
Effects of Dietary Supplementation with Vitamin A on Antioxidant and Intestinal Barrier Function of Broilers Co-Infected with Coccidia and Clostridium perfringens
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Diets
2.2. Antioxidative Enzymes and Peroxidation Products
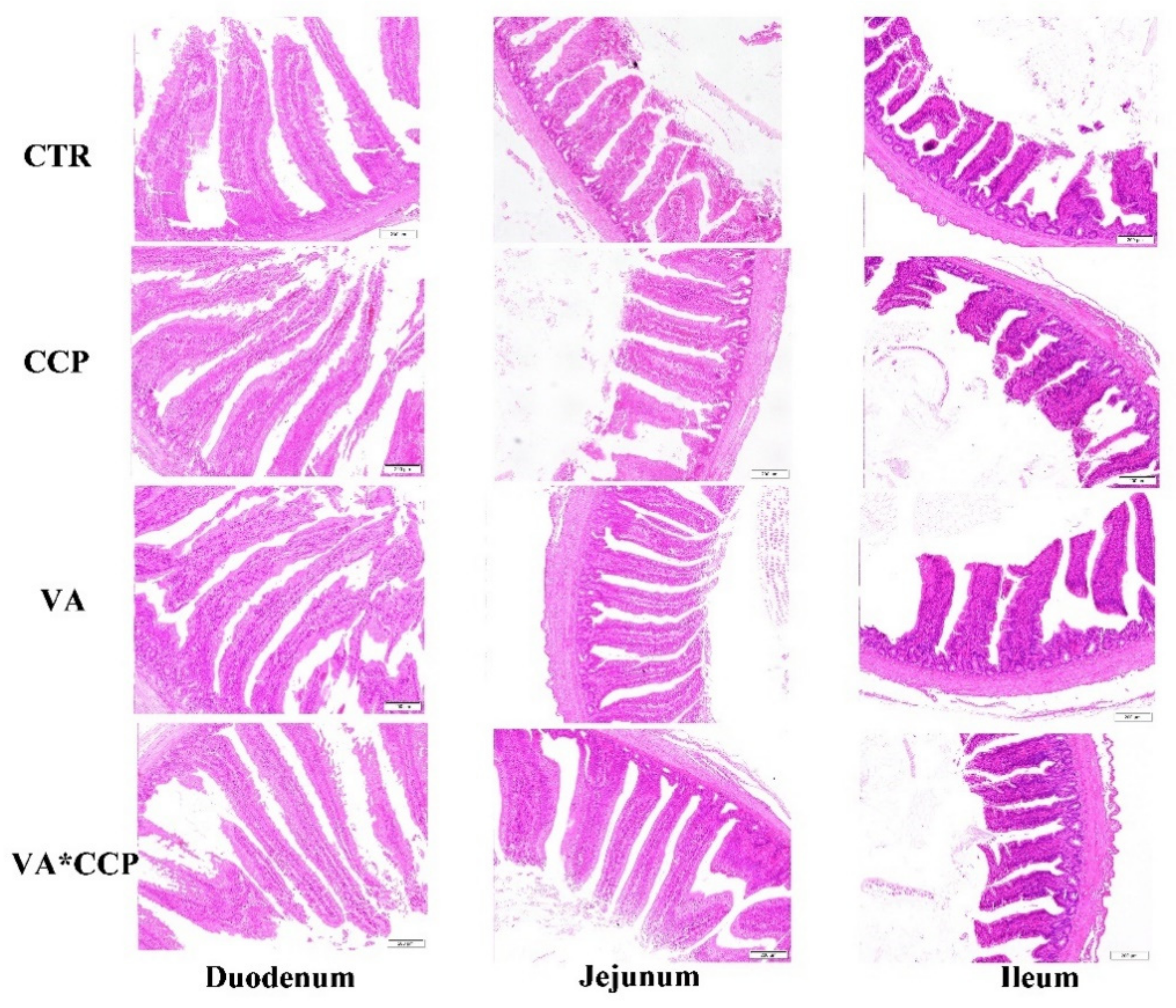
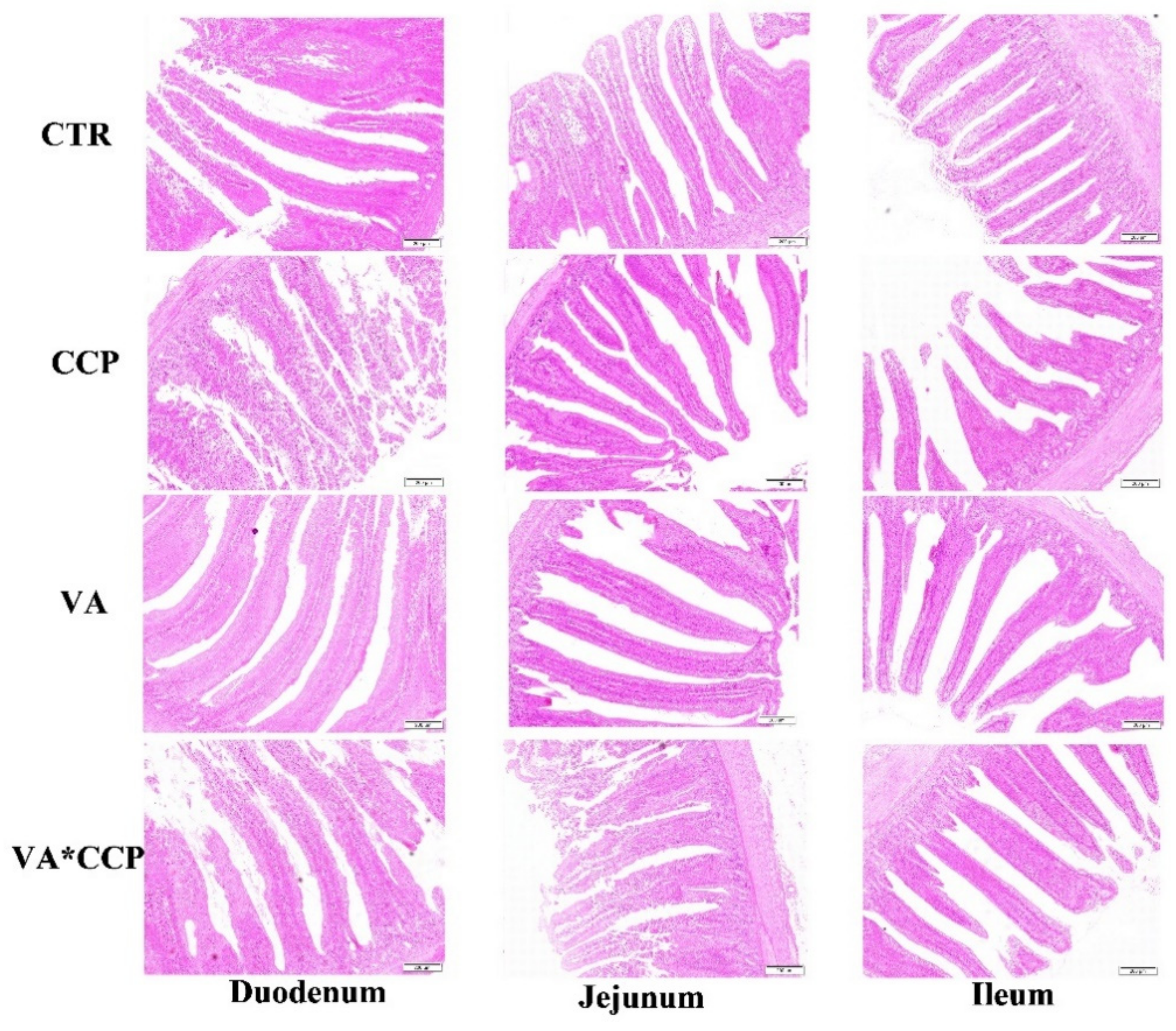
2.3. Serum Diamine Oxidase and Intestinal Morphology
2.4. Transcription Level of Genes in the Liver and Jejunum
2.5. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Antioxidative Enzymes and Peroxidation Products
3.3. The mRNA Levels of Antioxidant-Related Genes
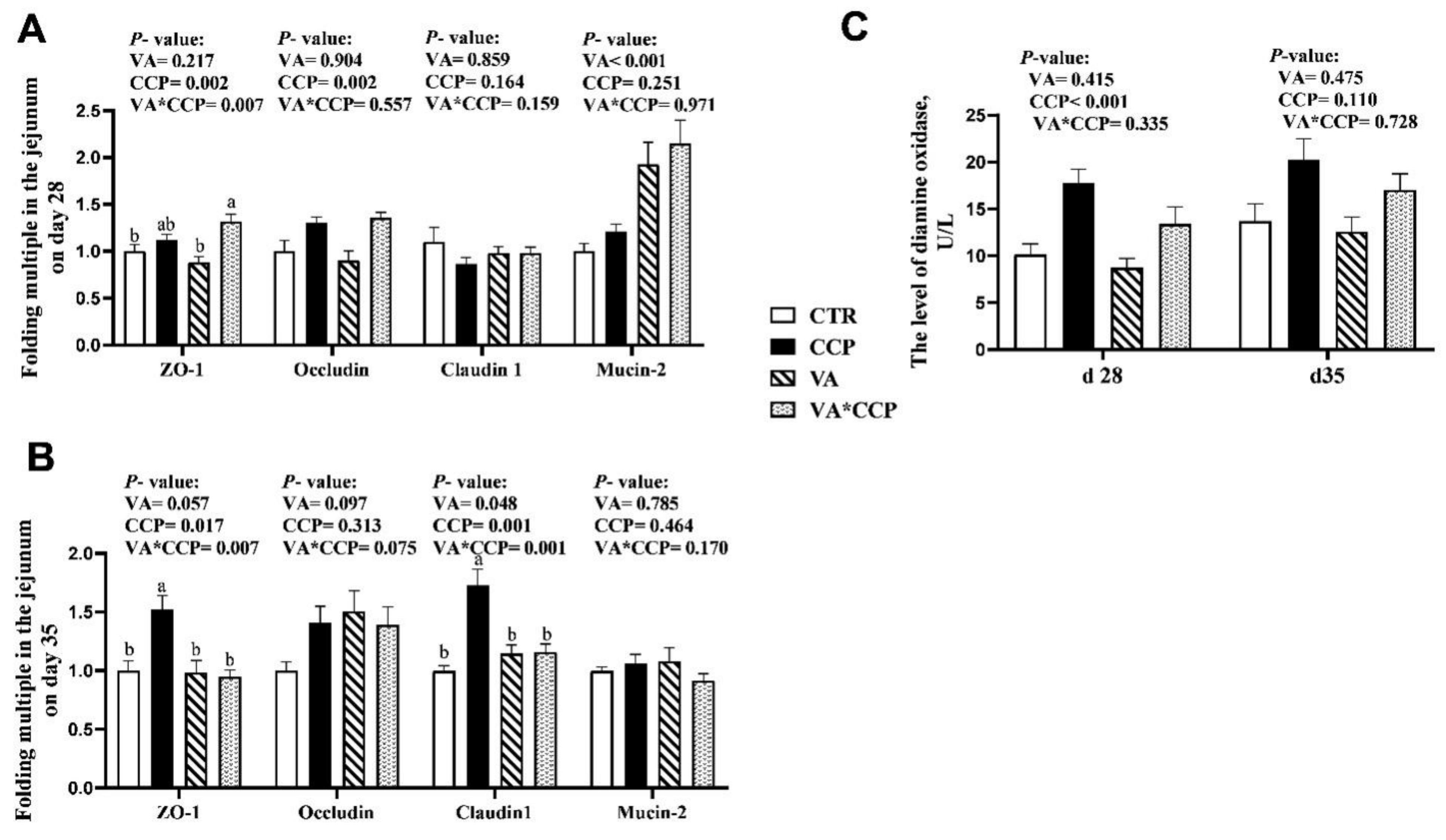
3.4. Intestinal Barrier Function
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Appiah, M.O.; He, B.; Lu, W.; Wang, J. Antioxidative effect of melatonin on cryopreserved chicken semen. Cryobiology 2019, 89, 90–95. [Google Scholar] [CrossRef]
- Balogh, K.; Kovesi, B.; Zandoki, E.; Kulcsar, S.; Ancsin, Z.; Erdelyi, M.; Dobolyi, C.; Bata-Vidacs, I.; Inotai, K.; Szekeres, A.; et al. Effect of sterigmatocystin or aflatoxin contaminated feed on lipid peroxidation and glutathione redox system and expression of glutathione redox system regulatory genes in broiler chicken. Antioxidants 2019, 8, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Zhao, C.; Zhang, M.; Zhang, S.; Zhai, J.; Gao, X.; Liu, C.; Lv, X.; Zheng, S. Changes in oxidation-antioxidation function on the thymus of chickens infected with reticuloendotheliosis virus. BMC Vet. Res. 2020, 16, 483. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Ning, F.; Zhang, J.; Tang, Y.; Teng, X. NF-kappaB pathway took part in the development of apoptosis mediated by miR-15a and oxidative stress via mitochondrial pathway in ammonia-treated chicken splenic lymphocytes. Sci. Total Environ. 2020, 729, 139017. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Moore, R.J.; Stanley, D.; Chousalkar, K.K. The gut microbiota of laying hens and its manipulation with prebiotics and probiotics to enhance gut health and food safety. Appl. Environ. Microbiol. 2020, 86, e00600-20. [Google Scholar] [CrossRef]
- Li, Q.; Wan, G.; Peng, C.; Xu, L.; Yu, Y.; Li, L.; Li, G. Effect of probiotic supplementation on growth performance, intestinal morphology, barrier integrity, and inflammatory response in broilers subjected to cyclic heat stress. Anim. Sci. J. 2020, 91, e13433. [Google Scholar] [CrossRef]
- Yu, C.; Tong, Y.; Li, Q.; Wang, T.; Yang, Z. Trans-anethole ameliorates intestinal injury through activation of Nrf2 signaling pathway in subclinical necrotic enteritis-induced broilers. Front. Vet. Sci. 2022, 9, 877066. [Google Scholar] [CrossRef]
- Memon, F.U.; Yang, Y.; Lv, F.; Soliman, A.M.; Chen, Y.; Sun, J.; Wang, Y.; Zhang, G.; Li, Z.; Xu, B.; et al. Effects of probiotic and Bidens pilosa on the performance and gut health of chicken during induced Eimeria tenella infection. J. Appl. Microbiol. 2021, 131, 425–434. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, Y.; Chen, Y.; Li, Y.; Jia, P.; Ji, S.; Zhou, Y.; Wang, T. Dietary pterostilbene supplementation attenuates intestinal damage and immunological stress of broiler chickens challenged with lipopolysaccharide. J. Anim. Sci. 2020, 98, skz373. [Google Scholar] [CrossRef]
- Liu, D.; Guo, Y.; Wang, Z.; Yuan, J. Exogenous lysozyme influences Clostridium perfringens colonization and intestinal barrier function in broiler chickens. Avian Pathol. 2010, 39, 17–24. [Google Scholar] [CrossRef]
- Howell, J.M.; Thompson, J.N. Observations on the lesions in vitamin A deficient adult fowls with particular reference to changes in bone and central nervous system. Br. J. Exp. Pathol. 1967, 48, 450–454. [Google Scholar]
- Wang, Y.; Li, L.; Gou, Z.; Chen, F.; Fan, Q.; Lin, X.; Ye, J.; Zhang, C.; Jiang, S. Effects of maternal and dietary vitamin A on growth performance, meat quality, antioxidant status, and immune function of offspring broilers. Poult. Sci. 2020, 99, 3930–3940. [Google Scholar] [CrossRef]
- Li, R.; Yang, Y.; Hong, P.; Zhang, Z.; Li, L.; Hui, J.; Zheng, X. Beta-carotene attenuates weaning-induced apoptosis via inhibition of PERK-CHOP and IRE1-JNK/p38 MAPK signalling pathways in piglet jejunum. J. Anim. Physiol. Anim. Nutr. 2020, 104, 280–290. [Google Scholar] [CrossRef]
- Pattanakitsakul, P.; Chongviriyaphan, N.; Pakakasama, S.; Apiwattanakul, N. Effect of vitamin A on intestinal mucosal injury in pediatric patients receiving hematopoietic stem cell transplantation and chemotherapy: A quasai-randomized trial. BMC Res. Notes 2020, 13, 464. [Google Scholar] [CrossRef]
- Hui, J.; Li, L.; Li, R.; Wu, M.; Yang, Y.; Wang, J.; Fan, Y.; Zheng, X. Effects of supplementation with beta-carotene on the growth performance and intestinal mucosal barriers in layer-type cockerels. Anim. Sci. J. 2020, 91, e13344. [Google Scholar] [CrossRef]
- Li, P.; Gao, M.; Fu, J.; Yan, S.; Liu, Y.; Mahmood, T.; Lv, Z.; Guo, Y. Dietary soya saponin improves the lipid metabolism and intestinal health of laying hens. Poult. Sci. 2022, 101, 101663. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, L.; Zhang, W.; Yang, Z.; Ding, B.; Zhu, H.; Liu, Y.; Qiu, Y.; Yin, Y.; Wu, G. Protective effects of N-acetylcysteine on intestinal functions of piglets challenged with lipopolysaccharide. Amino Acids 2012, 43, 1233–1242. [Google Scholar] [CrossRef]
- Heid, C.A.; Stevens, J.; Livak, K.J.; Williams, P.M. Real time quantitative PCR. Genome Res. 1996, 6, 986–994. [Google Scholar] [CrossRef] [Green Version]
- Latorre, J.D.; Adhikari, B.; Park, S.H.; Teague, K.D.; Graham, L.E.; Mahaffey, B.D.; Baxter, M.; Hernandez-Velasco, X.; Kwon, Y.M.; Ricke, S.C.; et al. Evaluation of the epithelial barrier function and ileal microbiome in an established necrotic enteritis challenge model in broiler chickens. Front. Vet. Sci. 2018, 5, 199. [Google Scholar] [CrossRef]
- Guo, S.; Xi, Y.; Xia, Y.; Wu, T.; Zhao, D.; Zhang, Z.; Ding, B. Dietary Lactobacillus fermentum and Bacillus coagulans supplementation modulates intestinal immunity and microbiota of broiler chickens challenged by Clostridium perfringens. Front. Vet. Sci. 2021, 8, 680742. [Google Scholar] [CrossRef]
- La Ragione, R.M.; Woodward, M.J. Competitive exclusion by Bacillus subtilis spores of Salmonella enterica serotype Enteritidis and Clostridium perfringens in young chickens. Vet. Microbiol. 2003, 94, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhang, X.; Wang, Y.; Guo, Y.; Zhu, P.; Li, G.; Zhang, J.; Ma, Q.; Zhao, L. Dietary ellagic acid ameliorated Clostridium perfringens-induced subclinical necrotic enteritis in broilers via regulating inflammation and cecal microbiota. J. Anim. Sci. Biotechnol. 2022, 13, 47. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Lv, Z.; Li, H.; Guo, S.; Liu, D.; Guo, Y. Dietary l-arginine inhibits intestinal Clostridium perfringens colonisation and attenuates intestinal mucosal injury in broiler chickens. Br. J. Nutr. 2017, 118, 321–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jelic, M.D.; Mandic, A.D.; Maricic, S.M.; Srdjenovic, B.U. Oxidative stress and its role in cancer. J. Cancer Res. Ther. 2021, 17, 22–28. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Zhou, M.; Xu, W.; Wang, J.; Yan, J.; Shi, Y.; Zhang, C.; Ge, W.; Wu, J.; Du, P.; Chen, Y. Boosting mTOR-dependent autophagy via upstream TLR4-MyD88-MAPK signalling and downstream NF-kappa B pathway quenches intestinal inflammation and oxidative stress injury. eBio Med. 2018, 35, 345–360. [Google Scholar]
- Sun, Z.Y.; Peng, T.; Zhang, M.J.; Cheng, W.U.; Luo, Y.X. Effects of β-Carotenes on Blood Component and Activities of SOD, GSH-PX and CAT in Mice. Prog. Vet. Med. 2007, 28, 25–28. (In Chinese) [Google Scholar]
- Gall, T.K.; Tomljanovic, M.; Jaganjac, M.; Matijevic, G.T.; Cipak, G.A.; Milkovic, L.; Borovic, S.S.; Buttari, B.; Profumo, E.; Saha, S.; et al. Oxidative stress and cancer heterogeneity orchestrate NRF2 roles relevant for therapy response. Molecules 2022, 27, 1468. [Google Scholar] [CrossRef]
- Leeson, S.; Caston, L.; Summers, J.D. Broiler response to energy or energy and protein dilution in the finisher diet. Poult. Sci. 1996, 75, 522–528. [Google Scholar] [CrossRef]
- Bragg, L.E.; Thompson, J.S.; West, W.W. Intestinal diamine oxidase levels reflect ischemic injury. J. Surg. Res. 1991, 50, 228–233. [Google Scholar] [CrossRef]
- Guo, Y.; Li, H.; Liu, Z.; Li, C.; Chen, Y.; Jiang, C.; Yu, Y.; Tian, Z. Impaired intestinal barrier function in a mouse model of hyperuricemia. Mol. Med. Rep. 2019, 20, 3292–3300. [Google Scholar] [CrossRef] [Green Version]
- Lv, Z.; Dai, H.; Wei, Q.; Jin, S.; Wang, J.; Wei, X.; Yuan, Y.; Yu, D.; Shi, F. Dietary genistein supplementation protects against lipopolysaccharide-induced intestinal injury through altering transcriptomic profile. Poult. Sci. 2020, 99, 3411–3427. [Google Scholar] [CrossRef]
- Cucco, M.; Guasco, B.; Malacarne, G.; Ottonelli, R. Effects of beta-carotene supplementation on chick growth, immune status and behaviour in the grey partridge, Perdix perdix. Behav. Process. 2006, 73, 325–332. [Google Scholar] [CrossRef]
- Dokladny, K.; Zuhl, M.N.; Moseley, P.L. Intestinal epithelial barrier function and tight junction proteins with heat and exercise. J. Appl. Physiol. 2016, 120, 692–701. [Google Scholar] [CrossRef]
- Liu, C.; Chen, C.; Yang, F.; Li, X.; Cheng, L.; Song, Y. Phytic acid improves intestinal mucosal barrier damage and reduces serum levels of proinflammatory cytokines in a 1,2-dimethylhydrazine-induced rat colorectal cancer model. Br. J. Nutr. 2018, 120, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Amiri, N.; Afsharmanesh, M.; Salarmoini, M.; Meimandipour, A.; Hosseini, S.A.; Ebrahimnejad, H. Nanoencapsulation (in vitro and in vivo) as an efficient technology to boost the potential of garlic essential oil as alternatives for antibiotics in broiler nutrition. Animal 2021, 15, 100022. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Items (%, Unless Otherwise Indicated) | Starter Period (Days 0 to 21) | Grower Period (Days 22 to 42) |
|---|---|---|
| Ingredient | ||
| Corn | 51.59 | 57.68 |
| Soybean meal | 40.78 | 35.15 |
| Soybean oil | 3.44 | 3.66 |
| Dicalcium phosphate | 1.92 | 1.33 |
| Limestone | 1.16 | 1.26 |
| Sodium chloride | 0.35 | 0.35 |
| DL-methionine | 0.26 | 0.13 |
| Choline chloride | 0.25 | 0.20 |
| Trance mineral premix 1 | 0.20 | 0.20 |
| Vitamin premix 2 | 0.05 | 0.04 |
| Calculated nutrient levels | ||
| Crude protein | 21.50 | 19.50 |
| ME (MJ/kg) | 12.22 | 12.56 |
| Ca | 1.00 | 1.00 |
| Available P | 0.45 | 0.35 |
| Lysine | 1.17 | 1.04 |
| Methionine + cystine | 0.90 | 0.72 |
| Gene Name | Primer Sequence (5′ to 3′) | NCBI Number | |
|---|---|---|---|
| β-actin | Forward | ACTCTGGTGATGGTGTTAC | NM 205518 |
| Reverse | GGCTGTGATCTCCTTCTG | ||
| Nrf-2 | Forward | ATCACCTCTTCTGCACCGAA | NM 205117 |
| Reverse | GCTTTCTCCCGCTCTTTCTG | ||
| SOD-1 | Forward | GGTGCTCACTTTAATCCTG | NM 205064 |
| Reverse | CTACTTCTGCCACTCCTCC | ||
| CAT | Forward | GGTTCGGTGGGGTTGTCTTT | NM_001031215.2 |
| Reverse | CACCAGTGGTCAAGGCATCT | ||
| GSH-Px1 | Forward | GACCAACCCGCAGTACATCA | NM_001277853.2 |
| Reverse | GAGGTGCGGGCTTTCCTTTA | ||
| GSH-Px3 | Forward | AAGTGCCAGGTGAACGGGAAGG | NM 001163232 |
| Reverse | AGGGCTGTAGCGGCGGAAAG | ||
| Claudin-1 | Forward | CATACTCCTGGGTCTGGTTGGT | AY750897 |
| Reverse | GACAGCCATCCGCATCTTCT | ||
| Occludin | Forward | ACGGCAGCACCTACCTCAA | D21837.1 |
| Reverse | GGGCGAAGAAGCAGATGAG | ||
| ZO-1 | Forward | CTTCAGGTGTTTCTCTTCCTCCTC | XM_413773 |
| Reverse | CTGTGGTTTCATGGCTGGATC | ||
| Mucin-2 | Forward | TTCATGATGCCTGCTCTTGTG | XM_421035 |
| Reverse | CCTGAGCCTTGGTACATTCTTGT |
| Item | Days 14 to 20 | Days 21 to 42 | |||||
|---|---|---|---|---|---|---|---|
| BWG (g) | FI (g) | FCR | BWG (g) | FI (g) | FCR | ||
| CTR | 176.2 | 302.3 | 1.755 | 1307.6 | 1966.4 | 1.506 | |
| CCP | 236.4 | 368.6 | 1.558 | 1391.8 | 2125.2 | 1.529 | |
| VA | 186.9 | 289.1 | 1.582 | 1306 | 1878.3 | 1.444 | |
| VA × CCP | 240.1 | 432.6 | 1.798 | 1388.3 | 2019.1 | 1.456 | |
| SEM | 7.20 | 16.60 | 0.07 | 76.00 | 32.50 | 0.02 | |
| Main effect | VA (−) | 181.2 b | 296.2 b | 1.675 | 1306.8 | 1925.7 | 1.478 |
| VA (+) | 238.2 a | 400.6 a | 1.678 | 1390 | 2072.1 | 1.492 | |
| CCP (−) | 206.3 | 335.5 | 1.656 | 1349.7 | 2045.8 a | 1.517 | |
| CCP (+) | 215.6 | 366.4 | 1.698 | 1350.3 | 1954.1 b | 1.45 | |
| p-Value | VA | 0.008 | 0.001 | 0.947 | 0.798 | 0.531 | 0.741 |
| CCP | 0.481 | 0.337 | 0.818 | 0.542 | 0.020 | 0.193 | |
| VA × CCP | 0.704 | 0.150 | 0.167 | 0.856 | 0.988 | 0.912 | |
| Item | d 28 | d 35 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CAT | H2O2 | MDA | GSH-px | T-SOD | CAT | H2O2 | MDA | GSH-px | T-SOD | ||
| U/mg Prot | nmol/mg Prot | U/mg Prot | U/mg Prot | nmol/mg Prot | U/mg Prot | ||||||
| CTR | 1.30 | 40.94 | 2.13 | 1493.92 | 90.57 | 1.17 | 24.47 | 2.76 | 1543.70 | 101.59 | |
| CCP | 1.10 | 34.23 | 2.17 | 1398.13 | 98.83 | 0.67 | 23.00 | 3.18 | 1641.23 | 102.87 | |
| VA | 0.78 | 37.48 | 2.72 | 1556.61 | 96.03 | 1.07 | 17.63 | 3.12 | 1544.98 | 105.19 | |
| VA × CCP | 0.77 | 38.29 | 2.76 | 1461.46 | 108.17 | 0.39 | 19.03 | 3.30 | 1493.25 | 103.79 | |
| SEM | 0.69 | 12.12 | 0.70 | 208.28 | 13.79 | 0.58 | 5.06 | 0.54 | 361.49 | 11.19 | |
| Main effect | VA (−) | 1.20 | 37.59 | 2.15 a | 1446.02 | 94.70 | 0.92 | 23.74 a | 2.97 | 1592.47 | 102.23 |
| VA (+) | 0.77 | 37.88 | 2.74 b | 1509.03 | 102.10 | 0.73 | 18.33 b | 3.21 | 1519.11 | 104.49 | |
| CCP (−) | 1.04 | 39.21 | 2.42 | 1525.26 | 93.30 b | 1.12 a | 21.05 | 2.94 | 1544.34 | 103.39 | |
| CCP (+) | 0.93 | 36.26 | 2.47 | 1429.79 | 103.50 a | 0.53 b | 21.02 | 3.24 | 1567.24 | 103.33 | |
| p-Value | VA | 0.090 | 0.928 | 0.018 | 0.345 | 0.073 | 0.286 | <0.001 | 0.158 | 0.537 | 0.541 |
| CCP | 0.671 | 0.371 | 0.855 | 0.156 | 0.015 | 0.003 | 0.981 | 0.083 | 0.847 | 0.987 | |
| VA × CCP | 0.700 | 0.255 | 0.989 | 0.996 | 0.631 | 0.607 | 0.306 | 0.478 | 0.530 | 0.715 | |
| Item | d 28 | d 35 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T-AOC | CAT | H2O2 | MDA | GSH-px | T-SOD | T-AOC | CAT | H2O2 | MDA | GSH-px | T-SOD | ||
| mM | U/mg Prot | nmol/mg Prot | U/mg Prot | mM | U/mg Prot | nmol/mg Prot | U/mg Prot | ||||||
| CTR | 1.18 b | 5.69 | 33.55 | 1.01 | 187.43 | 1187.10 | 1.08 | 10.48 | 48.40 | 1.11 | 189.97 | 1284.11 b | |
| CCP | 1.43 a | 5.43 | 31.03 | 1.29 | 147.73 | 1223.18 | 1.04 | 10.11 | 43.97 | 1.03 | 187.12 | 1452.47 a | |
| VA | 1.41 a | 5.06 | 44.77 | 1.00 | 145.66 | 1210.10 | 0.95 | 10.50 | 46.48 | 1.25 | 219.30 | 1123.94 c | |
| VA × CCP | 1.44 a | 4.71 | 41.56 | 1.10 | 115.65 | 1195.11 | 0.90 | 7.97 | 42.56 | 1.16 | 216.90 | 1088.80 c | |
| SEM | 0.14 | 1.76 | 7.86 | 0.30 | 41.83 | 105.34 | 0.16 | 2.67 | 12.39 | 0.27 | 35.79 | 198.04 | |
| Main effect | VA (−) | 1.30 b | 5.56 | 32.29 b | 1.15 | 167.58 a | 1205.14 | 1.06 a | 10.29 | 46.18 | 1.07 | 188.55 b | 1368.29 a |
| VA (+) | 1.43 a | 4.88 | 43.16 a | 1.05 | 130.65 b | 1202.61 | 0.93 b | 9.24 | 44.52 | 1.20 | 218.10 a | 1106.37 b | |
| CCP (−) | 1.30 b | 5.37 | 39.16 | 1.01 b | 166.54 a | 1198.60 | 1.01 | 10.49 | 47.44 | 1.18 | 204.64 | 1204.03 | |
| CCP (+) | 1.43 a | 5.07 | 36.30 | 1.20 a | 131.69 b | 1209.15 | 0.97 | 9.04 | 43.26 | 1.09 | 202.01 | 1270.63 | |
| p-Value | VA | <0.001 | 0.161 | <0.001 | 0.173 | <0.001 | 0.930 | 0.007 | 0.216 | 0.670 | 0.111 | 0.007 | <0.001 |
| CCP | <0.001 | 0.525 | 0.061 | 0.017 | 0.001 | 0.715 | 0.327 | 0.092 | 0.289 | 0.271 | 0.801 | 0.127 | |
| VA × CCP | <0.001 | 0.931 | 0.817 | 0.261 | 0.611 | 0.378 | 0.880 | 0.206 | 0.948 | 0.935 | 0.983 | 0.022 | |
| Item | d 28 | d 35 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T-AOC | CAT | H2O2 | MDA | GSH-px | T-SOD | T-AOC | CAT | H2O2 | MDA | GSH-px | T-SOD | ||
| mM | U/mg Prot | nmol/mg Prot | U/mg Prot | mM | U/mg Prot | nmol/mg Prot | U/mg Prot | ||||||
| CTR | 0.87 | 1.62 | 4.74 a | 1.92 b | 41.60 | 1274.41 | 0.45 | 1.51 | 3.89 | 0.68 | 21.32 | 1166.59 | |
| CCP | 0.86 | 0.98 | 2.93 b | 11.31 a | 38.65 | 997.07 | 0.53 | 1.35 | 4.86 | 0.52 | 21.08 | 789.92 | |
| VA | 0.38 | 1.25 | 4.42 a | 2.52 b | 24.80 | 953.01 | 0.53 | 1.31 | 3.24 | 0.38 | 18.05 | 849.46 | |
| VA × CCP | 0.37 | 1.28 | 4.54 a | 1.05 b | 26.71 | 779.95 | 0.57 | 0.63 | 3.34 | 0.45 | 20.79 | 555.99 | |
| SEM | 0.26 | 0.85 | 1.37 | 4.78 | 11.54 | 323.98 | 0.10 | 0.71 | 1.21 | 0.29 | 8.61 | 313.54 | |
| Main effect | VA (−) | 0.86 a | 1.30 | 3.83 b | 6.61 a | 40.12 a | 1135.74 a | 0.49 b | 1.43 a | 4.37 a | 0.60 a | 21.20 | 978.25 a |
| VA (+) | 0.38 b | 1.26 | 4.48 a | 1.79 b | 25.75 b | 866.48 b | 0.55 a | 0.97 b | 3.29 b | 0.42 b | 19.42 | 702.72 b | |
| CCP (−) | 0.62 | 1.44 | 4.58 a | 2.22 b | 33.20 | 1113.71 a | 0.49 b | 1.41 a | 3.57 | 0.53 | 19.68 | 1008.02 a | |
| CCP (+) | 0.61 | 1.13 | 3.73 b | 6.18 a | 32.68 | 888.51 b | 0.55 a | 0.99 b | 4.10 | 0.49 | 20.94 | 672.95 b | |
| p-Value | VA | <0.001 | 0.876 | 0.049 | <0.001 | <0.001 | 0.001 | 0.012 | 0.010 | <0.001 | 0.020 | 0.461 | <0.001 |
| CCP | 0.696 | 0.187 | 0.011 | <0.001 | 0.841 | 0.004 | 0.017 | 0.020 | 0.065 | 0.534 | 0.602 | <0.001 | |
| VA × CCP | 0.887 | 0.153 | 0.004 | <0.001 | 0.346 | 0.492 | 0.548 | 0.140 | 0.129 | 0.119 | 0.535 | 0.501 | |
| Item | d 28 | d 35 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Nrf-2 | SOD1 | CAT | GSH-px1 | GSH-px3 | Nrf-2 | SOD1 | CAT | GSH-px1 | GSH-px3 | ||
| CTR | 1.03 | 1.03 | 1.04 | 1.02 b | 1.06 a | 1.12 | 1.05 b | 1.04 | 1.01 | 1.05 | |
| CCP | 1.39 | 1.20 | 1.11 | 1.00 b | 1.01 a | 0.90 | 1.61 a | 1.75 | 1.33 | 0.80 | |
| VA | 1.62 | 1.19 | 1.33 | 0.57 c | 0.42 b | 1.11 | 0.81 b | 0.99 | 0.68 | 0.82 | |
| VA × CCP | 1.62 | 1.40 | 1.34 | 1.47 a | 1.33 a | 1.32 | 0.91 b | 1.30 | 0.81 | 0.63 | |
| SEM | 0.50 | 0.41 | 0.43 | 0.46 | 0.61 | 0.51 | 0.41 | 0.58 | 0.36 | 0.29 | |
| Main effect | VA (−) | 1.21 b | 1.11 | 1.07 | 1.01 | 1.04 | 1.01 | 1.33 a | 1.40 | 1.17 a | 0.92 a |
| VA (+) | 1.62 a | 1.30 | 1.34 | 1.02 | 0.88 | 1.22 | 0.86 b | 1.14 | 0.75 b | 0.72 b | |
| CCP (−) | 1.33 | 1.11 | 1.18 | 0.79 b | 0.74 b | 1.12 | 0.93 b | 1.02 b | 0.85 b | 0.93 a | |
| CCP (+) | 1.51 | 1.30 | 1.23 | 1.23 a | 1.17 a | 1.11 | 1.26 a | 1.53 a | 1.07 a | 0.71 b | |
| p-Value | VA | 0.008 | 0.156 | 0.054 | 0.917 | 0.350 | 0.242 | <0.001 | 0.120 | <0.001 | 0.045 |
| CCP | 0.225 | 0.142 | 0.738 | <0.001 | 0.018 | 0.968 | 0.002 | 0.003 | 0.038 | 0.026 | |
| VA × CCP | 0.228 | 0.878 | 0.842 | <0.001 | 0.009 | 0.228 | 0.022 | 0.225 | 0.338 | 0.753 | |
| Item | d 28 | d 35 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Nrf-2 | SOD1 | CAT | GSH-px1 | GSH-px3 | Nrf-2 | SOD1 | CAT | GSH-px1 | GSH-px3 | ||
| CTR | 1.07 | 1.02 | 1.03 | 1.01 b | 1.00 | 1.09 | 1.03 b | 1.04 c | 1.04 b | 1.06 b | |
| CCP | 0.89 | 1.29 | 0.91 | 0.90 b | 0.99 | 2.19 | 1.42 a | 1.76 a | 2.14 a | 1.72 a | |
| VA | 0.51 | 0.91 | 0.76 | 1.00 b | 1.44 | 1.03 | 1.04 bc | 1.84 ab | 1.36 b | 0.69 bc | |
| VA × CCP | 0.65 | 1.15 | 0.87 | 1.40 a | 1.46 | 1.22 | 0.84 c | 1.30 bc | 1.33 b | 0.60 c | |
| SEM | 0.43 | 0.34 | 0.28 | 0.34 | 0.50 | 0.89 | 0.36 | 0.55 | 0.62 | 0.63 | |
| Main effect | VA (−) | 0.98 a | 1.15 | 0.97 | 0.95 b | 0.99 b | 1.64 | 1.22 a | 1.40 | 1.59 | 1.39 a |
| VA (+) | 0.58 b | 1.03 | 0.81 | 1.20 a | 1.45 a | 1.13 | 0.94 b | 1.57 | 1.35 | 0.64 b | |
| CCP (−) | 0.79 | 0.97 b | 0.89 | 1.00 | 1.22 | 1.06 b | 1.03 | 1.44 | 1.20 a | 0.87 | |
| CCP (+) | 0.77 | 1.22 a | 0.89 | 1.15 | 1.22 | 1.70 a | 1.13 | 1.53 | 1.74 b | 1.16 | |
| p-Value | VA | 0.004 | 0.246 | 0.094 | 0.014 | 0.003 | 0.053 | 0.004 | 0.241 | 0.128 | <0.001 |
| CCP | 0.852 | 0.02 | 0.956 | 0.134 | 0.987 | 0.016 | 0.327 | 0.563 | 0.002 | 0.058 | |
| VA × CCP | 0.212 | 0.855 | 0.219 | 0.010 | 0.892 | 0.08 | 0.004 | <0.001 | 0.001 | 0.016 | |
| Item | Duodenum | Jejunum | Ileum | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| VH (μm) | CD (μm) | VH/CD | VH (μm) | CD (μm) | VH/CD | VH (μm) | CD (μm) | VH/CD | ||
| CTR | 864.02 | 93.49 | 9.57 | 490.22 | 64.89 | 7.65 | 274.11 | 60.23 | 4.55 | |
| CCP | 930.27 | 101.32 | 9.89 | 527.62 | 69.75 | 7.64 | 303.19 | 74.37 | 4.40 | |
| VA | 969.98 | 93.74 | 10.44 | 596.62 | 83.54 | 7.95 | 406.83 | 66.43 | 6.00 | |
| VA × CCP | 974.72 | 94.76 | 10.18 | 648.38 | 92.93 | 7.69 | 345.28 | 67.63 | 5.11 | |
| SEM | 101.39 | 16.56 | 1.66 | 135.07 | 32.05 | 1.81 | 146.08 | 21.27 | 1.51 | |
| Main effect | VA (−) | 897.14 b | 97.40 | 9.73 | 508.92 b | 67.32 b | 7.64 | 288.65 b | 67.30 | 4.47 b |
| VA (+) | 972.35 a | 94.25 | 10.31 | 622.50 a | 88.24 a | 7.82 | 376.05 a | 67.03 | 5.55 a | |
| CCP (−) | 917.00 | 93.61 | 10.00 | 543.42 | 74.22 | 7.80 | 340.47 | 63.33 | 5.27 | |
| CCP (+) | 952.49 | 98.04 | 10.03 | 588.00 | 81.34 | 7.66 | 324.23 | 71.00 | 4.75 | |
| p-Value | VA | 0.017 | 0.505 | 0.220 | 0.002 | 0.019 | 0.727 | 0.037 | 0.965 | 0.011 |
| CCP | 0.243 | 0.350 | 0.951 | 0.203 | 0.412 | 0.794 | 0.692 | 0.216 | 0.204 | |
| VA × CCP | 0.311 | 0.472 | 0.545 | 0.836 | 0.793 | 0.807 | 0.271 | 0.296 | 0.368 | |
| Item | Duodenum | Jejunum | Ileum | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| VH (μm) | CD (μm) | VH/CD | VH (μm) | CD (μm) | VH/CD | VH (μm) | CD (μm) | VH/CD | ||
| CTR | 819.86 | 103.85 | 8.21 | 719.49 | 88.15 | 8.58 | 519.11 | 96.53 | 5.45 | |
| CCP | 846.28 | 103.35 | 8.47 | 791.60 | 91.35 | 9.49 | 568.82 | 94.48 | 6.09 | |
| VA | 902.61 | 96.79 | 9.78 | 713.61 | 90.07 | 8.13 | 762.75 | 96.45 | 8.38 | |
| VA × CCP | 818.88 | 93.92 | 9.18 | 686.29 | 83.19 | 8.52 | 706.42 | 109.51 | 6.51 | |
| SEM | 141.00 | 22.22 | 2.32 | 110.45 | 15.23 | 1.45 | 328.89 | 18.78 | 3.57 | |
| Main effect | VA (−) | 833.07 | 103.60 | 8.34 | 755.55 | 89.75 | 9.04 | 543.97 b | 95.51 | 5.77 |
| VA (+) | 860.74 | 95.36 | 9.48 | 699.95 | 86.63 | 8.33 | 734.58 a | 102.98 | 7.44 | |
| CCP (−) | 861.23 | 100.32 | 9.00 | 716.55 | 89.11 | 8.36 | 640.93 | 96.49 | 6.91 | |
| CCP (+) | 832.58 | 98.63 | 8.83 | 738.95 | 87.27 | 9.01 | 637.62 | 102.00 | 6.30 | |
| p-Value | VA | 0.499 | 0.211 | 0.094 | 0.112 | 0.477 | 0.080 | 0.048 | 0.165 | 0.106 |
| CCP | 0.484 | 0.796 | 0.796 | 0.516 | 0.676 | 0.108 | 0.972 | 0.304 | 0.550 | |
| VA × CCP | 0.182 | 0.856 | 0.517 | 0.154 | 0.253 | 0.523 | 0.574 | 0.161 | 0.222 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Liu, C.; Niu, J.; Zhang, Y.; Li, C.; Zhang, Z.; Guo, S.; Ding, B. Effects of Dietary Supplementation with Vitamin A on Antioxidant and Intestinal Barrier Function of Broilers Co-Infected with Coccidia and Clostridium perfringens. Animals 2022, 12, 3431. https://doi.org/10.3390/ani12233431
Li P, Liu C, Niu J, Zhang Y, Li C, Zhang Z, Guo S, Ding B. Effects of Dietary Supplementation with Vitamin A on Antioxidant and Intestinal Barrier Function of Broilers Co-Infected with Coccidia and Clostridium perfringens. Animals. 2022; 12(23):3431. https://doi.org/10.3390/ani12233431
Chicago/Turabian StyleLi, Peng, Chengao Liu, Junlong Niu, Yuanke Zhang, Changwu Li, Zhengfan Zhang, Shuangshuang Guo, and Bingying Ding. 2022. "Effects of Dietary Supplementation with Vitamin A on Antioxidant and Intestinal Barrier Function of Broilers Co-Infected with Coccidia and Clostridium perfringens" Animals 12, no. 23: 3431. https://doi.org/10.3390/ani12233431