
Effects of Starch Overload and Cecal Buffering on Fecal Microbiota of Horses


 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Animals, Facilities, and Management
2.2. Experimental Design
2.3. Procedures
2.4. Data Processing
2.5. Bioinformatics and Statistical Analyses
3. Results
3.1. Clinical Findings
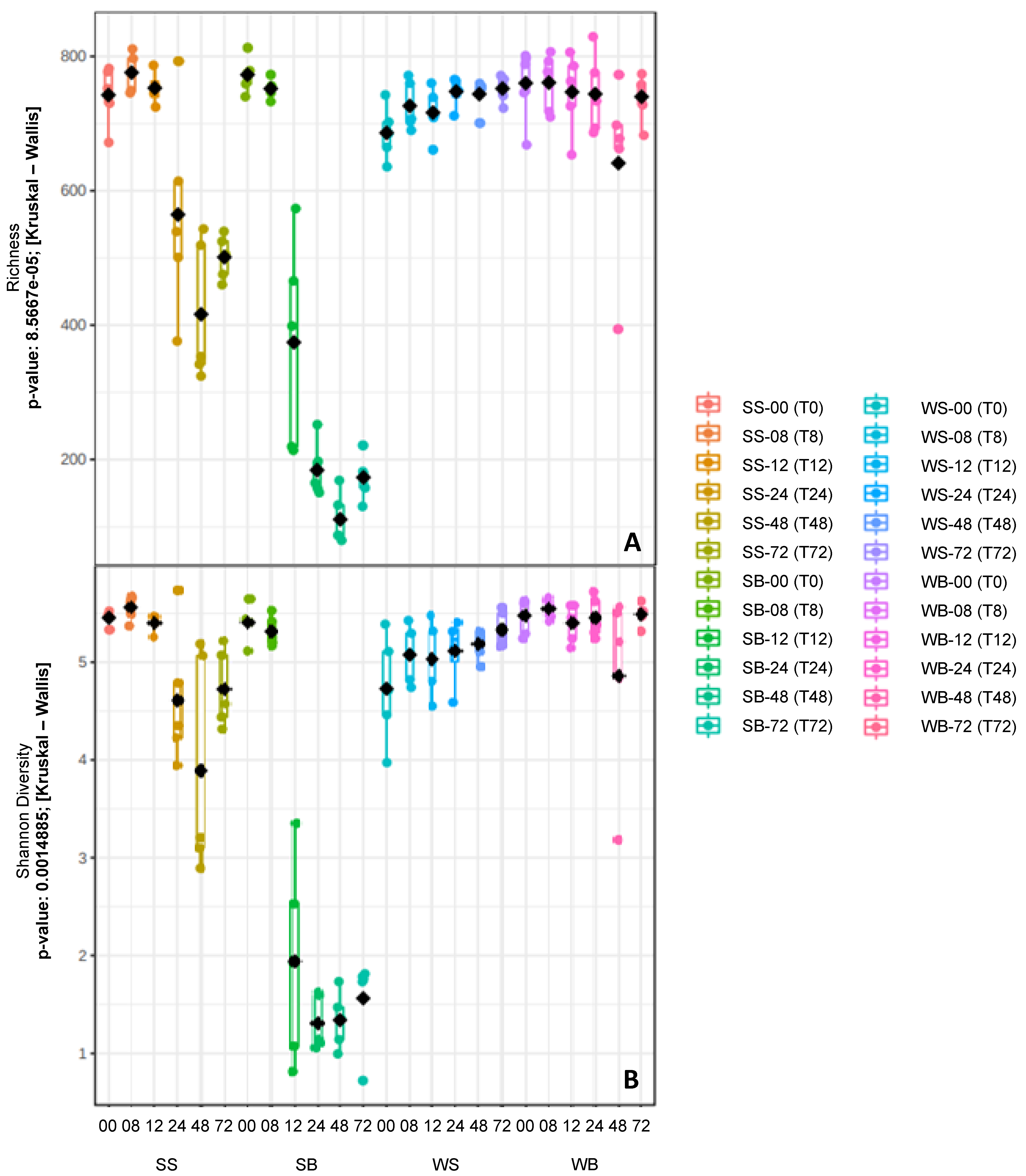
3.2. Sequencing Information and Comparison of the Bacterial Communities
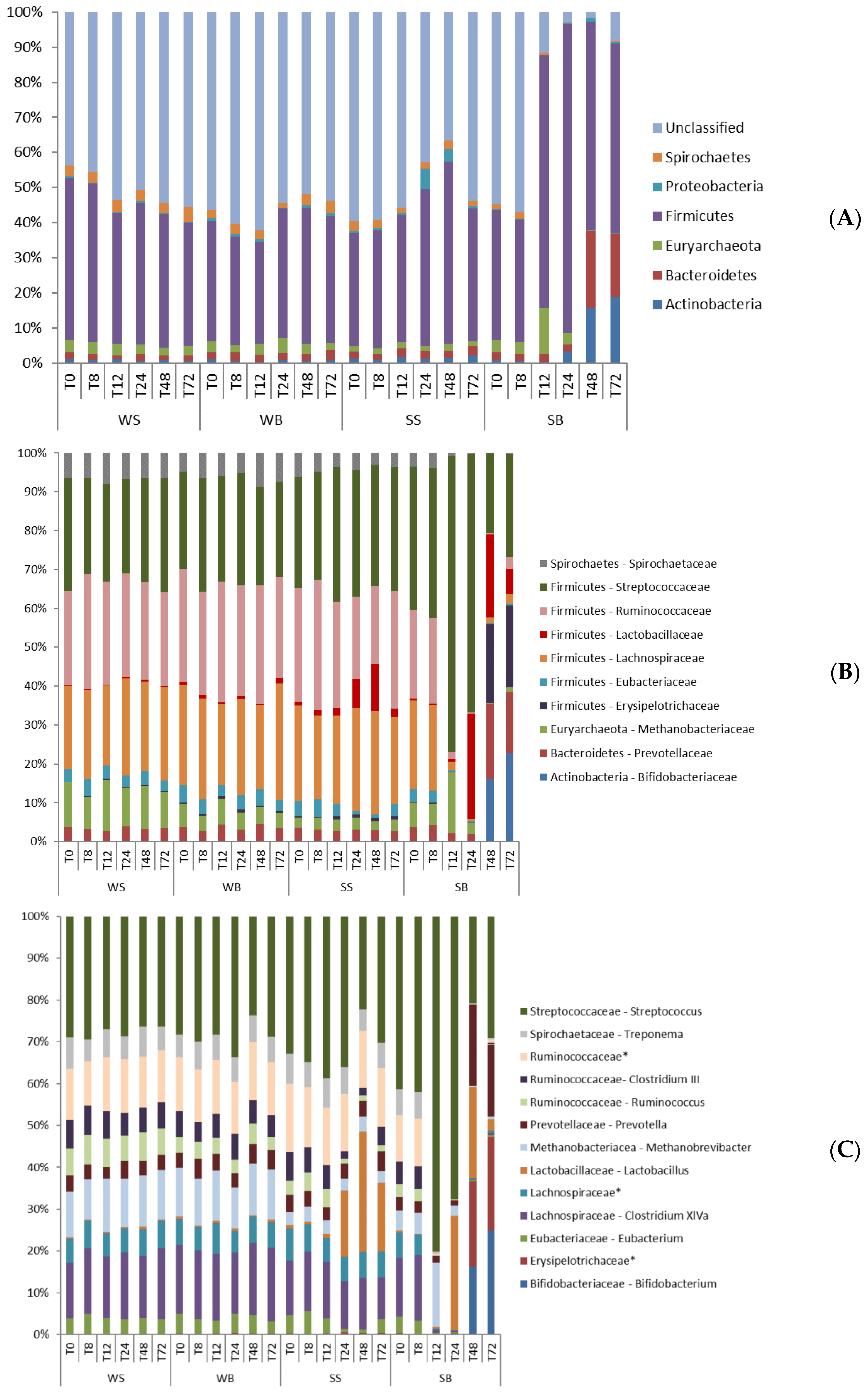
3.3. Taxonomic Composition of Bacterial Communities
3.4. Microbial Composition x Clinical Alterations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grimm, P.; Julliand, V.; Philippeau, C.; Sadet-Bourgeteau, S. Effect of yeast supplementation on hindgut microbiota and digestibility of horses subjected to an abrupt change of hays. Livest. Sci. 2016, 186, 34–40. [Google Scholar] [CrossRef]
- Milinovich, G.J.; Klieve, A.V.; Pollitt, C.C.; Trott, D.J. Microbial events in the hindgut during carbohydrate-induced equine laminitis. Vet. Clin. Equine Prat. 2010, 26, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Kamada, N.; Seo, S.U.; Chen, G.Y.; Nuñez, G. Role of the gut microbiota in immunity and inflammatory disease. Nat. Rev. Immunol. 2013, 13, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Vieira, S.M.; Hiltensperger, M.; Kumar, V.; Zegarra-Ruiz, D.; Dehner, C.; Khan, N.; Costa, F.R.C.; Tiniakou, E.; Greiling, T.; Ruff, W.; et al. Translocation of a gut pathobiont drives autoimmunity in mice and humans. Science 2018, 359, 1156–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, M.C.; Weese, J.S. Understanding the intestinal microbiome in health and disease. Vet. Clin. Equine Prat. 2018, 34, 1–12. [Google Scholar] [CrossRef]
- Garber, A.; Hastie, P.; Murray, J.A. Factors influencing equine gut microbiota: Current knowledge. J. Equine Vet. Sci. 2020, 88, 102943. [Google Scholar] [CrossRef]
- Al Jassim, R.A.; Andrews, F.M. The bacterial community of the horse gastrointestinal tract and its relation to fermentative acidosis; laminits; colic and stomach ulcers. Vet. Clin. Equine Pract. 2009, 25, 199–215. [Google Scholar] [CrossRef]
- Stewart, H.L.; Pitta, D.; Indugu, N.; Vecchiarelli, B.; Hennessy, M.L.; Engiles, J.B.; Southwood, L.L. Changes in the faecal bacterial microbiota during hospitalisation of horses with colic and the effect of different causes of colic. Equine Vet. J. 2020, 53, 1–13. [Google Scholar] [CrossRef]
- Moreau, M.M.; Eades, S.C.; Reinemeyer, C.R.; Fugaro, M.N.; Onishi, J.C. Illumina sequencing of the V4 hypervariable region 16S rRNA gene reveals extensive changes in bacterial communities in the cecum following carbohydrate oral infusion and development of early-stage acute laminitis in the horse. Vet. Microbiol. 2014, 168, 436–441. [Google Scholar] [CrossRef]
- Garner, H.E.; Moore, J.N.; Johnson, J.H.; Clark, L.; Amend, J.F.; Tritschler, L.G.; Coffmann, J.R.; Sprouse, R.F.; Hutcheson, D.P.; Salem, C.A. Changes in the caecal flora associated with the onset of laminitis. Equine Vet. J. 1978, 10, 249–252. [Google Scholar] [CrossRef]
- Costa, R.S.; Botteon, P.T.; Ferraz, V.; Leme, F.O.P.; Faleiros, R.R.; Alves, G.E.S. Variações plasmáticas de aminas vasoativas em equinos sob efeito de sobrecarga de carboidratos e anti-inflamatórios não esteroidais. Ciência Anim. Bras. 2012, 13, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Warzecha, C.M.; Coverdale, J.A.; Janecka, J.E.; Leatherwood, J.L.; Pinchak, W.E.; Wickersham, T.A.; McCann, J.C. Influence of short-term dietary starch inclusion on the equine cecal microbiome. J. Anim. Sci. 2017, 95, 5077–5090. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, M.; Hoskin, S.O.; Rogers, C.W.; Grinberg, A. Fecal pH and microbial populations in thoroughbred horses during transition from pasture to concentrate feeding. J. Equine Vet. Sci. 2013, 33, 215–222. [Google Scholar] [CrossRef]
- Goodson, J.; Tyznik, W.J.; Cline, J.H.; Dehority, B.A. Effects of an abrupt diet change from hay to concentrate on microbial numbers and physical environment in the cecum of the pony. Appl. Environ. Microbiol. 1988, 54, 1946–1950. [Google Scholar] [CrossRef] [Green Version]
- Harris, P.; Dunnett, C. Nutritional tips for veterinarians. Equine Vet. Educ. 2018, 30, 486–496. [Google Scholar] [CrossRef]
- Miller, J.K.; Slebondzinska-Brzezinska, E.; Madsen, F.C. Oxidative stress; antioxidants; and animal function. J. Dairy Sci. 1993, 76, 2812–2823. [Google Scholar] [CrossRef]
- Julliand, V.; Grimm, P. The Impact of Diet on the Hindgut Microbiome. J. Equine Vet. Sci. 2017, 52, 23–28. [Google Scholar] [CrossRef]
- Biddle, A.S.; Black, S.J.; Blanchard, J.L. An in vitro model of the horse gut microbiome enables identification of lactate-utilizing bacteria that differentially respond to starch induction. PLoS ONE 2013, 8, e77599. [Google Scholar] [CrossRef] [Green Version]
- Kuhl, J.; Winterhoff, N.; Wulf, M.; Schweigert, F.J.; Schwendenwein, I.; Bruckmaier, R.M.; Aurich, J.E.; Kutzer, P.; Aurich, C. Changes in faecal bacteria and metabolic parameters in foals during the first six weeks of life. Vet. Microbiol. 2011, 151, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Daly, K.; Proudman, C.J.; Duncan, S.H.; Flint, H.J.; Dyer, J.; Shirazi-Beechey, S.P. Alterations in microbiota and fermentation products in equine large intestine in response to dietary variation and intestinal disease. Br. J. Nutr. 2012, 107, 989–995. [Google Scholar] [CrossRef]
- Park, T.; Cheong, H.; Yoon, J.; Kim, A.; Yun, Y.; Unno, T. Comparison of the fecal microbiota of horses with intestinal disease and their healthy counterparts. Vet. Sci. 2021, 8, 113. [Google Scholar] [CrossRef] [PubMed]
- Maton, P.N.; Burton, M.E. Antacids revisited: A review of their clinical pharmacology and recommended therapeutic use. Drugs 1999, 57, 855–870. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.E.; Constable, P.D.; Misk, N.A. Effect of an orally administered antacid agent containing aluminum hydroxide and magnesium hydroxide on abomasal luminal pH in clinically normal milk-fed calves. J. Am. Vet. Med. Assoc. 2002, 1, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Clark, C.K.; Merritt, A.M.; Burrow, J.A.; Steible, C.K. Effect of aluminum hydroxide/magnesium hydroxide antacid and bismuth subsalicylate on gastric pH in horses. J. Am. Vet. Med. Assoc. 1996, 8, 1687–1691. [Google Scholar] [PubMed]
- Tillotson, K.; Traub-Dargatz, J.L. Gastrointestinal protectants and cathartics. Vet. Clin. Equine Pract. 2003, 19, 599–615. [Google Scholar] [CrossRef]
- McGorum, B.C.; Pirie, R.S. Antimicrobial associated diarrhoea in the horse. Part 1: Overview; pathogenesis; and risk factors. Equine Vet. Educ. 2009, 21, 610–616. [Google Scholar] [CrossRef]
- Maia, M.A.; Botteon, P.T.L.; Spindola, B.F.; Botteon, R.C.C.M. Alteration of the fecal pH of the equines by oral administration of alkalizing agents. Braz. J. Vet. Med. 2017, 39, 1–6. [Google Scholar] [CrossRef]
- Uribe Diaz, A.P.; Santana, A.E.; Valadão, C.A.A.; Souza, A.H. Canulação cecal em equinos. Ciência Anim. Bras. 2010, 11, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Santos, T.M.; Almeida, F.Q.; Godoi, F.N.; Silva, V.P.; França, A.B.; Santiago, J.M.; Santos, C.S. Capacidade tamponante; pH e consistência das fezes em equinos submetidos à sobrecarga dietética com amido. Ciência Rural. 2009, 39, 1782–1788. [Google Scholar] [CrossRef] [Green Version]
- Berg, E.L.; Fu, C.J.; Porter, J.H.; Kerley, M.S. Fructooligosaccharide supplementation in the yearling horse: Effects on fecal pH; microbial content; and volatile fatty acid concentrations. J. Anim. Sci. 2005, 83, 1549–1553. [Google Scholar] [CrossRef]
- Obel, N. Studies on the Histopathology of Acute Laminitis. Master’s Thesis, Almqvist and Wiksells, Uppsala, Sweden, 1948. [Google Scholar]
- Stewart, A.S.; Pratt-Phillips, S.; Gonzalez, L.M. Alterations in intestinal permeability: The role of the “Leaky Gut” in health and disease. J. Equine Vet. Sci. 2018, 52, 10–22. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, C.C.; Kishi, L.T.; Lopes, E.M.; Omori, W.P.; Souza, J.A.M.; Alves, L.M.C.; Lemos, E.G.M. Bacterial communities in mining soils and surrounding areas under regeneration process in a former ore mine. Braz. J. Microbiol. 2018, 1, 1–14. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [Green Version]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst—A web-based tool for comprehensive statistical; visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef]
- Lerner, A.; Aminov, R.; Matthias, T. Dysbiosis may trigger autoimmune diseases via inappropriate post-translational modification of host proteins. Front. Microbiol. 2016, 7, 84. [Google Scholar] [CrossRef]
- Elzinga, S.E.; Weese, J.S.; Adams, A.A. Comparison of the fecal microbiota in horses with Equine Metabolic Syndrome and metabolically normal controls fed a similar all-forage diet. J. Equine Vet. Sci. 2016, 44, 9–16. [Google Scholar] [CrossRef]
- Garner, H.E.; Coffman, J.R.; Hahn, A.W.; Hutcheson, D.P.; Tumbleson, M.E. Equine laminitis of alimentary origin: An experimental model. Am. J. Vet. Res. 1975, 36, 441–445. [Google Scholar] [PubMed]
- Morrison, P.K.; Newbold, C.J.; Jones, E.; Worgan, H.J.; Grove-White, D.H.; Dugdale, A.H.; Barfoot, C.; Harris, P.A.; Argo, C.M.G. The equine gastrointestinal microbiome: Impacts of age and obesity. Front. Microbiol. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Dougal, K.; De La Fuente, G.; Harris, P.A.; Girdwood, S.E.; Pinloche, E.; Geor, R.J.; Nielsen, B.D.; Schott, I.I.H.C.; Elzinga, S.; Newbold, C.J. Characterization of the faecal bacterial community in adult an elderly horses fed a high fiber; high oil or high starch diet using 454 pyrosequencing. PLoS ONE 2014, 9, e87424. [Google Scholar] [CrossRef]
- Bulmer, L.S.; Murray, J.A.; Burns, N.M.; Garber, A.; Wemelsfelder, F.; McEwan, N.R.; Hastie, P.M. High-starch diets alter equine faecal microbiota and increase behavioural reactivity. Sci. Rep. 2019, 9, 18621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coenen, M.; Vervuert, I. A minimum of roughage and a maximum of starch necessary benchmarks for equine diets. Pferdeheilkunde 2010, 26, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Picard, C.; Fioramonti, J.; Francois, A.; Robinson, T.; Neant, F.; Matuchansky, C. Bifidobacteria as probiotic agents–physiological effects and clinical benefits. Aliment. Pharmacol. Ther. 2005, 22, 495–512. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chen, L.; Zhou, R.; Wang, X.; Song, L.; Huang, S.; Wang, G.; Xia, B. Increased proportions of Bifidobacterium and the Lactobacillus group and loss of butyrate-producing bacteria in inflammatory bowel disease. J. Clin. Microbiol. 2014, 52, 398–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Million, M.; Tidjani Alou, M.; Khelaifia, S.; Bachar, D.; Lagier, J.-C.; Dione, N.; Brah, S.; Hugon, P.; Lombard, V.; Armougom, F. Increased gut redox and depletion of anaerobic and methanogenic prokaryotes in severe acute malnutrition. Sci. Rep. 2016, 6, 26051. [Google Scholar] [CrossRef] [Green Version]
- Van Soest, P.J. Nutritional Ecology of the Ruminant, 2nd ed.; Cornell University Press: London, UK, 1994; pp. 230–289. [Google Scholar]
- Milani, C.; Lugli, G.A.; Duranti, S.; Turroni, F.; Mancabelli, L.; Ferrario, C.; Mangifesta, M.; Hevia, A.; Viappiani, A.; Scholz, M. Bifidobacteria exhibit social behavior through carbohydrate resource sharing in the gut. Sci. Rep. 2015, 5, 15782. [Google Scholar] [CrossRef] [Green Version]
- O’Callaghan, A.; van Sinderen, D. Bifidobacteria and their role as members of the human gut microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef] [Green Version]
- Goer, R. Current concepts on the pathophysiology of pasture-associated laminitis. Vet. Clin. Equine Pract. 2010, 26, 265–276. [Google Scholar] [CrossRef]
- Kim, K.A.; Jeong, J.J.; Yoo, S.Y.; Kim, D.H. Gut microbiota lipopolysaccharide accelerates inflamm-agging in mice. BMC Microbiol. 2016, 16, 1–9. [Google Scholar] [CrossRef] [Green Version]
- McCall, I.C.; Betanzos, A.; Weber, D.A.; Nava, P.; Miller, G.W.; Parkos, C.A. Effects of phenol on barrier function of a human intestinal epithelial cell line correlate with altered tight junction protein localization. Toxicol. Appl. Pharmacol. 2009, 241, 61–67. [Google Scholar] [CrossRef]
- Jakobsson, H.E.; Rodriguez-Pineiro, A.M.; Schutte, A.; Ermund, A.; Boysen, P.; Bemark, M.; Sommer, F.; Backhed, F.; Hansson, G.C.; Johansson, M.E.V. The composition of the gut microbiota shapes the colon mucus barrier. EMBO Rep. 2015, 16, 164–177. [Google Scholar] [CrossRef]
- Hankel, J.; Mößeler, A.; Hartung, C.B.; Rath, S.; Schulten, L.; Visscher, C.; Kamphues, J.; Vital, M. Responses of ileal and fecal microbiota to withdrawal of pancreatic enzyme replacement therapy in a porcine model of exocrine pancreatic insufficiency. Int. J. Mol. Sci. 2022, 23, 11700. [Google Scholar] [CrossRef]
- Pryde, S.E.; Duncan, S.H.; Hold, G.L.; Stewart, C.S.; Flint, H.J. The microbiology of butyrate formation in the human colon. FEMS Microbiol. Lett. 2002, 217, 133–139. [Google Scholar] [CrossRef]
- Harlow, B.E.; Lawrence, L.M.; Hayes, S.H.; Crum, A.; Flythe, M.D. Effect of dietary starch source and concentration on equine fecal microbiota. PLoS ONE 2016, 11, e0154037. [Google Scholar] [CrossRef] [Green Version]
- Ze, X.; Le Mougen, F.; Duncan, S.H.; Louis, P.; Flint, H.J. Some are more equal than others: The role of “keystone” species in the degradation of recalcitrant substrates. Gut Microbes 2013, 4, 236–240. [Google Scholar] [CrossRef] [Green Version]
- Belknap, J.K.; Black, S.J. Sepsis-related laminitis. Equine Vet. J. 2012, 44, 738–739. [Google Scholar] [CrossRef]
- Lopes, M.A.F.; White, I.I.N.A.; Donaldson, L.; Crisman, M.V.; Ward, D.L. Effects of enteral and intravenous fluid therapy; magnesium sulfate; and sodium sulfate on colonic contents and feces in horses. Am. J. Vet. Res. 2004, 65, 695–704. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Hintz, H.F.; Schryver, H.F. Magnesium metabolism in the horse. J. Anim. Sci. 1972, 35, 755–759. [Google Scholar] [CrossRef]
- Schryver, H.F.; Parker, M.T.; Daniluk, P.D.; Pagan, K.I.; Williams, J.; Soderholm, L.V.; Hintz, H.F. Salt consumption and the effect of salt on mineral metabolism in horses. Cornell Vet. 1987, 77, 122–131. [Google Scholar]
- Hintz, H.F.; Cymbaluk, N.F. Nutrition of the horse. Annu. Rev. Nutr. 1994, 14, 243–267. [Google Scholar] [CrossRef] [PubMed]
- Franzan, B.C.; Coelho, I.S.; De Souza, M.T.; Santos, M.M.M.; De Almeida, F.Q.; Silva, V.P. Fecal Microbiome Responses to Sudden Diet Change in Mangalarga Marchador horses. J. Equine Vet. Sci. 2022, 108, 103803. [Google Scholar] [CrossRef] [PubMed]
- Blackmore, T.M.; Dugdale, A.; Argo, C.M.; Curtis, G.; Pinloche, E.; Harris, P.A.; Worgan, H.J.; Girdwood, S.E.; Dougal, K.; Newbold, C.J.; et al. Strong stability and host specific bacterial community in faeces of ponies. PLoS ONE 2013, 8, e75079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepherd, M.L.; Swecker, W.S.J.; Jensen, R.V.; Ponder, M.A. Characterization of the fecal bacteria communities of forage-fed horses by pyrosequencing of 16S rRNA V4 gene amplicons. FEMS Microbiol. Lett. 2012, 326, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Almeida, M.L.M.; Júnior, W.H.F.; Carvalho, J.R.G.; Rodrigues, I.M.; Jordão, L.R.; Fonseca, M.G.; Rezende, A.S.C.; Neto, A.Q.; Weese, J.S.; Costa, M.C.; et al. Intense exercise and aerobic conditioning associated with chromium or L-carnitine supplementation modified the fecal microbiota of fillies. PLoS ONE 2016, 11, e0167108. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Groups | Treatments | |
|---|---|---|
| I (n = 5) | WS | Water (10 L) administered by nasogastric tube and, after 8 hours, 5 L of NaCl 0.9% solution administered intracecally. |
| SB | Corn starch overload ‡ diluted in water (10 L) administered by nasogastric tube and, after 8 hours, 5 L of a buffer † diluted in NaCl 0.9% solution administered intracecally. | |
| II (n = 5) | SS | Corn starch overload ‡ diluted in water (10 L) administered by nasogastric tube and, after 8 hours, 5 L of NaCl 0.9% solution administered intracecally. |
| WB | Water (10 L) administered by nasogastric tube and, after 8 hours, 5 L of a buffer † diluted in NaCl 0.9% solution administered intracecally. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bustamante, C.C.; de Paula, V.B.; Rabelo, I.P.; Fernandes, C.C.; Kishi, L.T.; Canola, P.A.; Lemos, E.G.d.M.; Valadão, C.A.A. Effects of Starch Overload and Cecal Buffering on Fecal Microbiota of Horses. Animals 2022, 12, 3435. https://doi.org/10.3390/ani12233435
Bustamante CC, de Paula VB, Rabelo IP, Fernandes CC, Kishi LT, Canola PA, Lemos EGdM, Valadão CAA. Effects of Starch Overload and Cecal Buffering on Fecal Microbiota of Horses. Animals. 2022; 12(23):3435. https://doi.org/10.3390/ani12233435
Chicago/Turabian StyleBustamante, Caio C., Vanessa B. de Paula, Isabela P. Rabelo, Camila C. Fernandes, Luciano T. Kishi, Paulo A. Canola, Eliana Gertrudes de M. Lemos, and Carlos Augusto A. Valadão. 2022. "Effects of Starch Overload and Cecal Buffering on Fecal Microbiota of Horses" Animals 12, no. 23: 3435. https://doi.org/10.3390/ani12233435