
Molecular Dietary Analysis of Three Sympatric Mustelidae in Northeast China

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
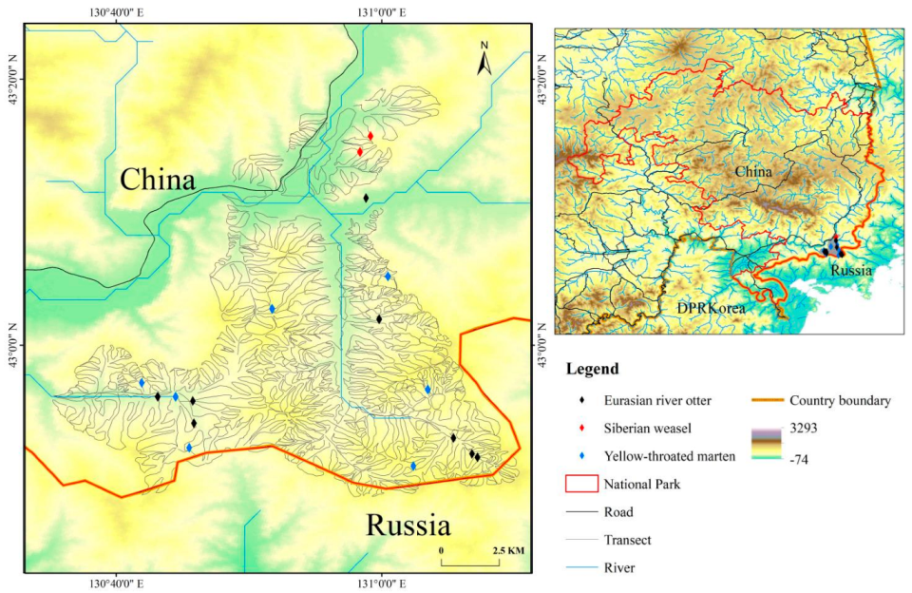
2.1. Study Area and Sample Collection
2.2. Molecular Species Identification
2.3. Diet Analysis
2.4. Data Analysis
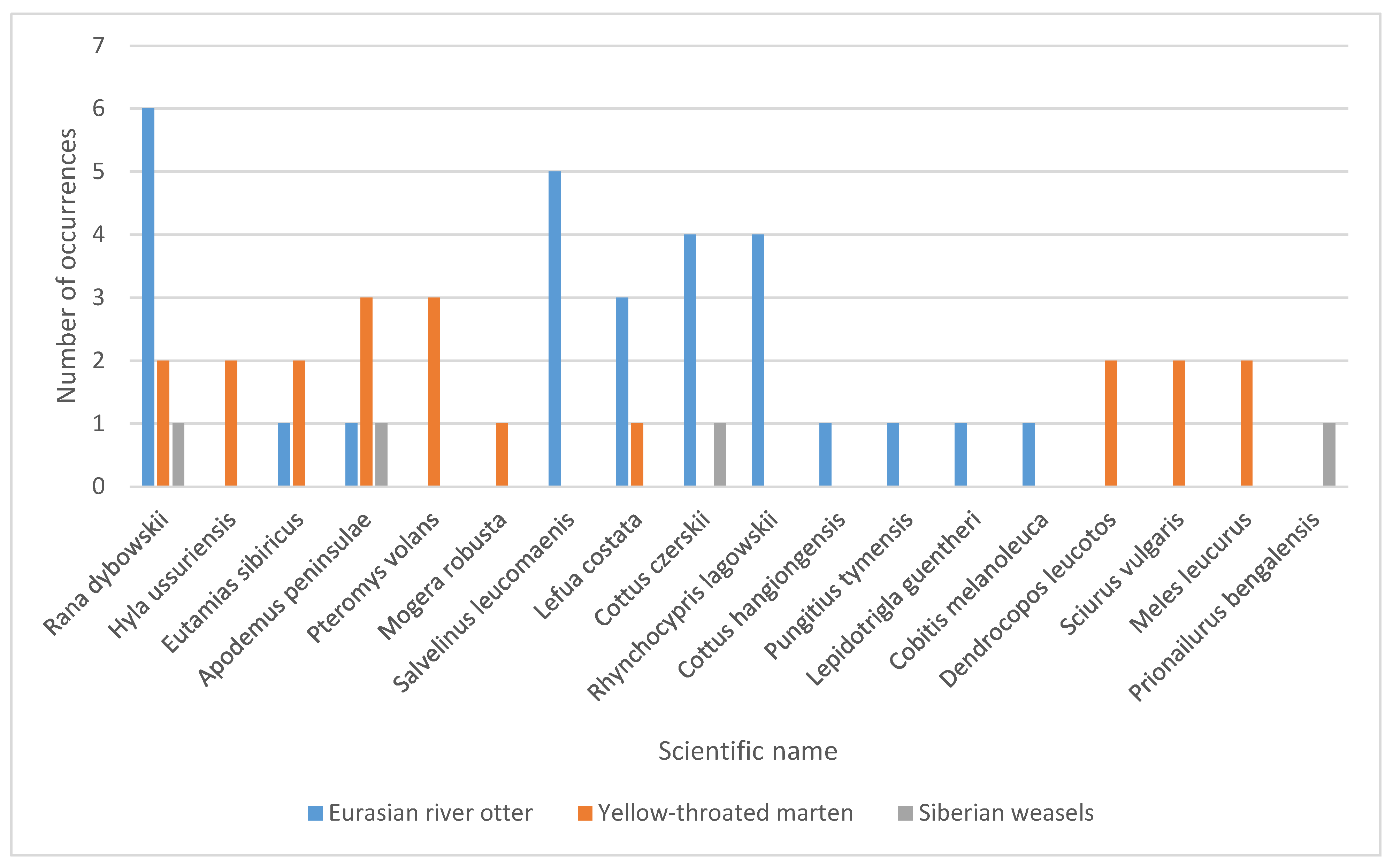
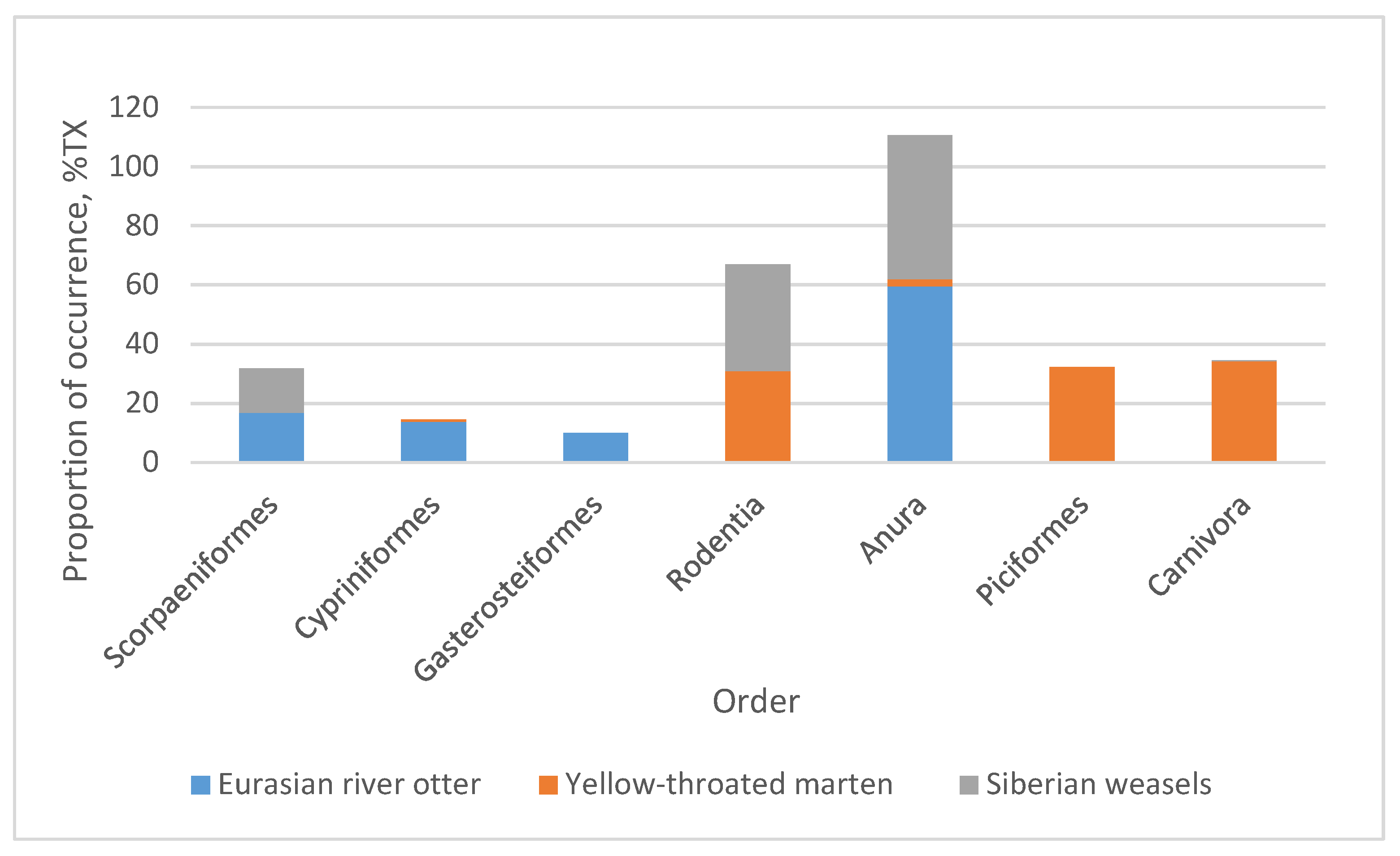
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carreon-Martinez, L.; Johnson, T.B.; Ludsin, S.A.; Heath, D.D. Utilization of stomach content DNA to determine diet diversity in piscivorous fishes. J. Fish Biol. 2011, 78, 1170–1182. [Google Scholar] [CrossRef] [PubMed]
- Symondson, W.O.C. Molecular identification of prey in predator diets. Mol. Ecol. 2002, 11, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Yang, C.; Ma, J.; Zhang, X.; Ran, J. Vertebrate prey composition analysis of the Pallas’s cat (Otocolobus manul) in the Gongga Mountain Nature Reserve, based on fecal DNA. Mammalia 2020, 84, 449–457. [Google Scholar] [CrossRef]
- Estes, J.A.; Terborgh, J.; Brashares, J.S.; Power, M.E.; Berger, J.; Bond, W.J.; Carpenter, S.R.; Essington, T.E.; Holt, R.D.; Jackson, J.B.C.; et al. Trophic Downgrading of Planet Earth. Science 2011, 333, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Shao, X.; Lu, Q.; Xiong, M.; Bu, H.; Shi, X.; Wang, D.; Zhao, J.; Li, S.; Yao, M. Prey partitioning and livestock consumption in the world’s richest large carnivore assemblage. Curr. Biol. 2021, 31, 4887–4897. [Google Scholar] [CrossRef]
- Xiong, M.; Wang, D.; Bu, H.; Shao, X.; Zhang, D.; Li, S.; Wang, R.; Yao, M. Molecular dietary analysis of two sympatric felids in the Mountains of Southwest China biodiversity hotspot and conservation implications. Sci. Rep. 2017, 7, 41909. [Google Scholar] [CrossRef] [Green Version]
- Koepfli, K.-P.; Deere, K.A.; Slater, G.J.; Begg, C.; Begg, K.; Grassman, L.; Lucherini, M.; Veron, G.; Wayne, R.K. Multigene phylogeny of the Mustelidae: Resolving relationships, tempo and biogeographic history of a mammalian adaptive radiation. BMC Biol. 2008, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Donadio, E.; Di Martino, S.; Aubone, M.; Novaro, A.J. Activity patterns, home-range, and habitat selection of the common hog-nosed skunk, Conepatus chinga (Mammmalia, Mustelidae), in northwestern Patagonia. Mammalia 2001, 65, 49–54. [Google Scholar] [CrossRef]
- Zhou, Y.-B.; Slade, E.; Newman, C.; Wang, X.-M.; Zhang, S.-Y. Frugivory and seed dispersal by the yellow-throated marten, Martes flavigula, in a subtropical forest of China. J. Trop. Ecol. 2008, 24, 219–223. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Wang, B.; Ran, J.; Li, B.; Huang, F.; Li, X.; Gu, X. Seasonal variation of daily activity patterns and diet of yellow-throated marten (Martes flavigula). ACTA Theriol. Sin. 2019, 39, 52–61. [Google Scholar]
- Smiroldo, G.; Balestrieri, A.; Remonti, L.; Prigioni, C. Seasonal and habitat-related variation of otter Lutra lutra diet in a Mediterranean river catchment (Italy). Folia Zool. Praha 2009, 58, 87–97. [Google Scholar]
- Remonti, L.; Balestrieri, A.; Prigioni, C. Altitudinal gradient of Eurasian otter (Lutra lutra) food niche in Mediterranean habitats. Can. J. Zool. 2009, 87, 285–291. [Google Scholar] [CrossRef]
- Sasaki, H.; Ono, Y. Habitat Use and Selection of the Siberian Weasel Mustela sibirica coreana during the Non-mating Season. J. Mammal. Soc. Jpn. 1994, 19, 21–32. [Google Scholar]
- Tsai, C.W. Food Resource Partitioning between Sympatric Yellow-throated Marten (Martes flavigula chrysosphila) and Siberian Weasel (Mustela sibirica taivana). Master’s Thesis, National Taiwan University, Taipei, Taiwan, 2007; pp. 1–82. [Google Scholar]
- Yang, H.; Han, S.; Xie, B.; Mou, P.; Kou, X.; Wang, T.; Ge, J.; Feng, L. Do prey availability, human disturbance and habitat structure drive the daily activity patterns of Amur tigers (Panthera tigris altaica)? J. Zool. 2019, 307, 131–140. [Google Scholar] [CrossRef]
- Pompanon, F.; Deagle, B.E.; Symondson, W.O.C.; Brown, D.S.; Jarman, S.N.; Taberlet, P. Who is eating what: Diet assessment using next generation sequencing. Mol. Ecol. 2012, 21, 1931–1950. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Feng, J.; Tseveen, A.; Yang, L.; Huang, C.; Li, D.; Zhu, X.; Feng, L.; Wang, T.; Jianping, G.E.; et al. Forest cattle grazing affects understory food resource of ungulates in the eastern part of the Northeast Tiger and Leopard National Park. Acta Theriol. Sin. 2019, 39, 386–396. [Google Scholar]
- Yang, S.H.; Huang, X.M.; Xia, R.; Xu, Y.C.; Dahmer, T.D. Use of femur bone density to segregate wild from farmed Dybowski’s frog (Rana dybowskii). Forensic Sci. Int. 2011, 207, 61–65. [Google Scholar] [CrossRef]
- Stankowich, T. Ungulate flight responses to human disturbance: A review and meta-analysis. Biol. Conserv. 2008, 141, 2159–2173. [Google Scholar] [CrossRef]
- Bascompte, J.; Melian, C.J.; Sala, E. Interaction strength combinations and the overfishing of a marine food web. Proc. Natl. Acad. Sci. USA 2005, 102, 5443–5447. [Google Scholar] [CrossRef] [Green Version]
- Long, S.X.; Chen, C. Ecological characteristics of planktonic diatoms and zooplankton community after the building of dam on the Wujiang River. J. Guizhou Norm. Univ. 2009, 3, 21–26. [Google Scholar]
- Dahshan, H.; Megahed, A.M.; Abd-Elall, A.M.M.; Abd-El-Kader, M.A.-G.; Nabawy, E.; Elbana, M.H. Monitoring of pesticides water pollution-The Egyptian River Nile. J. Environ. Health Sci. Eng. 2016, 14, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piao, Z.J.; Sui, Y.C.; Wang, Q.; Li, Z.; Niu, L.J. Population Fluctuation and Resource Protection of Otter (Lutra lutra) in Changbai Mountain Nature Reserve. J. Hydro Ecol. 2011, 32, 51–55. [Google Scholar]
- Wang, T.; Feng, L.; Yang, H.; Han, B.; Zhao, Y.; Juan, L.; Lü, X.; Zou, L.; Li, T.; Xiao, W.; et al. A science-based approach to guide Amur leopard recovery in China. Biol. Conserv. 2017, 210 Pt B, 47–55. [Google Scholar] [CrossRef]
- Vestheim, H.; Jarman, S.N. Blocking primers to enhance PCR amplification of rare sequences in mixed samples—A case study on prey DNA in Antarctic krill stomachs. Front. Zool. 2008, 5, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Ostell, J.; Pruitt, K.D.; Sayers, E.W. GenBank. Nucleic Acids Res. 2017, 46, D41–D47. [Google Scholar] [CrossRef]
- Riaz, T.; Shehzad, W.; Viari, A.; Pompanon, F.; Taberlet, P.; Coissac, E. ecoPrimers: Inference of new DNA barcode markers from whole genome sequence analysis. Nucleic Acids Res. 2011, 39, e145. [Google Scholar] [CrossRef]
- Shehzad, W.; Riaz, T.; Nawaz, M.A.; Miquel, C.; Poillot, C.; Shah, S.A.; Pompanon, F.; Coissac, E.; Taberlet, P. Carnivore diet analysis based on next-generation sequencing: Application to the leopard cat (Prionailurus bengalensis) in Pakistan. Mol. Ecol. 2012, 21, 1951–1965. [Google Scholar] [CrossRef]
- Xiong, M.; Shao, X.; Long, Y.; Bu, H.; Zhang, D.; Wang, D.; Li, S.; Wang, R.; Yao, M. Molecular analysis of vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) in southwest China. J. Mammal. 2016, 97, 1054–1064. [Google Scholar] [CrossRef]
- Brownstein, M.J.; Carpten, J.D.; Smith, J.R. Modulation of non-templated nucleotide addition by Taq DNA polymerase: Primer modifications that facilitate genotyping. Biotechniques 1996, 20, 1004–1006. [Google Scholar] [CrossRef] [Green Version]
- Boyer, F.; Mercier, C.; Bonin, A.; Le Bras, Y.; Taberlet, P.; Coissac, E. OBITools: A Unix-inspired software package for DNA metabarcoding. Mol. Ecol. Resour. 2016, 16, 176–182. [Google Scholar] [CrossRef] [PubMed]
- De Barba, M.; Miquel, C.; Boyer, F.; Mercier, C.; Rioux, D.; Coissac, E.; Taberlet, P. DNA metabarcoding multiplexing and validation of data accuracy for diet assessment: Application to omnivorous diet. Mol. Ecol. Resour. 2014, 14, 306–323. [Google Scholar] [CrossRef] [PubMed]
- Schnell, I.B.; Bohmann, K.; Gilbert, M.T.P. Tag jumps illuminated—Reducing sequence-to-sample misidentifications in metabarcoding studies. Mol. Ecol. Resour. 2015, 15, 1289–1303. [Google Scholar] [CrossRef]
- Jang-Liaw, N.-H. A barcoding-based scat-analysis assessment of Eurasian otter Lutra lutra diet on Kinmen Island. Ecol. Evol. 2021, 11, 8795–8813. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Abraín, A.; Marí-Mena, N.; Vizcaíno, A.; Vierna, J.; Veloy, C.; Amboage, M.; Guitián-Caamaño, A.; Key, C.; Vila, M. Determinants of Eurasian otter (Lutra lutra) diet in a seasonally changing reservoir. Hydrobiologia 2020, 847, 1803–1816. [Google Scholar] [CrossRef]
- Xiao, W.H.; Feng, L.M.; Zhao, X.D.; Yang, H.T.; Dou, H.L.; Cheng, Y.C.; Mou, P.; Wang, T.M.; Ge, J.P. Distribution and abundance of Amur tiger, Amur leopard and their ungulate preys in Hunchun National Nature Reserve, Jilin. Biodivers. Sci. 2014, 22, 717–724. [Google Scholar]
- Wang, T.; Feng, L.; Yang, H.; Bao, L.; Wang, H.; Ge, J. An introduction to Long-term Tiger-Leopard Observation Network based on camera traps in Northeast China. Biodivers. Sci. 2020, 28, 1059–1066. [Google Scholar] [CrossRef]
- Li, Z.; Wang, T.; Smith, J.L.D.; Feng, R.; Feng, L.; Mou, P.; Ge, J. Coexistence of two sympatric flagship carnivores in the human-dominated forest landscapes of Northeast Asia. Landsc. Ecol. 2019, 34, 291–305. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Name | Target Sequence | Product Size (bp) | Ta (°C) | Primer Sequences (5′-3′) | Reference |
|---|---|---|---|---|---|
| 16S | Carnivore mitochondrial 16S rRNA gene | ~350 bp | 54 | F-GAGAAGACCCTATGGAGC R-ATAGAAACCGACCTGGAT | Xiong et al., 2016 [6] |
| 12SV5 | Vertebrate mitochondrial 12S rRNA gene | ~100 bp | 55 | F-TAGAACAGGCTCCTCTAG R-TTAGATACCCCACTATGC | Riaz et al., 2011 [28] |
| OSB1 | otter mitochondrial 12S rRNA gene | ~100 bp | 56 | CTATGCTCAGCCCTAAACATAGATAGCTTACATAACAAAACTATCTGCC-C3 | This study |
| MartesB | yellow-throated marten mitochondrial 12S rRNA gene | ~100 bp | 55 | CTATGCCCAGCCCTAAACACAAACAATTTACGTAACAAAATTGTCTG-C3 | This study |
| MustelaB | Siberian weasel mitochondrial 12S rRNA gene | ~100 bp | 54 | CTATGCTCAGCCCTAAACATAAATAATTATCACAACAAAATTATCTG-C3 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, D.; Liu, Z.; Tan, M.; Wang, Y.; Dai, W.; Ge, J.; Feng, L. Molecular Dietary Analysis of Three Sympatric Mustelidae in Northeast China. Animals 2022, 12, 3290. https://doi.org/10.3390/ani12233290
Zhao D, Liu Z, Tan M, Wang Y, Dai W, Ge J, Feng L. Molecular Dietary Analysis of Three Sympatric Mustelidae in Northeast China. Animals. 2022; 12(23):3290. https://doi.org/10.3390/ani12233290
Chicago/Turabian StyleZhao, Dong, Zhihui Liu, Mengyu Tan, Yue Wang, Wenqian Dai, Jianping Ge, and Limin Feng. 2022. "Molecular Dietary Analysis of Three Sympatric Mustelidae in Northeast China" Animals 12, no. 23: 3290. https://doi.org/10.3390/ani12233290