
Review of the Foundational Knowledge Required for Assessing Horse Welfare

Abstract
:Simple Summary
Abstract
1. Introduction
2. Domain 1: Nutritionü
2.1. Water Requirements
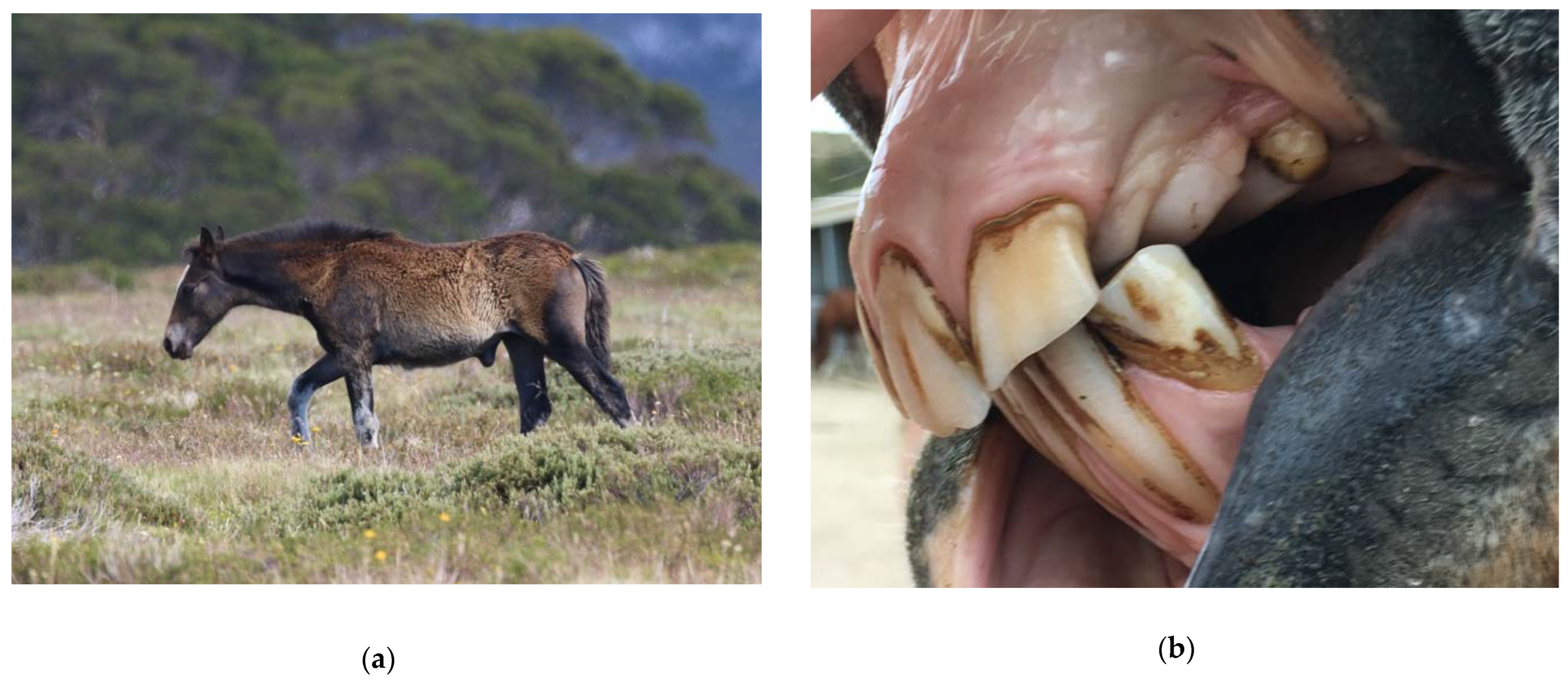
2.1.1. Factors Influencing Water Requirements
2.1.2. Adaptations to and Impacts of Water Restriction
2.2. Nutritional Requirements and Preferences
2.3. Common Nutritional Deficiencies and Excesses and Their Causes
2.4. Plant Toxicities
2.5. Body Condition Scoring
2.5.1. Optimal Body Condition Score
2.5.2. Factors Affecting Body Condition
3. Domain 2: Physical Environment
3.1. Habitat Preference and Factors Affecting Habitat Use
3.2. Preferred Underfoot Substrate and Terrain
3.3. Thermoneutral Zone
3.4. Thermoregulation and Factors Affecting It
3.5. Impacts of Ambient Cold
3.6. Impacts of Ambient Heat
3.7. Impacts of Extreme Climatic Events
4. Domain 3: Health
4.1. Common Non-Infectious Diseases
4.2. Common Infectious Diseases
4.3. Common Injuries
4.4. Sickness and Pain Behaviours
5. Domain 4: Behavioural Interactions
5.1. Social Organisation and Factors Affecting It
5.1.1. Social Structure
5.1.2. Factors Influencing Band and Herd Size
5.1.3. Band Composition
5.1.4. Factors Influencing Social Stability
5.1.5. Band Stallions
5.2. Population Dynamics
5.2.1. Impact of Food Resources
5.2.2. Birth and Survival Rates
5.2.3. Population Growth
5.2.4. Adult Mortality
5.2.5. Foal Mortality
5.2.6. Other Factors Influencing Population Growth Rates
5.3. Reproductive Physiology and Behaviours
5.4. Usual Range of Behaviours and Time Budgets
5.5. Social Behaviour and Communication
5.5.1. Affiliative Behaviours
5.5.2. Play Behaviour
5.5.3. Agonistic Behaviours
5.5.4. Visual, Auditory and Olfactory Communication
6. Concluding Comments
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mellor, D.J. Updating animal welfare thinking: Moving beyond the “Five Freedoms” towards “A Life worth Living”. Animals 2016, 6, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellor, D.J. Operational details of the Five Domains Model and its key applications to the assessment and management of animal welfare. Animals 2017, 7, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellor, D.J.; Beausoleil, N.J.; Littlewood, K.E.; McLean, A.N.; McGreevy, P.D.; Jones, B.; Wilkins, C. The 2020 Five Domains Model: Including Human–Animal Interactions in Assessments of Animal Welfare. Animals 2020, 10, 1870. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.M.; Beausoleil, N.J.; Ramp, D.; Mellor, D.J. A Ten-Stage Protocol for Assessing the Welfare of Individual Non-Captive Wild Animals: Free-Roaming Horses (Equus Ferus Caballus) as an Example. Animals 2020, 10, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cymbaluk, N.F. Chapter 4: Water, Equine Applied and Clinical Nutrition. In Equine Applied and Clinical Nutrition; Geor, R.J., Harris, P.A., Coenen, M., Eds.; Saunders Elsevier: London, UK, 2013; pp. 80–95. [Google Scholar]
- Sufit, E.; Houpt, K.A.; Sweeting, M. Physiological stimuli of thirst and drinking patterns in ponies. Equine Vet. J. 1985, 17, 12–16. [Google Scholar] [CrossRef]
- McDonnell, S.M.; Freeman, D.A.; Cymbaluk, N.F. Behaviour of stabled horses provided continuous or intermittent access to drinking water. Am. J. Vet. Res. 1999, 60, 1451–1456. [Google Scholar]
- Nyman, S.; Dahlborn, K. Effects of water supply method and flow rate on drinking behaviour and fluid balance in horses. Physiol. Behav. 2001, 73, 1–8. [Google Scholar] [CrossRef]
- Pellegrini, S. Home Range, Territoriality and Movement Patterns of Wild Horses in the Wassuk Range of Western Nevada. Ph.D. Thesis, University of Nevada, Reno, NV, USA, 1971. [Google Scholar]
- Feist, J.D.; McCullough, D.R. Behaviour patterns and communications in feral horses. Z. Tierpsy-Chologies 1976, 41, 337–371. [Google Scholar] [CrossRef]
- Scheibe, K.M.; Eichorn, K.; Kalz, B.; Striech, W.J.; Schiebe, A. Water consumption and watering behaviour of Przewalski mares in a semireserve. Zoo Biol. 1998, 17, 181–192. [Google Scholar] [CrossRef]
- Ganskopp, D.; Vavra, M. Habitat use of feral horses in the northern sagebrush steppe. J. Range Manag. 1986, 39, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; He, Z.Q.; Wang, H.; Niu, Y. Study on survival status of reintroduced Przewalskii in Dunhuang West Lake National Nature Reserve. J. Gansu For. Sci. Technol. 2012, 37, 45–46, 65. [Google Scholar]
- Berman, D.M. The Ecology of Feral Horses in Central Australia. Ph.D. Thesis, University of New England, Armidale, NSW, Australia, 1991. [Google Scholar]
- Hampson, B.A.; de Laat, M.A.; Mills, P.C.; Pollitt, C.C. Distances travelled by feral horses in ‘outback’ Australia. Equine Vet. J. 2010, 42, 582–586. [Google Scholar] [CrossRef] [PubMed]
- Hampson, B.; Morton, J.; Mills, P.; Trotter, M.; Lamb, D.; Pollitt, C. Monitoring distances travelled by horses using GPS tracking collars. Aust. Vet. J. 2010, 88, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Chen, J. Utilization of Food, Water and Space by Released Przewalski Horse with Reference to Survival Strategies Analysis. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2008. [Google Scholar]
- Zhang, F. The Study of Behaviour Time Budget and Its Influential Factors of Przewalski’s Horse. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2010. [Google Scholar]
- Schoenecker, K.A.; King, S.R.B.; Nordquist, M.K.; Nandintsetseg, D.; Cao, Q. Chapter 4: Habitat and diet of equids. In Wild Equids Ecology, Management and Conservation; Ransom, J.I., Kaczensky, P., Eds.; John Hopkins University Press: Baltimore, MD, USA, 2016; pp. 41–57. [Google Scholar]
- Krueger, K. Social ecology of horses. In Ecology of Social Evolution; Korb, J., Heinze, J., Eds.; Springer Science + Business Media: Berlin/Heidelberg, Germany, 2008; pp. 195–206. [Google Scholar] [CrossRef]
- Morgan, K.; Ehrlemark, A.; Sallvik, K. Dissipation of heat from standing horses exposed to ambient temperature between −3 and 37 °C. J. Therm. Biol. 1997, 22, 177–186. [Google Scholar] [CrossRef]
- McCutcheon, L.J.; Geor, R.J.; Hare, M.J.; Ecker, G.L.; Lindinger, M.I. Sweating rate and sweat composition during exercise and recovery in ambient heat and humidity. Equine Vet. J. 1995, 27, 153–157. [Google Scholar] [CrossRef]
- Jansson, A.; Dahlborn, K. Effects of feeding frequency and voluntary salt intake on fluid and electrolyte regulation in athletic horses. J. App. Physiol. 1999, 86, 1610–1616. [Google Scholar] [CrossRef]
- Cymbaluk, N.F.; Christison, G.I. Environmental effects of thermoregulation and nutrition of horses. Vet. Clin. N. Am. Equine. Pract. 1990, 6, 355–372. [Google Scholar] [CrossRef]
- Geor, R.J.; McCutcheon, L.J.; Lindinger, M.I. Adaptations to daily exercise in hot and humid conditions in trained Thoroughbred horses. Equine Vet. J. 1996, 28 (Suppl. 22), 63–68. [Google Scholar] [CrossRef]
- Mejdell, C.M.; Simensen, E.; Boe, K.E. Is snow a sufficient source of water for horses kept outdoors in winter? A case report. Acta Vet. Scand. 2005, 46, 19–22. [Google Scholar] [CrossRef]
- Kaczensky, P.; Deslez, V.; Vetter, D.; Otgonbayar, H.; Walzer, C. Water use of Asiatic wild asses in the Mongolian Gobi. In Exploration into the Biological Resources of Mongolia; University of Nebraska: Lincoln, NE, USA, 2010; pp. 291–298. [Google Scholar]
- Houpt, K.A.; Eggleston, K.; Kunkle, K. Effect of water restriction on equine behaviour and physiology. Equine Vet. J. 2000, 32, 341–344. [Google Scholar] [CrossRef]
- Luthersson, N.; Nielsen, K.H.; Harris, P. Risk factors associated with equine gastric ulceration syndrome (EGUS) in 201 horses in Denmark. Equine Vet. J. 2009, 41, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Hampson, B.A.; Owens, E.; Watts, K.A.; Mills, P.C.; Pollitt, C.C.; de Laat, M.A. Nutritional analysis of gastric contents and body condition score at a single time point in feral horses in Australia. Am. J. Vet. Res. 2011, 72, 1226–1233. [Google Scholar] [CrossRef] [PubMed]
- Carlson, G.P.; Rumbaugh, G.E.; Harrold, D. Physiologic alterations in the horse produced by food and water deprivation during periods of high environmental temperatures. Am. J. Vet. Res. 1979, 40, 982–985. [Google Scholar] [PubMed]
- Hansen, R.M. Foods of free-roaming horses in southern New Mexico. J. Range Manag. 1976, 29, 347. [Google Scholar] [CrossRef]
- Krysl, L.J.; Hubbert, M.E.; Sowell, B.F.; Plumb, G.E.; Jewett, T.K.; Smith, M.A. Horses and cattle grazing in the Wyoming red desert. J. Range Manag. 1984, 37, 72–76. [Google Scholar] [CrossRef] [Green Version]
- Crane, K.K.; Smith, M.A.; Reynolds, D. Habitat selection patterns of feral horses in southcentral Wyoming. J. Range Manag. 1997, 35, 152–158. [Google Scholar] [CrossRef]
- Janis, C. 1976. The evolutionary strategy of the Equidae and the origins of rumen and caecal digestion. Evolution 1976, 30, 757–774. [Google Scholar] [CrossRef]
- Duncan, P. Horses and Grasses: The Nutritional Ecology of Equids and Their Impact on the Camargue; Springer: New York, NY, USA, 1992. [Google Scholar]
- Salter, R.E.; Hudson, R.J. Feeding ecology of feral horses in Western Alberta. J. Range Manag. 1979, 32, 221–225. [Google Scholar] [CrossRef] [Green Version]
- McInnis, M.L.; Vavra, M. Dietary relationships among feral horses, cattle and pronghorn in the southeastern Oregon. J. Range Manag. 1987, 40, 60–66. [Google Scholar] [CrossRef]
- Smith, C.; Valdex, R.; Holecheck, J.L.; Zwank, P.J.; Cardenas, M. Diets of native and non-native ungulates in southcentral New Mexico. Southeast. Nat. 1998, 43, 163–169. [Google Scholar]
- Hubbard, R.E.; Hansen, R.M. Diets of wild horses, cattle, and mule deer in the Piceance Basin, Colorado. J. Range Manag. 1976, 29, 389–392. [Google Scholar] [CrossRef]
- Hanley, T.A.; Hanley, K.A. Food resource partitioning by sympatric ungulates on Great Basin rangeland. J. Range Manag. 1982, 35, 152–158. [Google Scholar] [CrossRef]
- Gordon, I.J. Vegetation community selection by ungulates on the Isle of Rhum. III Determinants of vegetation community selection. J. Appl. Ecol. 1989, 26, 65–79. [Google Scholar] [CrossRef]
- Naujeck, A.; Hill, J.; Gibb, M.J. Influence of sward height on diet selection by horses. Appl. Anim. Behav. Sci. 2005, 90, 49–63. [Google Scholar] [CrossRef]
- Fleurance, G.; Fritz, H.; Duncan, P.; Gordon, I.J.; Edouard, N.; Vial, C. Instantaneous intake rate in horses of different body sizes: Influence of sward biomass and fibrousness. Appl. Anim. Behav. Sci. 2009, 117, 84–92. [Google Scholar] [CrossRef]
- Cairns, M.C.; Cooper, J.J.; Davidson, H.P.B. Association in horses of orosensory characteristics of foods with their post-ingestive consequences. Anim. Sci. 2002, 75, 257–265. [Google Scholar] [CrossRef]
- Vinassa, M.; Cavallini, D.; Galaverna, D.; Baragli, P.; Raspa, F.; Nery, J.; Valle, E. Palatability assessment in horses in relation to lateralization and temperament. Appl. Anim. Behav. Sci. 2020, 232, 105110. [Google Scholar] [CrossRef]
- van Wieren, S. Do large herbivores select a diet that maximizes short-term energy intake rate? For. Ecol. Manag. 1996, 88, 149–156. [Google Scholar] [CrossRef]
- Raspa, F.; Vervuert, I.; Capucchio, M.T.; Colombino, E.; Bergero, D.; Forte, C.; Greppi, M.; Cavallarin, L.; Giribaldi, M.; Antoniazzi, S.; et al. A high-starch vs. high-fibre diet: Effects on the gut environment of the different intestinal compartments of the horse digestive tract. BMC Vet. Res. 2022, 18, 187. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Horses, 6th ed.; The National Academies Press: Washington, DC, USA, 2007. [Google Scholar] [CrossRef]
- Kronfeld, D.S. Starvation and nutrition of horses: Recognition and treatment. Proceedings of the first conference on equine rescue. Santa Barbara, California, USA. J. Equine Vet. Sci. 1993, 13, 298–303. [Google Scholar] [CrossRef]
- Kohnke, J.K.; Kelleher, F.; Trevor-Jones, P. Feeding Horses in Australia; RIRDC Publication: Canberra, Australia, 1999.
- Floyd, A. Pathological conditions of the equine foot. In Equine Podiatry; Saunders Elsevier: St Louis, MO, USA, 2007. [Google Scholar]
- Olstad, K.; Ekman, S.; Carlson, C.S. An Update on the Pathogenesis of Osteochondrosis. Vet. Pathol. 2015, 52, 785–802. [Google Scholar] [CrossRef] [PubMed]
- McGreevy, P.D.; Hawson, L.A.; Habermann, T.C.; Cattle, S.R. Geophagia in horses: A short note on 13 cases. Appl. Anim. Behav. Sci. 2001, 71, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Aytekin, I.; Onmaz, A.C.; Aypak, S.U.; Gunes, V.; Kucuk, O. Changes in Serum Mineral Concentrations, Biochemical and Hematological Parameters in Horses with Pica. Biol. Trace. Elem. Res. 2011, 139, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Coenen, M. Chapter 10: Macro and trace elements in equine nutrition. Assessment of body condition and bodyweight. In Equine Applied and Clinical Nutrition; Geor, R.J., Harris, P.A., Coenen, M., Eds.; Saunders Elsevier: London, UK, 2013; pp. 190–228. [Google Scholar]
- Pagan, J.D. The role of nutrition in developmental orthopaedic disease: Nutritional management. In Diagnosis and Management of Lameness in the Horse; Ross, M.W., Dyson, S.J., Eds.; Saunders: St Louis, Mo, USA, 2003; pp. 203–213. [Google Scholar]
- Granot, N.; Milgram, J.; Bdolah-Abram, T.; Shemesh, I.; Steinman, A. Surgical management of sand colic impactions in horses: A retrospective study of 41 cases. Aust. Vet. J. 2008, 86, 404–407. [Google Scholar] [CrossRef]
- McKenzie, R. Australia’s Poisonous Plants, Fungi and Cyanobacteria: A Guide to Species of Medical and Veterinary Importance; CSIRO: Collingwood, Australia, 2012.
- Huntington, P.J.; Jeffcott, L.B.; Friend, S.C.E.; Luff, A.R.; Finkelstein, D.I.; Flynn, R.J. Australian Stringhalt—Epidemiological, clinical and neurological investigations. Equine Vet. J. 1989, 21, 266–273. [Google Scholar] [CrossRef]
- Bell, A.T.; Everist, S.L. Indigofera enneaphylla: A plant toxic to horses (Birdsville disease). Aust. Vet. J. 1951, 27, 185–188. [Google Scholar] [CrossRef]
- Severne, B.C.; Brooks, R.R. A nickel-accumulating plant from Western Australia. Planta 1972, 103, 112–116. [Google Scholar] [CrossRef]
- Severne, B. Nickel accumulation by Hybanthus floribundas. Nature 1974, 248, 807–808. [Google Scholar] [CrossRef]
- Carrol, C.; Huntington, P. Body condition scoring and weight estimation of horses. Equine Vet. J. 1988, 20, 41–45. [Google Scholar] [CrossRef]
- Henneke, D.R.; Potter, G.D.; Kreider, J.L.; Yeates, B.F. Relationship between condition score, physical measurements and body fat percentage in mares. Equine Vet. J. 1983, 15, 371–372. [Google Scholar] [CrossRef]
- Henneke, D.R.; Potter, G.D.; Kreiger, J.L. Body condition during pregnancy and lactation, and reproductive efficiency of mares. Theriogenology 1984, 21, 897–909. [Google Scholar] [CrossRef]
- Pritchard, J.C.; Lindberg, A.C.; Main, D.C.J.; Whay, H.R. Assessment of the welfare of working horses, mules and donkeys, using health and behaviour parameters. Prev. Vet. Med. 2005, 69, 265–283. [Google Scholar] [CrossRef]
- Burn, C.C.; Pritchard, J.; Whay, H. Observer reliability for working equine welfare assessment: Problems with high prevalences of certain results. Anim. Welf. 2009, 18, 177–187. [Google Scholar]
- Burn, C.C.; Dennison, T.L.; Whay, H.R. Relationships between behaviour and health in working horses, donkeys, and mules in developing countries. Appl. Anim. Behav. Sci. 2010, 126, 109–118. [Google Scholar] [CrossRef]
- Dawson, M.J.; Hone, J. Demography and dynamics of three wild horse populations in the Australian Alps. Austral Ecol. 2012, 37, 97–109. [Google Scholar] [CrossRef]
- Samuel, E.K.; Whay, H.R.; Mullan, S. A preliminary study investigating the physical welfare and welfare code compliance for tethered and free-ranging horses on common land in South Wales. Anim. Welf. 2012, 21, 593–598. [Google Scholar] [CrossRef] [Green Version]
- Hockenhull, J.; Whay, H.R. A review of approaches to assessing equine welfare. Equine Vet. Educ. 2014, 26, 159–166. [Google Scholar] [CrossRef]
- Mullan, S.; Szmaraged, C.; Hotchkiss, I.; Whay, H.R. The welfare of long-line tethered and free-ranging horses kept on public grazing land in South Wales. Anim. Welf. 2014, 23, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Czycholl, I.; Büttner, K.; Klingbeil, P.; Krieter, J. An Indication of Reliability of the Two-Level Approach of the AWIN Welfare Assessment Protocol for Horses. Animals 2018, 8, 7. [Google Scholar] [CrossRef] [Green Version]
- Somerville, R.; Brown, A.F.; Upjohn, M. A standardised equine-based assessment tool used for six years in low and middle income countries. PLoS ONE 2018, 13, e0192354. [Google Scholar] [CrossRef] [Green Version]
- Gentry, L.R.; Thompson, D.L., Jr.; Gentry, G.T., Jr.; Del Vecchio, R.P.; Davis, K.A.; Del Vecchio, P.M. The relationship between body condition score and ultrasonic fat measurements in mares of high versus low body condition. J. Equine Vet. Sci. 2004, 24, 198–203. [Google Scholar] [CrossRef]
- Cameron, E.Z.; Linklater, W.L.; Stafford, K.J.; Minot, E.O. Maternal investment results in better foal condition through increased play behaviour in horses. Anim. Behav. 2008, 76, 1511–1518. [Google Scholar] [CrossRef]
- Dugdale, A.H.A.; Grove-White, D.; Curtis, G.C.; Harris, P.A.; Argo, C.M. Body condition scoring as a predictor of body fat in horses and ponies. Vet. J. 2012, 194, 173–178. [Google Scholar] [CrossRef]
- Jensen, R.B.; Danielsen, S.H.; Tauson, A. Body condition score, morphometric measurements and estimation of body weight in mature Icelandic horses in Denmark. Acta Vet. Scand. 2016, 58, 50. [Google Scholar] [CrossRef] [Green Version]
- Carter, R.A.; Dugdale, A.H.A. Chapter 2: Assessment of body condition and bodyweight. In Equine Applied and Clinical Nutrition; Geor, R.J., Harris, P.A., Coenen, M., Eds.; Saunders Elsevier: London, UK, 2013; pp. 393–403. [Google Scholar]
- Gentry, L.R.; Thompson, D.L., Jr.; Gentry, G.T., Jr. The relationship between body condition, leptin and reproductive and hormonal characteristics of mares during the seasonal anovulatory period. J. Anim. Sci. 2002, 80, 2695–2703. [Google Scholar] [CrossRef]
- Lawrence, L.M. Chapter 11 Feeding stallions and broodmares. In Equine Applied and Clinical Nutrition; Geor, R.J., Harris, P.A., Coenen, M., Eds.; Saunders Elsevier: London, UK, 2013; pp. 231–242. [Google Scholar]
- Cameron, E.Z.; Linklater, W.L.; Minot, E.O.; Stafford, K.J.; Veltman, C.J. Birth sex ratios relate to mare condition at conception in Kaimanawa horses. Behav. Ecol. 1999, 10, 472–475. [Google Scholar] [CrossRef]
- Stanley, C.; Shultz, S. Mummy’s boys: Sex differential maternal-offspring bonds in semi-feral horses. Behaviour 2012, 149, 251–274. [Google Scholar] [CrossRef]
- Watts, E.T.; Johnson, C.N.; Carver, S.; Butler, C.; Harvey, A.M.; Cameron, E.Z. Maternal protectiveness in feral horses: Responses to intraspecific and interspecific sources of risk. Anim. Behav. 2020, 159, 1–11. [Google Scholar] [CrossRef]
- Lima, S.L.; Dill, L.M. Behavioural decisions made under the risk of predation. Can. J. Zool. 1990, 68, 619–640. [Google Scholar] [CrossRef]
- Grange, S.; Duncan, P.; Gaillard, J.M. Poor horse traders: Large mammals trade survival for reproduction during the process of feralisation. Proc. Royal Soc. B 2009, 276, 1911–1919. [Google Scholar] [CrossRef] [Green Version]
- Debeffe, L.; Mcloughlin, P.D.; Medill, S.A.; Stewart, K.; Andres, D.; Shury, T.; Wagner, B.; Jenkins, E.; Gilleard, J.S.; Poissant, J. Negative covariance between parasite load and body condition in a population of feral horses. Parasitology 2016, 143, 983–997. [Google Scholar] [CrossRef] [PubMed]
- Cain, J.L.; Jarisch, K.; Macaluso, K.; Luedtke, B.E. Correlation between fecal egg count, presence of Strongylus vulgaris, and body score of feral horses on Fort Polk, Louisiana. Vet. Parasitol. Reg. Stud. Rep. 2018, 13, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Scorolli, A.L. Feral Horse Population Model and Body Condition: Useful Management Tools in Tornquist Park, Argentina. J. Wildl. Manag. 2021, 85, 1162–1168. [Google Scholar] [CrossRef]
- King, S.R.B.; Gurnell, J. Habitat use and spatial dynamics of takhi introduced to Hustai National Park, Mongolia. Biol. Conserv. 2005, 124, 227–290. [Google Scholar] [CrossRef]
- Henley, S.R.; Ward, D.; Schmidt, I. Habitat selection by two desert-adapted ungulates. J. Arid Environ. 2007, 70, 39–48. [Google Scholar] [CrossRef]
- Kaczensky, P.; Ganbataar, O.; Wehrden, H.V.; Walzer, C. Resource selection by Sympatric wild equids in the Mongolian Gobi. J. Appl. Ecol. 2008, 45, 1762–1769. [Google Scholar] [CrossRef]
- Salter, R.E. Ecology of Feral Horses in Western Alberta. Ph.D. Thesis, University of Alberta, Edmonton, AB, Canada, 1978. [Google Scholar]
- Berger, J. Wild Horses of the Great Basin, Social Competition and Population Size; University of Chicago Press: Chicago, IL, USA, 1986. [Google Scholar]
- Girard, T.L.; Bork, E.W.; Nielson, S.E.; Alexander, M.J. Seasonal variation in habitat selection by free-ranging feral horses within Alberta’s forest reserve. Rangel. Ecol. Manag. 2013, 66, 428–437. [Google Scholar] [CrossRef]
- Linklater, W.; Cameron, E.; Stafford, K.; Minot, E. Social and spatial structure and range use by Kaimanawa wild horses (Equus caballus: Equidae. N. Z. J. Ecol. 2000, 24, 139–152. [Google Scholar]
- Walter, M. The population ecology of wild horses in the Australian Alps. Ph.D. Thesis, University of Canberra, Bruce, ACT, Australia, 2002. [Google Scholar]
- McCort, W.D. Behaviour of feral horses and ponies. J. Anim. Sci. 1984, 58, 493–499. [Google Scholar] [CrossRef]
- Salter, R.E.; Hudson, R.J. Social organization of feral horses in western Canada. Appl. Anim. Ethol. 1982, 8, 207–223. [Google Scholar] [CrossRef]
- Miller, R. Seasonal movement patterns and home ranges of feral horses in Wyoming’s Red Desert. J. Range Manag. 1983, 36, 199–201. [Google Scholar] [CrossRef] [Green Version]
- Zervanos, S.; Keiper, R.R. Seasonal home ranges and activity patterns of feral Assateague ponies. In Symposium on the Ecology and Behaviour of Wild and Feral Equids; Denniston, R.H., Ed.; University of Wyoming: Laramie, WY, USA, 1979; pp. 3–14. [Google Scholar]
- Keiper, R.R.; Keenan, M.A. Nocturnal activity patterns of feral ponies. J. Mammal. 1980, 61, 116–118. [Google Scholar] [CrossRef]
- Rutberg, A.T. Horse fly harassment and the social behaviour of feral ponies. Ethology 1987, 75, 145–154. [Google Scholar] [CrossRef]
- Powell, D.M.; Danze, D.E.; Gwinn, M.A. Predicators of biting fly harassment and its impact on habitat use by feral horses (Equus caballus) on a barrier island. J. Ethol. 2006, 24, 147–154. [Google Scholar] [CrossRef]
- King, S.R.B.; Gurnell, J. Effects of fly disturbance on the behaviour of a population of reintroduced Przewalski horses in Mongolia. Appl. Anim. Behav. Sci. 2010, 125, 22–29. [Google Scholar] [CrossRef]
- Hampson, B.A. The Effects of the Environment on the Feral Horse Foot. Ph.D. Thesis, University of Queensland, St Lucia, QLD, Australia, 2011. [Google Scholar]
- Hampson, B.A.; de Laat, M.A.; Mills, P.C.; Pollitt, C.C. The feral horse foot. Part A: Observational study of the effect of environment on the morphometrics of the feet of 100 Australian feral horses. Aust. Vet. J. 2013, 91, 14–22. [Google Scholar] [CrossRef]
- Cymbaluk, N.F. Thermoregulation of horses in cold, winter weather: A review. Livest. Prod. Sci. 1994, 40, 15–71. [Google Scholar] [CrossRef]
- Dawson, M. The population ecology of feral horses in the Australian Alps Management Summary. In Proceedings of the Australian Alps Liaison Committee, Canberra, Australia, April 2005. [Google Scholar]
- Morgan, K. Thermoneutral zone and critical temperatures of horses. J. Therm. Biol. 1998, 23, 59–61. [Google Scholar] [CrossRef]
- Guthrie, A.J.; Lund, R.J. Thermoregulation: Base mechanisms and hyperthermia. Vet. Clin. N. Am. Equine Pract. 1998, 14, 45–59. [Google Scholar] [CrossRef]
- Berger, J. Ecology and catastrophic mortality in wild horses: Implications for interpreting fossil assemblages. Science 1983, 220, 1403–1404. [Google Scholar] [CrossRef]
- Garrott, R.A.; Taylor, L. Dynamics of a feral horse population in Montana. J. Wildl. Manag. 1990, 54, 603–612. [Google Scholar] [CrossRef]
- Scorolli, A.; Lopez-Cazorla, A.C.; Tejera, L.A. Unusual mass mortality of feral horses during a violent rainstorm in Parque Provincial Tornquist, Argentina. Mastozool. Neotrop. 2006, 13, 255–258. [Google Scholar]
- Kaczensky, P.; Ganbataar, O.; Altansukh, N.; Ankhsaikhan, M.; Stauffer, C.; Walzer, C. The danger of having all your eggs in one basket—Winter crash of the re-introduced Przewalski’s horses in the Mongolian Gobi. PLoS ONE 2011, 6, e28057. [Google Scholar] [CrossRef] [PubMed]
- Kane, A.J. Chapter 24: The welfare of wild horses in the Western USA. In Equine Welfare; McIlwraith, C.W., Rollin, B.E., Eds.; UFAW Animal Welfare Series; Wiley-Blackwell: West Sussex, UK, 2011; pp. 442–462. [Google Scholar]
- Kane, A.J. Wild horse health on and off the range. Proc. Annu. Conv. Am. Assoc. Equine Pract. 2014. Available online: https://thehorse.com/111530/wild-horse-health-on-and-off-the-range/ (accessed on 19 November 2022).
- Nunez, C.M.V.; Scorolli, A.; Lagos, L.; Berman, D.; Kane, A.J. Chapter 10: Management of free-roaming horses. In Wild Equids Ecology, Management and Conservation; Ransom, J.I., Kaczensky, P., Eds.; John Hopkins University Press: Baltimore, MD, USA, 2016; pp. 133–148. [Google Scholar]
- Cantley, C.E.L.; Firth, E.C.; Delahunt, J.W.; Pfeiffer, D.U.; Thompson, K.G. Naturally occurring osteoarthritis in the metacarpophalangeal joints of wild horses. Equine Vet. J. 1999, 31, 73–81. [Google Scholar] [CrossRef]
- Górecka-Bruzda, A.; Jaworski, Z.; Jaworska, J.; Siemieniuch, M. Welfare of Free-Roaming Horses: 70 Years of Experience with Konik Polski Breeding in Poland. Animals 2020, 10, 1094. [Google Scholar] [CrossRef] [PubMed]
- Leblond, A.; Corbin-Gardey, C.; Cadore, J.L. Studies of the pathology and causes of mortality of the old horse. In Proceedings of the 41st British Equine Veterinary Congress, Equine Veterinary Journal, Newmarket, ON, USA, 2002; pp. 202–203. [Google Scholar]
- McGowan, T.W.; Pinchbeck, G.L.; Hodgson, D.R. Clinical disease and mortality in in geriatric horses: An Australian Perspective. In Proceedings of the Havemeyer Foundation Equine Geriatric Workshop; Dorothy Havemeyer Foundation: Cambridge, MA, USA, 2010; p. 23. [Google Scholar]
- Hampson, B.A.; Connelley, A.D.; de Laat, M.A.; Mills, P.C.; Pollitt, C.C. Sole depth and weight-bearing characteristics of the palmar surface of the feet of feral horses and domestic Thoroughbreds. Am. J. Vet. Res. 2011, 72, 727–735. [Google Scholar] [CrossRef]
- Hampson, B.A.; de Laat, M.A.; Mills, P.C.; Walsh, D.M.; Pollitt, C.C. The feral horse foot. Part B: Radiographic, gross visual and histopathological parameters of foot health in 100 Australian feral horses. Aust. Vet. J. 2013, 91, 23–30. [Google Scholar] [CrossRef]
- Hampson, B.A.; Ramsey, G.; Macintosh, A.M.H.; Mills, P.C.; de Laat, M.A.; Pollitt, C.C. Morphometry and abnormalities of the feet of Kaimanawa feral horses in New Zealand. Aust. Vet. J. 2010, 88, 124–131. [Google Scholar] [CrossRef]
- Hampson, B.A.; de Laat, M.A.; Beausac, C.; Rovel, T.; Pollitt, C.C. Histopathological examination of chronic laminitis in Kaimanawa feral horses of New Zealand. N. Z. Vet. J. 2012, 60, 285–289. [Google Scholar] [CrossRef]
- Eades, S.C. Overview of What We Know About the Pathophysiology of Laminitis. J. Equine Vet. Sci. 2010, 30, 83–86. [Google Scholar] [CrossRef]
- Ramey, P. Care and Rehabilitation of the Equine Foot; Hoof Rehabilitation Publishing, LLC.: Lakemont, GA, USA, 2011. [Google Scholar]
- Mellish, M.A.; Lucas, Z.N.; Puchalski, S.M.; Kusch, T.A. An Estimation of Lameness in Sable Island Horses Using Radiographic Evaluation of the Distal Phalanx and Hoof Capsule. J. Appl. An. Welf. Sci. 2021, 17, 1–11. [Google Scholar] [CrossRef]
- Niinistö, K.; Sykes, B.W. Diagnosis and management of sand enteropathy in the horse. Equine Vet. Educ. 2021, 34, 600–606. [Google Scholar] [CrossRef]
- Morley, S.; Murray, J. The effects of body condition on the reproductive physiology of the broodmare: A review. J. Equine Vet. Sci. 2014, 34, 842–853. [Google Scholar] [CrossRef]
- Schroeder, D.G.; Brown, K.H. Nutritional status as a predictor of child survival: Summarising the association and quantifying its global impact. Bull. World Health Organ. 1984, 2, 569–579. [Google Scholar]
- Naylor, J.M.; Kenyon, S.J. Effects of total caloric deprivation on host defence in the horse. Res. Vet. Sci. 1981, 31, 369–372. [Google Scholar] [CrossRef] [PubMed]
- McGregor-Argo, C. Chapter 29: Feeding thin and starved horses. In Equine Applied & Clinical Nutrition; Geor, R.J., Harris, P.A., Coenen, M., Eds.; Saunders Elsevier: London, UK, 2013; pp. 503–511. [Google Scholar]
- Shetty, P.S. Adaptation to low energy intakes: The responses and limits to low intakes in infants, children and adults. Eur. J. Clin. Nutr. 1999, 53, S14–S33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobbie, W.R.; Berman, D.; Braysher, M.L. Managing Vertebral Pests: Feral Horses; Australian Government Publishing Service: Canberra, Australia, 1993.
- Singer, F.J.; Zeigenfuss, L.; Coates-Markle, L.; Schweiger, F. A demographic analysis, group dynamics, and genetic effective number in the Pryor Mountain wild horse population. In Managers’ Summary—Ecological Studies of the Pryor Mountain Wild Horse Range, 1992–1997; Singer, F.J., Schoenecker, K.A., Eds.; US Geological Survey: Fort Collins, CO, USA, 2000; pp. 73–89. [Google Scholar]
- Da Silva, A.S.; Tonin, A.A.; Lopes, L.S. Outbreak of lice in horses: Epidemiology, diagnosis, and treatment. J. Equine Vet. Sci. 2013, 33, 530–532. [Google Scholar] [CrossRef]
- Kay, G.; Knottenbelt, D.C. Tetanus in equids: A report of 56 cases. Equine Vet. Educ. 2007, 19, 107–112. [Google Scholar] [CrossRef]
- Boyle, A.; Timoney, J.; Newton, J.; Hines, M.; Waller, A.; Buchanan, B. Streptococcus equi infections in horses: Guidelines for treatment, control, and prevention of strangles—revised consensus statement. J. Vet. Intern. Med. 2018, 32, 633–647. [Google Scholar] [CrossRef] [Green Version]
- Torres, S.M.; Koch, S.N. Papillomavirus-associated diseases. Vet. Clin. N. Am. Equine Pract. 2013, 29, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Lunn, D.P.; Davis-Poynter, N.; Flaminio, M.J.; Horohov, D.W.; Osterrieder, K.; Pusterla, N.; Townsend, H.G. Equine Herpesvirus-1 Consensus Statement. J. Vet. Intern. Med. 2009, 23, 450–461. [Google Scholar] [CrossRef] [PubMed]
- El-Hage, C.M.; Bamford, N.J.; Gilkerson, J.R.; Lynch, S.E. Ross River Virus Infection of Horses: Appraisal of Ecological and Clinical Consequences. J. Equine Vet. Sci. 2020, 93, 103143. [Google Scholar] [CrossRef]
- Middleton, D. Hendra virus. Vet. Clin. N. Am. Equine Pract. 2014, 30, 579–589. [Google Scholar] [CrossRef]
- Warrilow, D. Australian bat lyssavirus: A recently discovered new rhabdovirus. In The World of Rhabdoviruses; Fu, Z.F., Ed.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 24–44. [Google Scholar] [CrossRef]
- Dubey, J.P.; Mitchell, S.M.; Morrow, J.K.; Rhyan, J.C.; Stewart, L.M.; Granstrom, D.E.; Romand, S.; Thulliez, P.; Saville, W.J.; Lindsay, D.S. Prevalence of Antibodies to Neospora Caninum, Sarcocystis Neurona, and Toxoplasma Gondii in Wild Horses from Central Wyoming. J. Parasitol. 2003, 89, 716–720. [Google Scholar] [CrossRef]
- Mateu, M.E.; Finol, H.J.; Sucre, L.E.; Torres, S.H. Muscular changes in Venezuelan wild horses naturally infected with Trypanosoma evansi. J. Comp. Pathol. 1994, 110, 79–89. [Google Scholar] [CrossRef]
- Rubenstein, D.I.; Hohmann, M.E. Parasites and social behaviour of island feral horses. Oikos 1989, 55, 312–320. [Google Scholar] [CrossRef] [Green Version]
- Slivinska, K.; Dvojnos, G.; Kopij, G. Helminth fauna of sympatric Przwalski’s Equus przewalskii Poljav, 1881 and domestic horses E. caballus L. in the Chernobyl exclusion zone, Ukraine. Helminthologia 2006, 43, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Slivinska, K.; Gawoe, J.; Jaworski, Z. Gastro-intestinal parasite in yearlings of wild Polish primitive horses from the Popielno Forest Reserve, Poland. Helminthologia 2009, 46, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Harvey, A.M.; Meggiolaro, M.N.; Hall, E.; Watts, E.T.; Ramp, D.; Šlapeta, J. Wild horse populations in south-east Australia have a high prevalence of Strongylus vulgaris and may act as a reservoir of infection for domestic horses. Int. J. Parasitol. Parasites Wildl. 2019, 8, 156–163. [Google Scholar] [CrossRef]
- Slivinska, K.; Klich, D.; Yasynetska, N.; Żygowska, M. The effects of seasonality and group size on fecal egg counts in wild Przewalski’s horses (Poljakov, 1881) in the Chernobyl Exclusion Zone, Ukraine during 2014–2018. Helminthologia 2020, 57, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Clayton, H.; Duncan, J. Clinical signs associated with parascaris equorum infection in worm-free pony foals and yearlings. Vet. Parasitol. 1978, 4, 69–78. [Google Scholar] [CrossRef]
- Srihakim, S.; Swerczek, T.W. Pathological changes and pathogenesis of Parascaris equorum infection in parasite-free pony foals. Am. J. Vet. Res. 1978, 39, 1155–1160. [Google Scholar] [PubMed]
- Love, S.; Murphy, D.; Mellor, D. Pathogenicity of cyathostome infection. Vet. Parasitol. 1999, 85, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Peregrine, A.S.; McEwen, B.; Bienzle, D.; Koch, T.G.; Weese, J.S. Larval cyathostominosis in horses in Ontario: An emerging disease? Can. Vet. J. 2006, 47, 80–82. [Google Scholar]
- Corning, S. Equine cyathostomins: A review of biology, clinical significance and therapy. Parasites Vectors 2009, 2, 1–6. [Google Scholar] [CrossRef]
- McCraw, B.M.; Slocombe, J.O.D. Strongylus vulgaris in the horse: A review. Can. Vet. J. 1976, 17, 150–157. [Google Scholar]
- Ogbourne, C.P.; Duncan, J.L. Strongylus Vulgaris in the Horse: Its Biology and Veterinary Importance, 2nd ed.; Commonwealth Agricultural Bureaux: Slough, UK, 1985. [Google Scholar]
- Kane, A.J.; Traub-Dargatz, J.; Losinger, W.C.; Garber, L.P. The occurrence and causes of lameness and laminitis in the US horse population. Proc. Annu. Conv. Am. Assoc. Equine Pract. 2000, 46, 277–280. [Google Scholar]
- Ashley, F.H.; Waterman-Pearson, A.E.; Whay, H.R. Behavioural assessment of pain in horses and donkeys: Application to clinical practice and future studies. Equine Vet. J. 2005, 37, 565–575. [Google Scholar] [CrossRef]
- Hausberger, M.; Fureix, C.; Lesimple, C. Detecting horses’ sickness: In search of visible signs. Appl. Anim. Behav. Sci. 2016, 175, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Lesimple, C. Indicators of Horse Welfare: State-of-the-Art. Animals 2020, 10, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, S.R.B.; Asa, C.; Pluhacek, J.; Houpt, K.; Ransom, J.I. Chapter 3: Behaviour of horses, zebras and asses. In Wild Equids Ecology, Management and Conservation; Ransom, J.I., Kaczensky, P., Eds.; John Hopkins University Press: Baltimore, MD, USA, 2016; pp. 23–40. [Google Scholar]
- Waring, G.H. Horse Behaviour, 2nd ed.; Noyes: Norwich, NJ, USA, 2003. [Google Scholar]
- Fureix, C.; Bourjade, M.; Henry, S.; Sankey, C.; Hausberger, M. Exploring aggression regulation in managed groups of horses Equus caballus. Appl. Anim. Behav. 2012, 138, 216–228. [Google Scholar] [CrossRef] [Green Version]
- Popescu, S.; Diugan, E.A. The relationship between behavioral and other welfare indicators of working horses. J. Equine Vet. Sci. 2013, 33, 1–12. [Google Scholar] [CrossRef]
- Whitehead, G.; French, J.; Ikin, P. Welfare and veterinary care of donkeys. Practice 1991, 13, 62–68. [Google Scholar] [CrossRef]
- Duffield, H.F.; Bell, N.; Henson, F.M.D. Factors associated with impactive colic in the donkey. In Proceedings of the 7th International Equine Colic Research Symposium, Equine Veterinary Journal, Newmarket, ON, USA, 2022; p. 122. [Google Scholar]
- Taylor, P.M.; Pascoe, P.J.; Mama, K.R. Diagnosing and treating pain in the horse; where are we today? Vet. Clin. N. Am. Equine Pract. 2002, 18, 1–19. [Google Scholar] [CrossRef]
- Dalla Costa, E.; Minero, M.; Lebelt, D.; Stucke, D.; Canali, E.; Leach, M.C. Development of the Horse Grimace Scale (HGS) as a Pain Assessment Tool in Horses Undergoing Routine Castration. PLoS ONE 2014, 9, e92281. [Google Scholar] [CrossRef] [Green Version]
- Boyd, L. Behaviour problems of equids in zoos. Vet. Clin. N. Am. Equine Pract. 1986, 2, 653–664. [Google Scholar] [CrossRef]
- Divers, T.J.; Mohammed, H.O.; Cummings, J.F.; Valentine, B.A.; De Lahunta, A.; Jackson, C.A.; Summers, B.A. Equine motor neuron disease: Findings in 28 horses and proposal of a pathophysiological mechanism for the disease. Equine Vet. J. 1994, 26, 409–415. [Google Scholar] [CrossRef]
- De la Rúa-Domènech, R.; Mohammed, H.O.; Cummings, J.F.; Divers, T.J.; De Lahunta, A.; Summers, B.A. Association between plasma vitamin E concentration and the risk of equine motor neuron disease. Vet. J. 1997, 154, 203–213. [Google Scholar] [CrossRef]
- Klingel, H. Social organization and reproduction in equids. J. Reprod. Fertil. 1975, 23, 7–11. [Google Scholar]
- Klingel, H. Social organisation of feral horses. J. Reprod. Fertil. 1982, 32, 89–95. [Google Scholar]
- Rubenstein, D.I. The ecology of female social behaviour in horses, zebras and assses. In Animal Societies: Individuals, Interactions and Organisations; Jarma, P., Rossiter, A., Eds.; Kyoto University Press: Kyoto, Japan, 1994; pp. 13–28. [Google Scholar]
- Keiper, R.R. Social Structure. Vet. Clin. N. Am. Equine Pract. 1986, 2, 465–484. [Google Scholar] [CrossRef] [PubMed]
- Roelle, J.E.; Singer, F.J.; Zeigenfuss, L.C.; Ransom, J.I.; Coates-Markle, L.; Schoenecker, K.A. Demography of the Pryor Mountain Wild Horses 1993–2007; Scientific Investigations Report 2010-5125; US Geological Survey: Reston, VA, USA, 2010.
- Pacheco, M.A.; Herrera, E.A. Social structure of feral horses in the Ilanos of Venuzuela. J. Mammal. 1997, 78, 15–22. [Google Scholar] [CrossRef]
- Rubenstein, D.I. Ecology and sociality in horses and zebras. In Ecological Aspects of Social Evolution; Rubenstein, D.I., Wrangham, R.W., Eds.; Princeton University Press: Princeton, NJ, USA, 1986; pp. 282–302. [Google Scholar]
- Boyd, L.; Keiper, R.R. Behavioural ecology of feral horses. In The Domestic Horse: The Origins, Development and Management of Its Behaviour; Mills, D.S., McDonnell, S.M., Eds.; Cambridge University Press: Cambridge, UK, USA, 2005; pp. 55–82. [Google Scholar]
- Miller, R. Band organisation and stability in Red Desert feral horses. In Symposium on the Ecology and Behaviour of Wild and Feral Equids; Denniston, R.H., Ed.; University of Wyoming: Laramie, WY, USA, 1979; pp. 113–128. [Google Scholar]
- Salter, R.E.; Hudson, R.J. Social organisation of feral horse in western Canada. Appl. Anim. Ethol. 1979, 8, 207. [Google Scholar] [CrossRef]
- Kaseda, Y.; Khalil, A.M. Harem size and reproductive success of stallions in Misaki feral horses. Appl. Anim. Behav. Sci. 1996, 47, 163–173. [Google Scholar] [CrossRef]
- Scorolli, A.L.; Lopez-Cazorla, A.C. Demography of feral horses (Equus caballus): A long-term study in Tornquist Park, Argentina. Wildl. Res. 2010, 37, 207–214. [Google Scholar] [CrossRef]
- Rubenstein, D.I. Behavioural ecology of island feral horses. Equine Vet. J. 1981, 13, 27–34. [Google Scholar] [CrossRef]
- Feh, C. Alliances and reproductive success in Camargue stallions. Anim. Behav. 1999, 57, 705–713. [Google Scholar] [CrossRef] [Green Version]
- Goodloe, R.B.; Warren, R.J.; Osborn, D.A.; Hall, C. Population characteristics of feral horses on Cumberland Island, Georgia, and their management implications. J. Wildl. Manag. 2000, 64, 114–121. [Google Scholar] [CrossRef] [Green Version]
- Cameron, E.Z.; Linklater, W.L.; Minot, E.O.; Stafford, K.J. Population Dynamics 1994–1998, and Management of Kaimanawa wild Horses; Science for Conservation Publication 171; Department of Conservation: Otago, New Zealand, 2001.
- Franke-Stevens, E. Instability of harems of feral horses in relation to season and presence of subordinate stallions. Behaviour 1990, 112, 149–161. [Google Scholar] [CrossRef]
- Linklater, W.L.; Cameron, E.Z.; Stafford, K.J. Stallion harassment and the mating system of horses. Anim. Behav. 1999, 58, 295–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginsberg, J.R. The ecology of female behaviour and the male mating success in Grevy’s zebra. Symp. Zool. Soc. Lond. 1989, 61, 89–110. [Google Scholar]
- Berger, J. Organisational systems and dominance in feral horses in the Grand Canyon. Behav. Ecol. Sociobiol. 1977, 2, 131–146. [Google Scholar] [CrossRef]
- Franke-Stevens, E. Contests between bands of feral horses for access to fresh water: The resident wins. Anim. Behav. 1988, 36, 1851–1853. [Google Scholar] [CrossRef]
- Welsh, D.A. The life of Sable Island’s wild horses. Nat. Can. 1973, 2, 7–14. [Google Scholar]
- Khalil, A.M.; Murakami, N. Factors affecting the harem formation process by young Misaki feral stallions. J. Vet. Med. Sci. 1999, 61, 667–671. [Google Scholar] [CrossRef] [Green Version]
- Rutberg, A.T.; Keiper, R.R. Proximate causes of natal dispersal in feral ponies: Some sex differences. Anim. Behav. 1993, 46, 969–975. [Google Scholar] [CrossRef]
- Linklater, W.L.; Cameron, E.Z. Tests for cooperative behaviour between stallions. Anim. Behav. 2000, 60, 731–743. [Google Scholar] [CrossRef] [Green Version]
- Kaseda, Y.; Khalil, A.M.; Ogawa, H. Harem stability and reproductive success of Misaki feral mares. Equine Vet. J. 1995, 27, 163–173. [Google Scholar] [CrossRef]
- Boyd, L. The Natality, Foal Survivorship and Mare-Foal Behaviour of Feral Horses in Wyoming’s Red Desert. Ph.D. Thesis, University of Wyoming, Laramie, WY, USA, 1980. [Google Scholar]
- Fowler, C.W. A review of density dependence in populations of large mammals. In Current Mammology; Genoways, H.H., Ed.; Plenum: New York, NY, USA, 1987; pp. 401–441. [Google Scholar]
- Gaillard, J.M.; Fest-Bianchet, M.; Yoccuz, N.G. Population dynamics of large herbivores: Variable recruitment with constant adult survival. Trends Ecol. Evol. 1998, 13, 58–63. [Google Scholar] [CrossRef]
- Gaidet, N.; Gaillard, J.M. Density dependent body condition and recruitment in a tropical ungulate. Can. J. Zool. 2008, 86, 24–32. [Google Scholar] [CrossRef]
- Eberhardt, L.L. Population projections from simple models. J. Appl. Ecol. 1987, 24, 103–118. [Google Scholar] [CrossRef]
- Tatin, L.; King, S.R.B.; Munkhtuya, B.; Hewison, A.J.M.; Feh, C. Demography of a socially natural herd of Przewalski’s horses: An example of a small, closed population. J. Zool. 2009, 277, 134–140. [Google Scholar] [CrossRef]
- Ransom, J.I.; Lagos, L.; Hrabar, H.; Nowzari, H.; Usukhjargal, D.; Spasskaya, N. Chapter 6: Wild and feral equid population dynamics. In Wild Equids Ecology, Management and Conservation; Ransom, J.I., Kaczensky, P., Eds.; John Hopkins University Press: Baltimore, MD, USA, 2016; pp. 68–83. [Google Scholar]
- Roberts. S.J. Abortion and other gestational diseases in mares. In Current Therapy in Theriogenology; Morrow, D.A., Ed.; W.B. Saunders: Philadelphia, PA, USA, 1986; pp. 705–710. [Google Scholar]
- Siniff, D.B.; Tester, J.R.; McMahaon, G.L. Foaling rate and survival of feral horses in western Nevada. J. Range Manag. 1986, 39, 296–297. [Google Scholar] [CrossRef]
- Ransom, J.I.; Hobbs, N.T.; Bruemmer, J. Contraception can lead to trophic asynchrony between birth pulse and resources. PLoS ONE 2013, 8, e54972. [Google Scholar]
- Ransom, J.I. Population Ecology of Feral Horses in an Era of Fertility Control Management. Ph.D. Thesis, Colarado State University, Fort Collins, CO, USA, 2012. [Google Scholar]
- Eberhardt, L.; Majorowicz, A.; Wilcox, J. Apparent rates of increase for two feral horse herds. J. Wildl. Manag. 1982, 46, 367–374. [Google Scholar] [CrossRef]
- Garrott, R.A.; Siniff, D.B.; Eberhardt, L.L. Growth Rates of Feral Horse Populations. J. Wildl. Manag. 1991, 55, 641–648. [Google Scholar] [CrossRef]
- Kirkpatrick, J.F.; Turner, A. Immunocontraception and increased longevity in equids. Zoo Biol. 2007, 26, 237–244. [Google Scholar] [CrossRef]
- Dorj, U.; Namkhai, B. Reproduction and mortality of re-introduced Przewalski horses in Hustai National Park, Mongolia. J. Life Sci. 2013, 7, 624–631. [Google Scholar]
- Kie, J.G. Optimal foraging and risk predation: Effects on behaviour and social structure in ungulates. J. Mammal 1999, 80, 1114–1129. [Google Scholar] [CrossRef] [Green Version]
- Ransom, J.I.; Kaczensky, P. Wild Equids Ecology, Management and Conservation; John Hopkins University Press: Baltimore, MD, USA, 2016. [Google Scholar]
- Davis, N.E.; Forsyth, D.M.; Triggs, B.; Pascoe, C.; Benshemesh, J.; Robley, A.; Lawrence, J.; Ritchie, E.G.; Nimmo, D.G.; Lumsden, L.F. Interspecific and geographic variation in the diets of sympatric carnivores: Dingoes/wild dogs and red foxes in south-eastern Australia. PLoS ONE 2015, 10, e0120975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Research Council. Using Science to Improve the BLM Wild Horse and Burro Program: A Way Forward; National Academies Press: Washington, DC, USA, 2013. [Google Scholar]
- Hobbs, R.J.; Hinds, L.A. Could current fertility control methods be effective for landscape-scale management of populations of wild horses (Equus caballus) in Australia? Wildl. Res. 2018, 45, 195–207. [Google Scholar] [CrossRef]
- Feist, J.D.; McCullough, D.R. Reproduction in feral horses. J. Reprod. Fertil. 1975, 23 (Suppl. 23), 13–18. [Google Scholar]
- Boyd, L.; Houpt, K.A. Activity patterns. In Przewalski’s Horse: The History and Biology of an Endangered Species; Boyd, L., Houpt, K.A., Eds.; State University of New York Press: Albany, NY, USA, 1994; pp. 195–254. [Google Scholar]
- Ransom, J.I.; Cade, B.S. Quantifying equid behavior: A research ethogram for free-roaming feral horses. In U.S. Geological Survey Techniques and Methods Report 2-A9; USGS Fort Collins Science Center: Fort Collins, CO, USA, 2009. [Google Scholar]
- Asa, C.S. Sexual behaviour of mares. Vet. Clin. N. Am. Equine Pract. 1986, 2, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Monfort, S.L.; Arthur, N.P.; Wildt, D.E. Reproduction on the Przewalski’s horse. In Przewalski’s Horse: The History and Biology of an Endangered Species; Boyd, L., Houpt, K., Eds.; State University of New York Press: Albany, NY, USA, 1994; pp. 173–193. [Google Scholar]
- Mendl, M.; Paul, E.S. Observation of nursing and sucking behaviour as an indicator of milk transfer and parental investment. Anim. Behav. 1989, 37, 513–515. [Google Scholar] [CrossRef]
- Duncan, P.; Harvey, P.H.; Wells, S.M. On lactation and associated behaviour in a natural herd of horses. Anim. Behav. 1984, 32, 255–263. [Google Scholar] [CrossRef]
- Cameron, E.Z.; Linklater, W.L. Individual mares bias investment in sons and daughters in relation to their condition. Anim. Behav. 2000, 60, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Cameron, E.; Linklater, W.; Stafford, K.; Minot, E.O. Social grouping and maternal behaviour in feral horses (Equus caballus): The influence of males on maternal protectiveness. Behav. Ecol. Sociobiol. 2003, 53, 92–101. [Google Scholar] [CrossRef]
- Berger, A.; Scheibe, K.M.; Eichorn, K.; Scheibe, A.; Streich, J. Diurnal and ultradian rhythms of behaviour in a mare group of Przewalski horse measured through one year under semi-reserve conditions. Anim. Behav. 1999, 64, 1–17. [Google Scholar] [CrossRef]
- Souris, A.; Kaczensky, P.; Julliard, R.; Walzer. Time budget, behavioural synchrony and body score development of a newly released Przewalski’s horse group in the Great Gobi B Strictly Protected Area in SW Mongolia. Appl. Anim. Behav. Sci. 2007, 107, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Duncan, P. Time-budgets of Camargue horses III: Environmental influences. Behaviour 1985, 92, 188–208. [Google Scholar] [CrossRef]
- Mayes, E.; Duncan, P. Temporal patterns of feeding behaviour in free-ranging horses. Behaviour 1986, 96, 105–129. [Google Scholar] [CrossRef]
- Arnold, G.W. Comparison of time budgets and circadian patterns of maintenance activities in sheep, cattle and horses grouped together. Appl. Anim. Behav. Sci. 1984, 13, 19–30. [Google Scholar] [CrossRef]
- King, S.R.B.; Gurnell, J. Scent marking behaviour by stallions: An assessment of function in a reintroduced population of Przewalski horses (Equus ferus przewalskii. J. Zool. 2006, 272, 30–36. [Google Scholar] [CrossRef]
- Kaseda, Y. Seasonal changes in time spent grazing and resting of Misaki horses. Jpn. J. Zootech. Sci. 1983, 54, 464–469. [Google Scholar]
- King, S.R.B. Behavioural Ecology of Przewalski Horses Reintroduced to Hustai National Park, Mongolia. Ph.D. Thesis, Queen Mary, University of London, London, UK, 2002. [Google Scholar]
- Ransom, J.I.; Cade, B.S.; Hobbs, N.T. Influences of immunocontraception on time budgets, social behaviour, and body condition in feral horses. Appl. Anim. Behav. Sci. 2010, 124, 51–60. [Google Scholar] [CrossRef]
- Boyd, L. Time budgets of adult Przewalski horses: Effects of sex, reproductive status and enclosure. Appl. Anim. Behav. Sci. 1988, 60, 291–299. [Google Scholar] [CrossRef]
- Madosky, J.M. Factors that affect harem stability in a feral horse population on Shackleford Banks Island, NC. Ph.D. Thesis, University of New Orleans, New Orleans, LA, USA, 2011. [Google Scholar]
- Tyler, S. The behaviour and social organisation of the New Forest ponies. Anim. Behav. Monogr. 1972, 5, 87–196. [Google Scholar] [CrossRef]
- Boy, V.; Duncan, P. Time budgets of Camargue horses I: Developmental changes in the time-budgets of foals. Behaviour 1979, 71, 187–202. [Google Scholar] [CrossRef]
- McDonnell, S.M. The Equid Ethogram: A Practical Field Guide to Horse Behaviour; CAB International: Cambridge, MA, USA, 2003. [Google Scholar]
- Feh, C.; de Mazieres, J. Grooming at a preferred site reduces heart rate in horses. Anim. Behav. 1993, 46, 1191–1194. [Google Scholar] [CrossRef] [Green Version]
- Houpt, K.A.; Boyd, L. Social Behaviour. In Przewalski’s Horse: The History and Biology of an Endangered Species; Boyd, L., Houpt, K.A., Eds.; State University of New York Press: Albany, NY, USA, 1994; pp. 229–254. [Google Scholar]
- Wolter, R.; Stefanski, V.; Krueger, K. Parameters for the Analysis of Social Bonds in Horses. Animals 2018, 8, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, E.Z.; Setsaas, T.H.; Linklater, W.L. Social bonds between unrelated females increases reproductive success in feral horses. Proc. Natl. Acad. Sci. USA 2009, 106, 13850–13853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, R. Mutual grooming and preferred associate relationships in a band of free-ranging horses. Appl. Anim. Behav. Sci. 1998, 59, 265–276. [Google Scholar] [CrossRef]
- Sigurjónsdóttir, H.; Haraldsson, H. Significance of group composition for the welfare of pastured horses. Animals 2019, 9, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Dierendonck, M.C.; Sigurjonsdottir, H.; Colenbrander, B.; Thorhallsdottir, A.G. Differences in social behaviour between late pregnant, post-partum and barren mares in a herd of Icelandic ponies. Appl. Anim. Behav. Sci. 2004, 89, 283–297. [Google Scholar] [CrossRef] [Green Version]
- Crowell-Davis, S.L.; Houpt, K.A.; Carini, C.M. Mutual grooming and nearest neighbour relationships among foals of Equus caballus. Appl. Anim. Behav. Sci. 1986, 15, 113–123. [Google Scholar] [CrossRef]
- van Dierendonck, M.C.; de Vries, H.; Schilder, M.B.H. An analysis of dominance, its behavioural parameters and possible determinants in a herd of Icelandic horses in captivity. J. Zool. 1995, 45, 362–385. [Google Scholar]
- McDonnell, S.M.; Poulin, A. Equid play ethogram. Appl. Anim. Behav. Sci. 2002, 78, 263–290. [Google Scholar] [CrossRef]
- Feh, C. Social behaviour and relationships of Przewalski horses in Dutch semi-reserves. Appl. Anim. Behav. Sci. 1988, 21, 171–187. [Google Scholar] [CrossRef]
- Moehlman, P.D. Behavioural patterns and communication in feral asses (Equus africanus). Appl. Anim. Behav. Sci. 1998, 60, 125–169. [Google Scholar] [CrossRef]
- McDonnell, S.M.; Haviland, J.C.S. Agonistic ethogram of the equid bachelor band. Appl. Anim. Behav. Sci. 1995, 43, 147–188. [Google Scholar] [CrossRef]
- Rutberg, A.T.; Greenberg, S. Dominance, aggression frequencies and modes of aggressive competition in feral pony mares. Anim. Behav. 1990, 40, 945–952. [Google Scholar] [CrossRef]
- Rho, J.R.; Srygley, R.B.; Choe, J.C. Behavioural ecology of the Jeju pony (Equus caballus): Effects of aternal age, maternal dominance hierarchy and foal age on mare aggression. Ecol. Res. 2004, 19, 55–63. [Google Scholar] [CrossRef]
- Houpt, K.A.; Wolski, I. Stability of equine hierarchies and the prevention of dominance related aggression. Equine Vet. J. 1980, 12, 15–18. [Google Scholar] [CrossRef]
- Keiper, R.R.; Receveur, H. Social interactions of free-ranging Przewalski horses in semi-reserves in the Netherlands. Appl. Anim. Behav. Sci. 1992, 33, 303–318. [Google Scholar] [CrossRef]
- Feh, C. Long term paternity data in relation to different aspects of rank for Camargue stallions, Equus caballus. Anim. Behav. 1990, 40, 995–996. [Google Scholar] [CrossRef]
- Powell, D.M. Female-female competition or male mate choice? Patterns of courtship and breeding behaviour among feral horses (Equus caballus) on Assateague Island. J. Ethol. 2008, 26, 137–144. [Google Scholar] [CrossRef]
- Krueger, K.B.; Flauger, B.; Farmer, K.; Hemelrijk, C.K. Movement initiation in groups of feral horses. Behav. Process. 2014, 103, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Tilson, R.; Sweeny, K.; Binclik, G.; Reindle, N. Buddies and bullies: Social structure of a bachelor group of Przewalski horses. Appl. Anim. Behav. Sci. 1988, 21, 169–185. [Google Scholar] [CrossRef]
- Rubenstein, D.I.; Hack, M.A. Horse signals: The sounds and scents of fury. Evol. Ecol. 1992, 6, 254–260. [Google Scholar] [CrossRef]
- Proops, L.; McComb, K.; Reby, D. Cross-modal individual recognition in domestic horses (Equus caballus). Proc. Natl. Acad. Sci. USA 2009, 106, 947–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krueger, K.B.; Flauger, B. Olfactory recognition of individual competitors by means of faeces in horse (Equus caballus). Anim. Cogn. 2011, 14, 245–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rørvang, M.V.; Nielsen, B.L.; McLean, A.N. Sensory Abilities of Horses and Their Importance for Equitation Science. Front. Vet. Sci. 2020, 7, 633. [Google Scholar] [CrossRef] [PubMed]
- Timney, B.; Macuda, T. Vision and hearing in horses. J. Am. Vet. Med. Assoc. 2001, 218, 1567–1574. [Google Scholar] [CrossRef]
- Policht, R.; Karadzos, A.; Frynta, D. Comparative analysis of long-range calls in equid stallions: Are acoustic parameters related to social organisation? Afr. Zool. 2011, 46, 18–26. [Google Scholar] [CrossRef]
- Basile, M.; Boivin, S.; Boutin, A.; Blois-Heulin, C.; Hausberger, M.; Lemasson, A. Socially dependent auditory laterality in domestic horses (Equus caballus). Anim. Cogn. 2009, 12, 611–619. [Google Scholar] [CrossRef]
- Stomp, M.; Leroux, M.; Cellier, M.; Henry, S.; Lemasson, A.; Hausberger, M. An unexpected acoustic indicator of positive emotions in horses. PLoS ONE 2018, 13, e0197898. [Google Scholar] [CrossRef]
- Saslow, C.A. Understanding the perceptual world of horses. Appl. Anim. Behav. Sci. 2002, 78, 209–224. [Google Scholar] [CrossRef]
- Stahlbaum, C.C.; Houpt, K.A. The role of the Flehmen response in the behavioral repertoire of the stallion. Physiol. Behav. 1989, 45, 1207–1214. [Google Scholar] [CrossRef]
- Hothersall, B.; Harris, P.; Sortoft, L.; Nicol, C.J. Discrimination between conspecific odour samples in the horse (Equus caballus). Appl. Anim. Behav. Sci. 2010, 126, 37–44. [Google Scholar] [CrossRef]
- Harvey, A.M. Wild Horse Welfare: Assessment and Associations with Population and Behavioural Ecology. Ph.D. Thesis, University of Technology Sydney, Ultimo, NSW, Australia, 2022. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
|
| Domain | Species-Specific Information Required |
|---|---|
| 1: Nutrition | Water requirements: volume, frequency, preferred water sources, factors influencing water requirements, adaptations to and impacts of water restriction |
| Nutritional requirements and preferences | |
| Common nutritional deficiencies and excesses and their causes, plant toxicities | |
| Assessing body condition, body condition scoring systems, optimal body condition score, factors affecting body condition | |
| 2: Physical environment | Habitat preferences, and factors affecting habitat selection and use |
| Preferred underfoot substrate and terrain | |
| Thermoneutral zone, impacts of extreme climate events, signs of thermal discomfort | |
| 3: Health | Common non-infectious diseases and their clinical signs, risk factors, aetiologies, diagnosis and prognosis |
| Common infectious diseases and their clinical signs, epidemiology, mode of infection, characteristics of infectious agent (e.g., life cycle, survival in environment, involvement of other species) | |
| Common injuries and their clinical signs, risk factors, aetiologies, diagnosis and prognosis | |
| Sickness and pain behaviours | |
| 4: Behavioural interactions | Social organisation and factors affecting it |
| Population dynamics | |
| Reproductive physiology and behaviours; oestrous, courtship, mating, gestation, parturition, lactation, maternal and newborn behaviour | |
| Normal range of behaviours and time budgets | |
| Social behaviour (including ‘rewarding behaviours’, e.g., play) and communication |
| Body Condition Score | Description | Example Photo |
|---|---|---|
| 1 (Poor) | Extremely emaciated. Spinous processes, ribs, tailhead, tuber coxae and ischii projecting prominently. Bone structure of withers, shoulders and neck easily visible. No fatty tissue. |  |
| 2 (Very thin) | Emaciated. Slight tissue covering over base of spinous processes. Spinous processes, ribs, tailhead, tuber coxae and ischii prominent. Withers, shoulders and neck structures visible. |  |
| 3 (Thin) | Slight fat coverage of spinous processes and ribs but still visible. Tailhead prominent, but individual vertebrae cannot be visually identified. Tuber coxae rounded but easily discernible. Withers, shoulders and neck structures visible but with some tissue coverage. |  |
| 4 (Moderately thin) | Ridge of spine and outline of ribs just visible, tailhead not prominent. Tuber coxae not discernible. Withers, shoulders and neck not obviously thin. |  |
| 5 (Moderate) | Spine and ribs cannot be seen but ribs can be felt. Fat around tailhead. Withers rounded over spinous processes. Shoulders and neck rounded and blend smoothly into body. |  |
| 6 (Moderately fleshy) | Slight crease down spine. Fat over ribs and tailhead. Fat deposits along sides of withers, behind shoulders and along the sides of the neck. |  |
| 7 (Fleshy) | Crease down spine. Fat filling between ribs and around tailhead. Fat deposited along withers, behind shoulders and along the neck. |  |
| 8 (Fat) | Crease down spine. Difficult to feel ribs. Fat around tailhead. Area along withers filled with fat. Fat deposits along withers and behind shoulders. Noticeable thickening of neck. Fat deposited along inner thighs. |  |
| 9 (Extremely fat) | Obvious crease down spine. Patchy fat appearing over ribs. Bulging fat around tailhead, along withers, behind shoulders and along neck. Fat fills in flank and is distributed along inner thighs. |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harvey, A.M.; Ramp, D.; Mellor, D.J. Review of the Foundational Knowledge Required for Assessing Horse Welfare. Animals 2022, 12, 3385. https://doi.org/10.3390/ani12233385
Harvey AM, Ramp D, Mellor DJ. Review of the Foundational Knowledge Required for Assessing Horse Welfare. Animals. 2022; 12(23):3385. https://doi.org/10.3390/ani12233385
Chicago/Turabian StyleHarvey, Andrea M., Daniel Ramp, and David J. Mellor. 2022. "Review of the Foundational Knowledge Required for Assessing Horse Welfare" Animals 12, no. 23: 3385. https://doi.org/10.3390/ani12233385