt”: Bogotá, Colombia, 2005. [Google Scholar]
- Lasso, C.; Agudelo-Córdoba, E.; Jiménez-Segura, L.; Ramírez-Gil, H.; Morales-Betancourt, M.; Ajiaco-Martínez, R.; Paula-Gutiérrez, F.; Usma-Oviedo, J.; Muñoz, S.; Sanabria, A.I. Catálogo de los recursos pesqueros continentales de Colombia. Serie Editorial Recursos Hidrobiológicos y Pesqueros Amazonian migratory catfish Brachyplatystoma rousseauxii in the Iquitos Region, Peru. J. Fish Biol. 2011, 75, 2527–2551. [Google Scholar]
- Pinilla, G.A.; Duarte, J. La Importancia Ecológica de Las Ciénagas del Canal del Dique y la Detreminación de su Estado Limnológico; Departamento de Biología, Universidad Nacional de Colombia: Bogotá, Colombia, 2006. [Google Scholar]
- Madera, A.S.B.; Porto, J.L.G. Ecosistema Estuarino de la Ciénaga del Totumo; Corporación Universitaria de la Costa CUC: Barranquilla, Colombia, 2010. [Google Scholar]
- Rohlf, F.J.J.H. The tps series of software. Hystrix 2015, 26, 9–12. [Google Scholar]
- Rohlf, F.J. TPSdig; v. 2.17.; State University at Stony Brook: Stony Brook, NY, USA, 2013. [Google Scholar]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.P.; Barluenga, M.; Meyer, A. Shape analysis of symmetric structures: Quantifying variation among individuals and asymmetry. Evolution 2002, 56, 1909–1920. [Google Scholar] [CrossRef] [Green Version]
- González, R.; Tobón, A.B. Determinación de dimorfismo sexual usando técnicas morfométricas en Rachycentron canadum (Perciformes: Rachycentridae) cultivados en cautiverio. Boletín Investig. Mar. Costeras 2021, 50, 79–90. [Google Scholar] [CrossRef]
- Fruciano, C. Measurment error in geometric morphometrics. Dev. Genes Evol. 2016, 226, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, L.R. Multivariate regression models and geometric morphometrics: The search for causal factors in the analysis of shape. Syst. Biol. 1999, 48, 192–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohlf, F.J.; Corti, M. Use of two-block partial least-squares to study covariation in shape. Syst. Biol. 2000, 49, 740–753. [Google Scholar] [CrossRef] [Green Version]
- Bohórquez-Herrera, J.; Cruz-Escalona, V.H.; Adams, D.C.; Peterson, M.S. Feeding ecomorphology of seven demersal marine fish species in the Mexican Pacific Ocean. Environ. Biol. Fishes 2015, 98, 1459–1473. [Google Scholar] [CrossRef]
- Adams, D.C.; Otárola-Castillo, E. Geomorph: An R package for the collection and analysis of geometric morphometric shape data. Methods Ecol. Evol. 2013, 4, 393–399. [Google Scholar] [CrossRef]
- Baken, E.K.; Collyer, M.L.; Kaliontzopoulou, A.; Adams, D.C. geomorph v4. 0 and gmShiny: Enhanced analytics and a new graphical interface for a comprehensive morphometric experience. Methods Ecol. Evol. 2021, 12, 2355–2363. [Google Scholar] [CrossRef]
- Perazzo, G.X.; Corrêa, F.; Salzburger, W.; Gava, A. Morphological differences between an artificial lentic and adjacent lotic environments in a characid species. Rev. Fish Biol. Fish. 2019, 29, 935–949. [Google Scholar] [CrossRef]
- Schofield, P.; Loftus, W.; Fontaine, J. Salinity effects on behavioural response to hypoxia in the non-native Mayan cichlid Cichlasoma urophthalmus from Florida Everglades wetlands. J. Fish Biol. 2009, 74, 1245–1258. [Google Scholar] [CrossRef]
- Crispo, E.; Chapman, L.J. Temporal variation in population genetic structure of a riverine African cichlid fish. J. Hered. 2010, 101, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Burleson, M.; Wilhelm, D.; Smatresk, N. The influence of fish size size on the avoidance of hypoxia and oxygen selection by largemouth bass. J. Fish Biol. 2001, 59, 1336–1349. [Google Scholar] [CrossRef]
- Reddon, A.R.; Hurd, P.L. Water pH during early development influences sex ratio and male morph in a West African cichlid fish, Pelvicachromis pulcher. Zoology 2013, 116, 139–143. [Google Scholar] [CrossRef]
- O’Dea, R.E.; Lagisz, M.; Hendry, A.P.; Nakagawa, S. Developmental temperature affects phenotypic means and variability: A meta-analysis of fish data. Fish Fish. 2019, 20, 1005–1022. [Google Scholar] [CrossRef] [Green Version]
- Haas, T.C.; Blum, M.J.; Heins, D.C. Morphological responses of a stream fish to water impoundment. Biol. Lett. 2010, 6, 803–806. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DF | SS | MS | Rsq | F | Z | Pr (<F) | |
|---|---|---|---|---|---|---|---|
| PC1 | 1 | 0.001685 | 0.001685 | 0.01283 | 1.5603 | 1.2106 | 0.12 |
| PC2 | 1 | 0.003574 | 0.0035741 | 0.02722 | 3.3095 | 2.7622 | 0.003 ** |
| PC3 | 1 | 0.011577 | 0.0115768 | 0.08816 | 10.7197 | 5.0011 | 0.001 ** |
| Residuals | 106 | 0.114475 | 0.00108 | 0.87179 | |||
| Total | 109 | 0.131311 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernandez, J.; Villalobos-Leiva, A.; Bermúdez, A.; Ahumada-C, D.; Suazo, M.J.; Correa, M.; Díaz, A.; Benítez, H.A. Ecomorphology and Morphological Disparity of Caquetaia Kraussii (Perciformes: Cichlidae) in Colombia. Animals 2022, 12, 3438. https://doi.org/10.3390/ani12233438
Hernandez J, Villalobos-Leiva A, Bermúdez A, Ahumada-C D, Suazo MJ, Correa M, Díaz A, Benítez HA. Ecomorphology and Morphological Disparity of Caquetaia Kraussii (Perciformes: Cichlidae) in Colombia. Animals. 2022; 12(23):3438. https://doi.org/10.3390/ani12233438
Chicago/Turabian StyleHernandez, Jordan, Amado Villalobos-Leiva, Adriana Bermúdez, Daniela Ahumada-C, Manuel J. Suazo, Margarita Correa, Angie Díaz, and Hugo A. Benítez. 2022. "Ecomorphology and Morphological Disparity of Caquetaia Kraussii (Perciformes: Cichlidae) in Colombia" Animals 12, no. 23: 3438. https://doi.org/10.3390/ani12233438