
Demographic Analysis of Shortfin Mako Shark (Isurus oxyrinchus) in the South Pacific Ocean


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
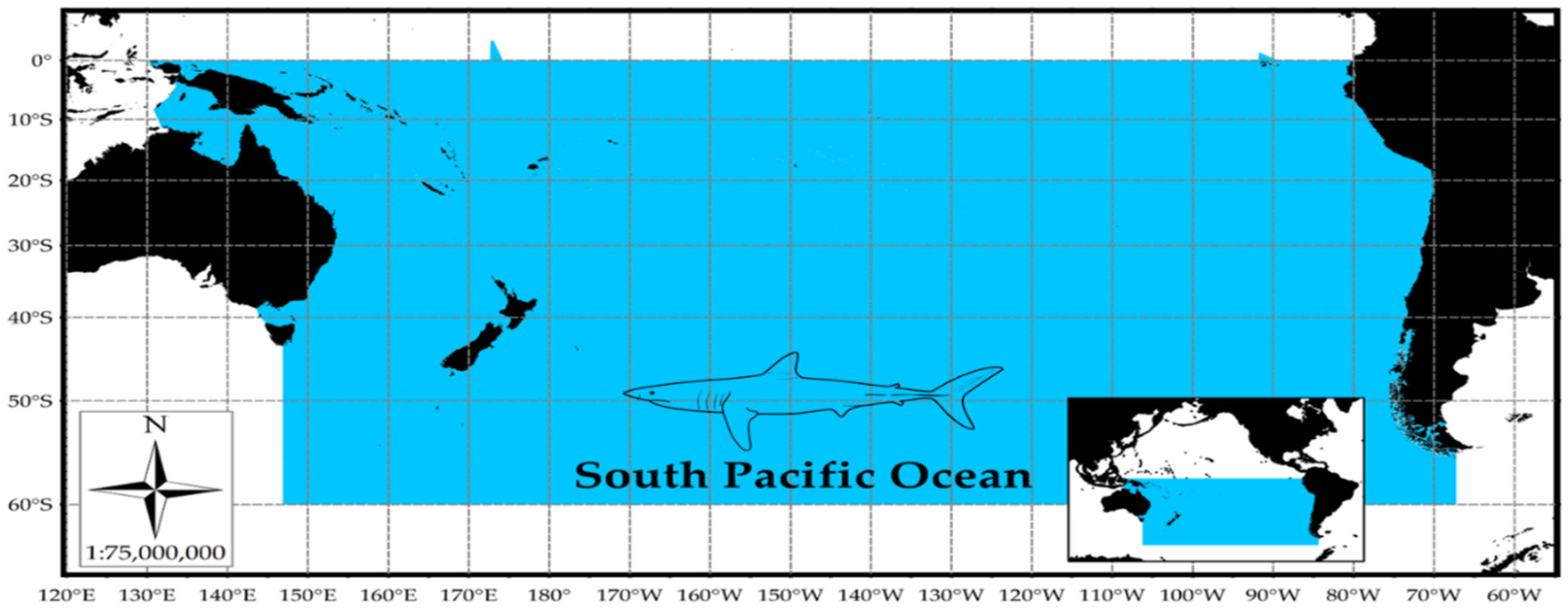
2.1. Study Area and Biological Parameters
2.2. Mako Shark Life History
2.3. Model Development
2.4. Mortality Estimation
2.5. Demographic Methods Accounting for Uncertainty
2.6. Size Limit Measures
- Scenario 1 (S1): Under natural conditions, F was set to 0 for all ages;
- Scenario 2 (S2): F was set to 80% of Ms by stage;
- Scenario 3 (S3): F was set to 60% of Ms by stage;
- Scenario 4 (S4): F was set to 40% of Ms by stage;
- Scenarios 5 (S5): F was set to 20% of Ms by stage;
- Scenario 6 (S6): With the protection of immature sharks, F was set to 80% of Ms by stage; F values of neonates, juveniles, and subadults of both sexes were set to 0;
- Scenario 7 (S7): With the protection of mature sharks, F was set to 80% of Ms by stage, and F values of adults of both sexes were set to 0;
- Scenario 8 (S8): With the protection of immature male sharks, F was set to 80% of Ms by stage, F values of male neonates, juveniles, and subadults were set to 0;
- Scenarios 9 (S9): With the protection of mature male sharks, F was set to 80% of Ms by stage, F values of male adults was set to 0;
- Scenario 10 (S10): With the protection of immature female sharks, F was set to 80% of Ms by stage; F values of female neonates, juveniles, and subadults were set to 0;
- Scenario 11 (S11): With the protection of mature female sharks, F was set to 80% of Ms by stage; F value of female adults was set to 0.
3. Results
3.1. Ms Estimation
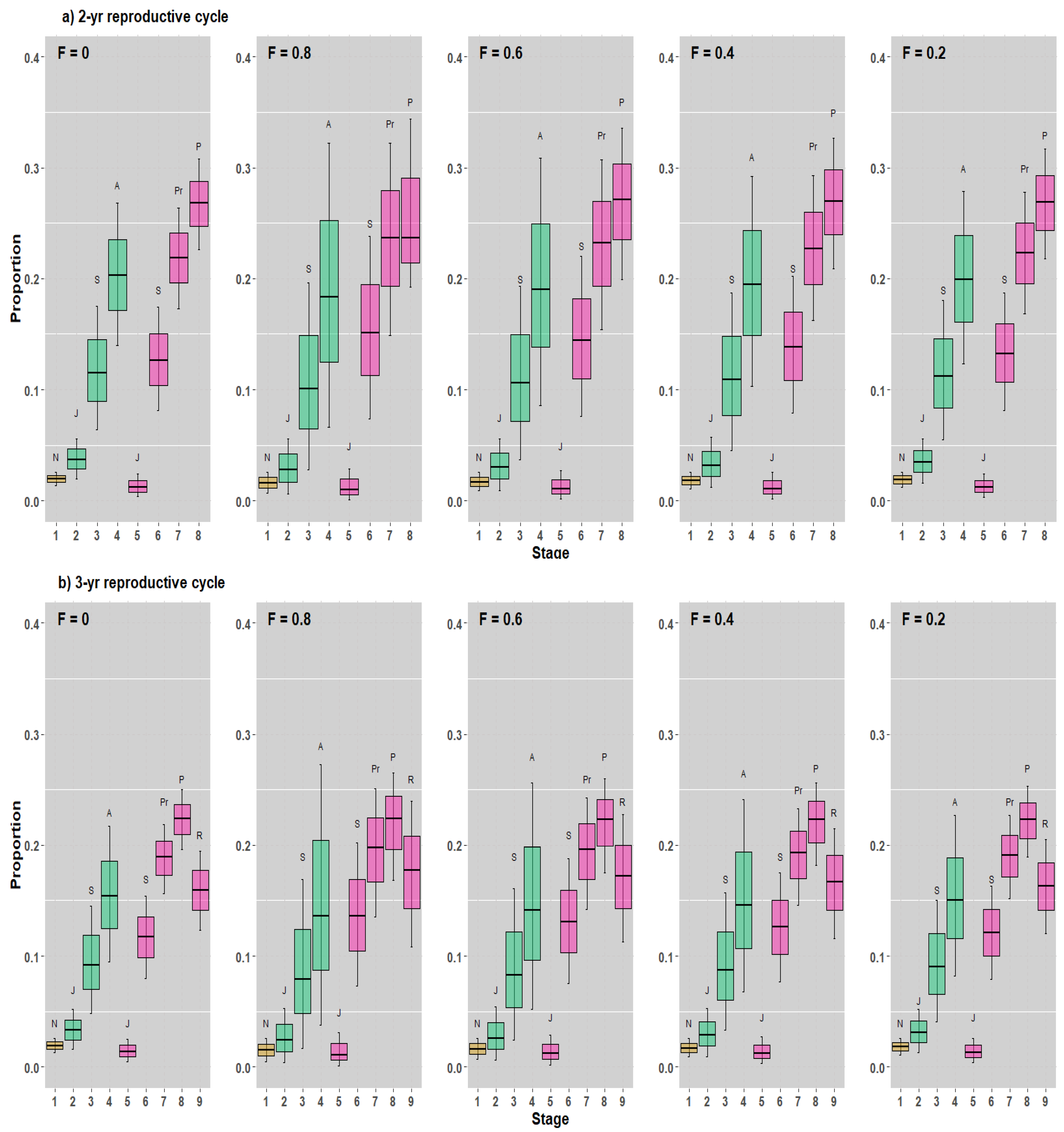
3.2. Demographic Analysis for the 2- and 3-Year Reproductive Cycle Models
3.3. SSD and RV
3.4. Size Limits Measures
4. Discussion
4.1. Ms
4.2. Demographic Analysis for Mako Shark Stock Assessment
4.3. Conservation Problems and Management Strategies for Mako Sharks
4.4. Life History Parameter Uncertainty and Their Limitation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dulvy, N.K.; Fowler, S.L.; Musick, J.A.; Cavanagh, R.D.; Kyne, P.M.; Harrison, L.R.; Carlson, J.K.; Davidson, L.N.K.; Fordham, S.V.; Francis, M.P.; et al. Extinction risk and conservation of the world’s sharks and rays. eLife 2014, 3, e00590. [Google Scholar] [CrossRef] [Green Version]
- Costello, C.; Ovando, D.; Clavelle, T.; Strauss, C.K.; Hilborn, R.; Melnychuk, M.C.; Branch, T.A.; Gaines, S.D.; Szuwalski, C.S.; Cabral, R.B.; et al. Global fishery prospects under contrasting management regimes. Proc. Natl. Acad. Sci. USA 2016, 113, 5125–5129. [Google Scholar] [CrossRef] [Green Version]
- Simpfendorfer, C.A.; Dulvy, N.K. Bright spots of sustainable shark fishing. Curr. Biol. 2017, 27, R97–R98. [Google Scholar] [CrossRef] [Green Version]
- Compagno, L.J.V. Sharks of the World: An Annotated and Illustrated Catalogue of Shark Species Known to Date. Bullhead, Mackerel and Carpet Sharks (Heterodontiformes, Lamniformes and Orectolobiformes); Food and Agriculture Organization of the United Nations: Rome, Italy, 2001; p. 269.
- Compagno, L.J.V.; Dando, M.; Fowler, S. A Field Guide to the Sharks of the World; Princeton University Press: Princeton, NJ, USA, 2005. [Google Scholar]
- Rigby, C.L.; Barreto, R.; Carlson, J.; Fernando, D.; Fordham, S.; Francis, M.P.; Jabado, R.W.; Liu, K.-M.; Marshall, A.; Pacoureau, N.; et al. Isurus oxyrinchus. The IUCN Red List of Threatened Species. 2019. Available online: https://www.iucnredlist.org/species/39341/2903170 (accessed on 21 February 2021).
- CITES (Convention on International Trade in Endangered Species of Wild Fauna and Flora). Updates on Decisions Made on Proposals to Amend Appendices I and II at CoP18. Decisions Made on Proposals to Amend Appendices I and II. 2019. Available online: https://cites.org/eng/updates_decisions_cop18_species_proposals (accessed on 21 February 2022).
- Stevens, J.D. Observations on reproduction in the Shortfin Mako, Isurus oxyrinchus. Copeia 1983, 1, 126–130. [Google Scholar] [CrossRef]
- Semba, Y.; Aoki, I.; Yokawa, K. Size at maturity and reproductive traits of shortfin mako, Isurus oxyrinchus, in the western and central North Pacific. Mar. Freshw. Res. 2011, 62, 20–29. [Google Scholar] [CrossRef]
- Mollet, H.F.; Cliff, G.; Pratt, H.L.; Stevens, J.D., Jr. Reproductive biology of the female shortfin mako, Isurus oxyrinchus, Rafinesque, 1810, with comments on the embryonic development of lamnoids. Fish. Bull. 2000, 98, 299–318. [Google Scholar]
- Francis, M.P.; Duffy, C. Length at maturity in three pelagic sharks (Lamna nasus, Isurus oxyrinchus, and Prionace glauca) from New Zealand. Fish. Bull. 2005, 103, 489–500. [Google Scholar]
- Joung, S.-J.; Hsu, H.-H. Reproduction and embryonic development of the Shortfin Mako, Isurus oxyrinchus, Rafinesque, 1810, in the northwestern Pacific. Zool. Stud. 2005, 44, 487–496. [Google Scholar]
- Murua, H.; Coelho, R.; Santos, M.N.; Arrizabalaga, H.; Yokawa, K.; Romanov, E.; Zhu, J.F.; Kim, Z.G.; Bach, P.; Chavance, P.; et al. Preliminary Ecological Risk Assessment (ERA) for shark species caught in fisheries managed by the Indian Ocean Tuna Commission (IOTC). IOTC 2012, 13, 16. [Google Scholar]
- Cortés, E.; Domingo, A.; Miller, P.; Forselledo, R.; Mas, F.; Arocha, F.; Campana, S.; Coelho, R.; Da Silva, C.; Holtzhausen, H.; et al. Expanded ecological risk assessment of pelagic sharks caught in Atlantic pelagic longline fisheries. Collect. Vol. Sci. Pap. ICCAT 2015, 71, 2637–2688. [Google Scholar]
- Griffiths, S.; Duffy, L.; Aires-da-Silva, A. A preliminar ecological risk assessment of the large-scale tuna longline fishery in the Eastern Pacific Ocean using productivity-susceptibility analysis. In Proceedings of the 8th Meeting of the Scientific Advisory Committee of the IATTC, La Jolla, CA, USA, 8–12 May 2017. [Google Scholar]
- Takeuchi, Y.; Senba, Y.; Nakano, H. Demographic analysis on Atlantic blue and shortfin mako sharks. Collect. Vol. Sci. Pap. ICCAT 2005, 58, 1157–1165. [Google Scholar]
- Chang, J.-H.; Liu, K.-M. Stock assessment of the Shortfin Mako shark, Isurus oxyrinchus, in the Northwest Pacific Ocean using per recruit and virtual population analyses. Fish. Res. 2009, 98, 92–101. [Google Scholar] [CrossRef]
- Tsai, W.-P.; Sun, C.-L.; Wang, S.-P.; Liu, K.-M. Evaluating the impacts of uncertainty on the estimation of biological reference points for the shortfin mako shark, Isurus oxyrinchus, in the Northwest Pacific Ocean. Mar. Freshw. Res. 2011, 62, 1383–1394. [Google Scholar] [CrossRef]
- Tsai, W.-P.; Sun, C.-L.; Punt, A.E.; Liu, K.-M. Demographic analysis of the shortfin mako shark, Isurus oxyrinchus, in the Northwestern Pacific using a two-sex stage-based matrix model. ICES J. Mar. Sci. 2014, 71, 1604–1618. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.-H.; Liu, K.-M. Demographic Analysis of the Shortfin Mako Shark, Isurus oxyrinchus, in the western North Pacific Using Stage-Based Models. J. Fish. Soc. Taiwan 2018, 45, 153–172. [Google Scholar]
- Semba, Y.; Yokoi, H.; Kai, M. Estimate of intrinsic rate of natural increase (r) of shortfin mako, Isurus oxyrinchus, based on life history parameters from Indian Ocean. In Proceedings of the IOTC—15th Working Party on Ecosystem and Bycatch, La Saline Les Bains, Reunion Island, France, 3–7 September 2019; IOTC: Victoria, Seychelles, 2019. WPEB15-20. [Google Scholar]
- Wu, X.-H.; Liu, S.Y.V.; Wang, S.-P.; Tsai, W.-P. Distribution patterns and relative abundance of shortfin mako shark caught by the Taiwanese large-scale longline fishery in the Indian Ocean. Reg. Stud. Mar. Sci. 2021, 44, 101691. [Google Scholar] [CrossRef]
- Large, K.; Neubauer, P.; Brouwer, S.; Kai, M. Stock Assessment of Southwest Pacific Shortfin Mako Shark; WCPFC: Colonia, Micronesia, 2022; (WCPFC-SC18-2022/SA-IP-13).
- Large, K.; Neubauer, P.; Brouwer, S. Input Data for the 2022 South Pacific Shortfin Mako Shark Stock Assessment; WCPFC: Colonia, Micronesia, 2022; (WCPFC-SC18-2022/SA-WP-02).
- Chan, R.W.K. Biological Studies on Sharks Caught Off the Coast of New South Wales. Ph.D. Thesis, University of New South Wales, Sydney, Australia, 2001; p. 323. [Google Scholar]
- Bishop, S.D.H.; Francis, M.P.; Duffy, C.; Montgomery, J.C. Age, growth, maturity, longevity and natural mortality of the shortfin mako shark, Isurus oxyrinchus, in New Zealand waters. Mar. Freshw. Res. 2006, 57, 143–154. [Google Scholar] [CrossRef]
- Cerna, F.; Licandeo, R. Age and growth of the shortfin mako, Isurus oxyrinchus, in the southeastern Pacific off Chile. Mar. Freshw. Res. 2009, 60, 394–403. [Google Scholar] [CrossRef]
- Tsai, W.-P.; Liu, K.-M.; Chang, Y.-J. Evaluation of Biological Reference Points for Conservation and Management of the Bigeye Thresher Shark, Alopias superciliosus, in the Northwest Pacific. Sustainability 2020, 12, 8646. [Google Scholar] [CrossRef]
- Zhu, J.; Chen, Y.; Dai, X.; Harley, S.J.; Hoyle, S.D.; Maunder, M.N.; Aires-da Silva, A.M. Implications of uncertainty in the spawner–recruitment relationship for fisheries management: An illustration using bigeye tuna, Thunnus obesus, in the eastern Pacific ocean. Fish. Res. 2012, 119, 89–93. [Google Scholar] [CrossRef]
- Tsai, W.-P.; Liu, K.-M.; Joung, S.J. Demographic analysis of the pelagic thresher shark, Alopias pelagicus, in the Northwestern Pacific using a stochastic stage-based model. Mar. Freshw. Res. 2010, 61, 1056–1066. [Google Scholar] [CrossRef]
- Tsai, W.-P.; Huang, C.-H. Data–limited approach to the management and conservation of the pelagic thresher shark in the Northwest Pacific. Conserv. Sci. Pract. 2022, 4, e12682. [Google Scholar] [CrossRef]
- Tsai, W.-P.; Wang, Y.-J.; Yamaguchi, A. Demographic analyses of the data limited silky shark population in the Indian Ocean using a two-sex stochastic matrix framework. J. Mar. Sci. Technol. 2019, 27, 55–63. [Google Scholar]
- Geng, Z.; Wang, Y.; Kingdong, R.; Zhu, J.; Dai, X. Demographic and harvest analysis for blue shark, Prionace glauca, in the Indian Ocean. Reg. Stud. Mar. Sci. 2021, 41, 101583. [Google Scholar] [CrossRef]
- Dowling, N.A.; Dichmont, C.M.; Haddon, M.; Smith, D.C.; Smith, A.D.M.; Sainsbury, K. Guidelines for developing formal harvest strategies for data-poor species and fisheries. Fish. Res. 2015, 171, 130–140. [Google Scholar] [CrossRef]
- Dowling, N.A.; Smith, A.D.M.; Smith, D.C.; Parma, A.M.; Dichmont, C.M.; Sainsbury, K.; Wilson, J.R.; Dougherty, D.T.; Cpoe, J.M. Generic solutions for data-limited fishery assessments are not so simple. Fish Fish. 2019, 20, 174–188. [Google Scholar] [CrossRef] [Green Version]
- Chrysafi, A.; Kuparinen, A. Assessing abundance of populations with limited data: Lessons learned from data-poor fisheries stock assessment. Environ. Rev. 2016, 24, 25–38. [Google Scholar] [CrossRef]
- Simpfendorfer, C.A. Demographic models: Life tables, matrix models and rebound potential. In Management Techniques for Elasmobranch Fisheries; Musick, J.A., Bonfil, R., Eds.; Fisheries Technical Paper (474); FAO: Rome, Italy, 2005; pp. 143–144. [Google Scholar]
- Smart, J.J.; Chin, A.; Tobin, A.; White, W.; Kumasi, B.; Simpfendorger, C.A. Stochastic demographic analyses of the silvertip shark, Carcharhinus albimarginatus, and the common blacktip shark, Carcharhinus limbatus, from the Indo-Pacific. Fish. Res. 2017, 191, 95–107. [Google Scholar] [CrossRef]
- Chen, P.; Yuan, W. Demographic analysis based on the growth parameter of sharks. Fish. Res. 2006, 78, 374–379. [Google Scholar] [CrossRef]
- Romine, J.G.; Musick, J.A.; Burgess, G.H. Demographic analyses of the dusky shark, Carcharhinus obscurus, in the Northwest Atlantic incorporating hooking mortality estimates and revised reproductive parameters. Environ. Biol. Fishes 2009, 84, 277–289. [Google Scholar] [CrossRef]
- Hisano, M.; Connolly, S.R.; Robbins, W.D. Population growth rates of reef sharks with and without fishing on the great barrier reef: Robust estimation with multiple models. PLoS ONE 2011, 6, e25028. [Google Scholar] [CrossRef] [Green Version]
- Tribuzio, C.A.; Kruse, G.H. Demographic and risk analyses of spiny dogfish, Squalus suckleyi, in the Gulf of Alaska using age and stage-based population models. Mar. Freshw. Res. 2011, 62, 1395–1406. [Google Scholar] [CrossRef]
- Cortés, E. Estimates of maximum population growth rate and steepness for blue sharks in the north and south Atlantic ocean. Collect. Vol. Sci. Pap. ICCAT 2016, 72, 1180–1185. [Google Scholar]
- Semba, Y.; Nakano, H.; Aoki, I. Age and growth analysis of the shortfin mako, Isurus oxyrinchus, in the western and central North Pacific Ocean. Environ. Biol. Fishes 2009, 84, 377–391. [Google Scholar] [CrossRef]
- Tsai, W.-P.; Liu, K.-M.; Punt, A.E.; Sun, C.-L. Assessing the potential biases of ignoring sexual dimorphism and mating mechanism in using a single-sex demographic model: The shortfin mako shark as a case study. ICES J. Mar. Sci. 2015, 72, 793–803. [Google Scholar] [CrossRef] [Green Version]
- Hüssy, K.; Coad, J.O.; Farrell, E.D.; Clausen, L.W.; Clarke, M.W. Sexual dimorphism in size, age, maturation, and growth characteristics of boarfish, Capros aper, in the Northeast Atlantic. ICES J. Mar. Sci. 2012, 69, 1729–1735. [Google Scholar] [CrossRef] [Green Version]
- Garcia, E.Q.; Zuanon, J. Sexual dimorphism in the electric knifefish, Gymnorhamphichthys rondoni, Rhamphichthyidae: Gymnotiformes. Acta Amazon. 2019, 49, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Faggion, S.; Vandeputte, M.; Vergnet, A.; Clota, F.; Blanc, M.; Sanchez, P.; Ruelle, F.; Allal, F. Sex dimorphism in European sea bass, Dicentrarchus labrax L.: New insights into sexrelated growth patterns during very early life stages. PLoS ONE 2021, 16, e0239791. [Google Scholar] [CrossRef]
- Caswell, H.; Weeks, D. Two-sex models: Chaos, extinction, and other dynamic consequences of sex. Am. Nat. 1986, 128, 41–47. [Google Scholar] [CrossRef]
- Lindström, J.; Kokkom, H. Sexual reproduction and population dynamics: The role of polygyny and demographic sex differences. Proc. Biol. Sci. 1998, 265, 483–488. [Google Scholar] [CrossRef] [Green Version]
- Gerber, L.R.; White, E.R. Two-sex matrix models in assessing population viability: When do male dynamics matter? J. Appl. Ecol. 2014, 51, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Cortés, E. Incorporating uncertainty into demographic modeling: Application to shark populations and their conservation. Conserv. Biol. 2002, 16, 1048–1062. [Google Scholar] [CrossRef]
- Yokoi, H.; Ijima, H.; Ohshimo, S.; Yokawa, K. Impact of biology knowledge on the conservation and management of large pelagic sharks. Sci. Rep. 2017, 7, 10619. [Google Scholar] [CrossRef]
- Brewster-Geisz, K.K.; Miller, T.J. Management of the sandbar shark, Carcharhinus plumbeus, implications of a stage-based model. Fish. Bull. 2000, 98, 236–249. [Google Scholar]
- Gilmore, G.R. Reproductive biology of lamnoid sharks. Environ. Biol. Fishes 1990, 38, 95–114. [Google Scholar] [CrossRef]
- Snelson, F.F.; Roman, B.L.; Burgess, G.H. The reproductive biology of pelagic elasmobranchs. In Sharks of the Open Ocean: Biology, Fisheries and Conservation; Camhi, M.D., Pikitch, E.K., Babcock, E.A., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2008; pp. 24–54. [Google Scholar]
- Duffy, C.; Francis, M.P. Evidence of summer parturition in shortfin mako, Isurus oxyrinchus, sharks from New Zealand waters. N. Z. J. Mar. Freshw. Res. 2001, 35, 319–324. [Google Scholar] [CrossRef] [Green Version]
- Caswell, H. Matrix Population Models: Construction, Analysis and Interpretation; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Caswell, H. Matrix Population Models. Construction, Analysis, and Interpretation, 2nd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2001; Volume 22, 772p. [Google Scholar]
- Frisk, M.G.; Miller, T.J.; Fogarty, M.J. The population dynamics of little skate, Leucoraja erinacea, winter skate, Leucoraja cellata, and barndoor skate, Dipturus laevis: Predicting exploitation limits using matrix analyses. ICES J. Mar. Sci. 2002, 59, 576–586. [Google Scholar] [CrossRef] [Green Version]
- Coale, A.J. The Growth and Structure of Human Populations: A Mathematical Investigation; Princeton University Press: Princeton, NJ, USA, 1972. [Google Scholar]
- Gotelli, N.J. A Primer of Ecology; Sinauer Associates: Sunderland, MA, USA, 1995; p. 206. [Google Scholar]
- Brodziak, J.; Ianelli, J.; Lorenzen, K.; Methot, R.D. Estimating Natural Mortality in Stock Assessment Applications; NOAA Technical Memorandum NMFS-F/SPO-119; U.S. Department of Commerce: Washington, DC, USA, 2011; p. 38.
- Kenchington, T.J. Natural mortality estimators for informationlimited fisheries. Fish Fish. 2014, 15, 533–562. [Google Scholar] [CrossRef]
- Punt, A.E.; Castillo-Jordán, C.; Hamel, O.S.; Cope, J.M.; Maunder, M.N.; Ianelli, J.N. Consequences of error in natural mortality and its estimation in stock assessment models. Fish. Res. 2021, 233, 105759. [Google Scholar] [CrossRef]
- Campana, S.; Joyce, W.; Marks, L.; Harley, S. Analytical Assessment of the Porbeagle Shark, Lamna nasus, Population in the Northwest Atlantic, with Estimates of Long-Term Sustainable Yield; Canadian Stock Assessment Research Document 2001/067; Canadian Science Advisory Secretariat: Ottawa, ON, Canada, 2001.
- Hoenig, J.M. Empirical use of longevity data to estimate mortality rates. Fish. Bull. 1983, 82, 898–903. [Google Scholar]
- Then, A.Y.; Hoenig, J.M.; Hall, N.G.; Hewitt, D.A. Evaluating the predictive performance of empirical estimators of natural mortality rate using information on over 200 fish species. ICES J. Mar. Sci. 2015, 72, 82–92. [Google Scholar] [CrossRef]
- Hamel, O.S. A method for calculating a meta-analytical prior for the natural mortality rate using multiple life history correlates. ICES J. Mar. Sci. 2015, 72, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.I.; Megrey, B.A. A revised Alverson and Carney model for estimating the instantaneous rate of natural mortality. Trans. Am. Fish. Soc. 2006, 135, 620–633. [Google Scholar] [CrossRef]
- Jensen, A.L. Beverton and Holt life history invariants result from optimal trade-off of reproduction and survival. Can. J. Fish. Aquat. Sci. 1996, 53, 820–822. [Google Scholar] [CrossRef]
- Frisk, M.G.; Miller, T.J.; Fogarty, M.J. Estimation and analysis of biological parameters in elasmobranch fishes: A comparative life history study. Can. J. Fish. Aquat. Sci. 2001, 58, 969–981. [Google Scholar] [CrossRef]
- Cubillos, L.A.; Alarcon, R.; Brante, A. Empirical estimates of natural mortality for the Chilean hake, Merluccius gayi, evaluation of precision. Fish. Res. 1999, 42, 147–153. [Google Scholar] [CrossRef]
- Caswell, H.; Brault, S.; Read, A.J.; Smith, T.D. Harbor porpoise and fisheries: An uncertainty analysis of incidental mortality. Ecol. Appl. 1998, 8, 1226–1238. [Google Scholar] [CrossRef]
- Cortés, E. A stochastic stage-based population model of the sandbar shark in the western North Atlantic. Am. Fish. Soc. Symp. 1999, 23, 115–136. [Google Scholar]
- Branstetter, S. Biological notes on the sharks of the north central Gulf of Mexico. Contrib. Mar. Sci. 1981, 24, 13–34. [Google Scholar]
- Stevens, J.D. The Biology and Ecology of the shortfin mako shark, Isurus oxyrinchus. In Sharks of the Open Ocean: Biology, Fisheries and Conservation; Camhi, M.D., Pikitch, E.K., Babcock, E.A., Eds.; Blackwell: Oxford, UK, 2008; pp. 87–91. [Google Scholar]
- Taniuchi, T. Some biological aspects of sharks caught by floating longlines-3. Reproduction. Rep. Jpn. Group Elasmobranch Stud. 1997, 33, 6–13. [Google Scholar]
- Mollet, H.F.; Cailliet, G.M. Comparative population demography of elasmobranches using life history tables, Leslie matrices and stage-based matrix models. Mar. Freshw. Res. 2002, 53, 503–516. [Google Scholar] [CrossRef]
- Taylor, C.C. Cod growth and temperature. ICES J. Mar. Sci. 1958, 23, 366–370. [Google Scholar] [CrossRef]
- Gompertz, B. On the nature of the function expressive of the law of human mortality and on a new mode of determining life contingencies. Philos. Trans. R. Soc. Lond. 1825, 115, 513–585. [Google Scholar]
- Ricker, W.E. Growth rates and models. In Fish Physiology; Hoar, W.S., Randall, D.J., Brett, J.R., Eds.; Academic Press: New York, NY, USA, 1979; Volume 8, pp. 677–743. [Google Scholar]
- Mucientes, G.R.; Queiroz, N.; Sousa, L.L.; Tarroso, P.; Sims, D.W. Sexual segregation of pelagic sharks and the potential threat from fisheries. Biol. Lett. 2009, 5, 156–159. [Google Scholar] [CrossRef] [Green Version]
- Nakano, H.; Nagasawa, K. Distribution of pelagic elasmobranchs caught by salmon research gillnets in the North Pacific. Fish. Sci. 1996, 62, 860–865. [Google Scholar] [CrossRef]
- Hood, G. PopTools. Pest Animal Control Co-Operative Research Center; CSIRO: Canberra, ACT, Austraila, 2004.
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Hoenig, J.M.; Then, A.Y.-H.; Babcock, E.A.; Hall, N.G.; Hewitt, D.A.; Hesp, S.A. The logic of comparative life history studies for estimating key parameters, with a focus on natural mortality rate. ICES J. Mar. Sci. 2016, 73, 2453–2467. [Google Scholar] [CrossRef]
- Brooks, E.N.; Pollock, K.H.; Hoenig, J.M.; Hern, W.S. Estimation of fishing and natural mortality from tagging studies on fisheries with two user groups. Can. J. Fish. Aquat. Sci. 1998, 55, 2001–2010. [Google Scholar] [CrossRef]
- Hewitt, D.A.; Lambert, D.M.; Hoenig, J.M.; Lipcius, R.N.; Bunnell, D.B.; Miller, T.J. Direct and indirect estimates of natural mortality for Chesapeake Bay blue crab. Trans. Am. Fish. Soc. 2007, 136, 1030–1040. [Google Scholar] [CrossRef] [Green Version]
- Hightower, J.E.; Jackson, J.R.; Pollock, K.H. Use of telemetry methods to estimate natural and fishing mortality of striped bass in Lake Gaston, North Carolina. Trans. Am. Fish. Soc. 2001, 130, 557–567. [Google Scholar] [CrossRef]
- Heupel, M.R.; Simpfendorfer, C.A. Estimation of mortality of juvenile blacktip sharks, Carcharhinus limbatus, within a nursery area using telemetry data. Can. J. Fish. Aquat. Sci. 2002, 59, 624–632. [Google Scholar] [CrossRef] [Green Version]
- Rudd, M.B.; Thorson, J.T.; Sagarese, S.R.; Kuparinen, A. Ensemble models for data-poor assessment: Accounting for uncertainty in life-history information. ICES J. Mar. Sci. 2019, 76, 870–883. [Google Scholar] [CrossRef]
- Murua, H.; Santiago, J.; Coelho, R.; Zudaire, I.; Neves, C.; Rosa, D.; Semba, Y.; Geng, Z.; Bach, P.; Arrizabalaga, H.; et al. Updated Ecological Risk Assessment (ERA) for Shark Species Caught in Fisheries Managed by the Indian Ocean Tuna Commission (IOTC). IOTC–2018–SC21–14_Rev_1. Available online: https://www.fao.org/3/bj492e/bj492e.pdf (accessed on 1 March 2021).
- Cortés, E.; Arocha, F.; Beerkircher, L.; Carvalho, F.; Domingo, A.; Heupel, M.; Holtzhausen, H.; Santos, M.N.; Ribera, M.; Simpfendorfer, C. Ecological risk assessment of pelagic sharks caught in Atlantic pelagic longline fisheries. Aquat. Living Resour. 2010, 23, 25–34. [Google Scholar] [CrossRef]
- ICCAT. 2012 Shortfin mako stock assessment and ecological risk assessment meeting. Collect. Vol. Sci. Pap. ICCAT 2013, 69, 1427–1570. [Google Scholar]
- Miller, T.E.X.; Inouye, B.D. Confronting two-sex demographic models with data. Ecology 2011, 92, 2141–2151. [Google Scholar] [CrossRef]
- Kokko, H.; Rankin, D.J. Lonely hearts or sex in the city? density-dependent effects in mating systems. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 319–334. [Google Scholar] [CrossRef] [Green Version]
- McAllister, M.K.; Pikitch, E.K.; Babcock, E.A. Using demographic methods to construct Bayesian priors for the intrinsic rate of increase in the Schaefer model and implications for stock rebuilding. Can. J. Fish. Aquat. Sci. 2001, 58, 1871–1890. [Google Scholar] [CrossRef]
- Tsai, W.-P.; Chang, Y.-Y.; Liu, K.-M. Development and testing of a Bayesian population model for the bigeye thresher shark, Alopias superciliosus, in an area subset of the Western North Pacific. Fish. Manag. Ecol. 2019, 26, 269–294. [Google Scholar] [CrossRef]
- Duarte, H.O.; Droguet, E.L.; Moura, M.C. Quantitative ecological risk assessment of shortfin mako shark, Isurus oxyrinchus: Proposed model and application example. Appl. Ecol. Environ. Res. 2018, 16, 3691–3709. [Google Scholar] [CrossRef]
- Liu, K.-M.; Wu, C.-B.; Joung, S.-J.; Tsai, W.-P.; Su, K.-Y. Multi-Model Approach on Growth Estimation and Association With Life History Trait for Elasmobranchs. Front. Mar. Sci. 2021, 8, 591692. [Google Scholar] [CrossRef]
- Pauly, D. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. ICES J. Mar. Sci. 1980, 39, 175–192. [Google Scholar] [CrossRef]
- Chen, S.; Watanabe, S. Age dependence of natural mortality coefficient in fish population dynamics. Nippon Suisan Gakk. 1989, 55, 205–208. [Google Scholar] [CrossRef]
- Peterson, I.; Wroblewski, J.S. Mortality rate of fishes in the pelagic ecosystem. Can. J. Fish. Aquat. Sci. 1984, 41, 1117–1120. [Google Scholar] [CrossRef]
- Ruckstuhl, K.E.; Cultton-Brock, T.H. Sexual segregation and the ecology of the two sexes. In Sexual Segregation in Vertebrates; Ruckstuhl, K.E., Neuhaus, P., Eds.; Cambridge University Press: Cambride, UK, 2006; pp. 3–8. [Google Scholar]
- Breed, M.D.; Moore, J. Animal Behavior, 2nd ed.; Elsevier: San Diego, CA, USA, 2015. [Google Scholar]
- Kraus, C.; Eberle, M.; Kappeler, P.M. The costs of risky male behaviour: Sex differences in seasonal survival in a small sexually monomorphic primate. Proc. R. Soc. B Biol. Sci. 2008, 275, 1635–1644. [Google Scholar] [CrossRef] [Green Version]
- Oro, D.; Torres, R.; Rodríguez, C.; Drummond, H. Climatic influence on demographic parameters of a tropical seabird varies with age and sex. Ecology 2010, 91, 1205–1214. [Google Scholar] [CrossRef] [Green Version]
- Jenouvrier, S.; Holland, M.; Stroeve, J.; Barbraud, C.; Weimerskirch, H.; Serreze, M.; Caswell, H. Effects of climate change on an emperor penguin population: Analysis of coupled demographic and climate models. Glob. Chang. Biol. 2012, 18, 2756–2770. [Google Scholar] [CrossRef] [Green Version]
- Vaughn, D.; Turnross, O.R.; Carrington, E. Sex-specific temperature dependence of foraging and growth of intertidal snails. Mar. Biol. 2014, 161, 75–87. [Google Scholar] [CrossRef]
- ICCAT. Report of the 2019 shortfin mako shark stock assessment update meeting. In Proceedings of the SMA SHK SA Intersessional Meeting, Madrid, Spain, 20–24 May 2019; Volume 76, pp. 1–77. [Google Scholar]
- de Bruyn, P. Report of the 2017 ICCAT Shortfin Mako Stock Assessment Meeting; International Commission for the Conservation of Atlantic Tunas: Madrid, Spain, 2018; Volume 74, pp. 1465–1561.
- Casey, J.G.; Kohler, N.E. Tagging studies on the shortfin mako shark, Isurus oxyrinchus, in the Western North Atlantic. Aust. J. Mar. Freshw. Res. 1992, 43, 45–60. [Google Scholar] [CrossRef]
- Frisk, M.G.; Miller, T.J.; Dulvy, N.K. Life histories and vulnerability to exploitation of elasmobranchs: Inferences from elasticity, perturbation and phylogenetic analyses. J. Northwest Atl. Fish. Sci. 2005, 35, 27–45. [Google Scholar] [CrossRef]
- Coelho, R.; Alpizar-Jara, R.; Erzini, K. Demography of a deep-sea lantern shark, Etmopterus spinax, caught in trawl fisheries of the northeastern Atlantic: Application of Leslie matrices with incorporated uncertainties. Deep-Sea Res. II Top. Stud. Oceanogr. 2015, 115, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Kinney, M.; Wells, R.; Kohin, S. Oxytetracycline age validation of an adult shortfin mako shark, Isurus oxyrinchus, after 6 years at liberty. J. Fish Biol. 2016, 89, 1828–1833. [Google Scholar] [CrossRef]
- Fogarty, M.J.; Rosenberg, A.A.; Sissenwine, M.P. Fisheries risk assessment: A case study of Georges Bank haddock. Environ. Sci. Technol. 1992, 26, 440–447. [Google Scholar] [CrossRef]
- Rose, K.A.; Cowam, J.H.; Winemiller, K.O.; Myers, R.A.; Hilborn, R. Compensatory density dependence in fish populations: Importance, controversy, understanding and prognosis. Fish Fish. 2002, 2, 293–327. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, R.I.; Deriso, R.B. Fishing in dangerous waters: Remarks on a controversial appeal to spawner-recruit theory for long-term impact assessment. Am. Fish. Soc. Monogr. 1988, 4, 232–243. [Google Scholar]
- Barnthouse, L.W.; Klauda, R.J.; Vaughan, D.S. Introduction to the monograph. In Science, Law, and Hudson River Power Plants: A Case Study in Environmental Impact Assessment; U.S. Department of Energy Office of Scientific and Technical Information: Washington, DC, USA, 1988; Volume 4, pp. 1–8. [Google Scholar]
- Macaluso, J. Red snapper season offered. In The Advocate; The Advocate: Baton Rouge, LA, USA, 1999. [Google Scholar]
- Heist, E.J.; Musick, J.A.; Graves, J.E. Genetic population structure of shortfin mako, Isurus oxyrinchus, inferred from restriction fragment length polymorphism analysis of mitochondrial DNA. Can. J. Fish. Aquat. Sci. 1996, 53, 583–588. [Google Scholar] [CrossRef]
- Schery, A.W.; Heist, E.J. Microsatellite markers for the shortfin mako and cross-species amplification in lamniformes. Conserv. Genet. 2002, 3, 459–461. [Google Scholar] [CrossRef]
- Francis, M.P.; Shivji, M.S.; Duffy, C.A.J.; Rogers, P.J.; Byrne, M.E.; Wetherbee, B.M.; Tindale, S.C.; Lyon, W.S.; Meyres, M.M. Oceanic nomad or coastal resident? Behavioural switching in the shortfin mako shark, Isurus oxyrinchus. Mar. Biol. 2019, 166, 5. [Google Scholar] [CrossRef]
- Sæther, B.E.; Coulson, T.; Grøtan, V.; Engen, S.; Altwegg, R.; Armitage, K.B.; Barbraud, C.; Becker, P.H.; Blumstein, D.T.; Dobson, F.S.; et al. How Life History Influences Population Dynamics in Fluctuating Environments. Am. Nat. 2013, 182, 743–759. [Google Scholar] [CrossRef]
- Zhou, S.; Deng, R.A.; Dunn, M.R.; Hoyle, S.D.; Lei, Y.; Williams, A.J. Evaluating methods for estimating shark natural mortality rate and management reference points using life-history parameters. Fish Fish. 2021, 23, 462–477. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| L∞ (cm) | K (Year−1) | t0 (Years) | L0 (cm) | Reference | |
|---|---|---|---|---|---|
| Male | 296.604 | 0.087 | −3.579 | 79.3 | [52] |
| Female | 325.293 | 0.076 | −3.182 | 70.0 | [52] |
| Sex | Stage-Class | Approximate Ages (Years) | Expected Stage Duration (Years) |
|---|---|---|---|
| Male | Neonates | 0–1 | 1 |
| Juveniles | 1–6 | 5 | |
| Subadults | 6–amat | 1–4 | |
| Adults | amat–amax | 12–24 | |
| Female | Neonates | 0–1 | 1 |
| Juveniles | 1–17 | 16 | |
| Subadults | 17–amat | 2–4 | |
| Adults | amat–amax | 15–26 | |
| Included | Pregnant adults | >21 | 1 |
| Parturient adults | >21 | 1 | |
| Resting adults * | >21 | 1 |
| ID | Method | Equation |
|---|---|---|
| M1 | Campana et al. [66] | |
| M2 | Hoeing [67] | |
| M3 | Then et al. [68] | |
| M3 | Then et al. [68] | |
| M5 | Hamel [69] | |
| M6 | Zhang and Megrey [70] | |
| M7 | Jensen [71] | |
| M8 | Hisano et al. [41] | |
| M9 | Frisk et al. [72] | |
| M10 | Cubillos et al. [73] | |
| M11 | Jensen [71] | |
| M12 | Frisk et al. [72] | |
| M13 | Hamel [69] | |
| M14 | Then et al. [68] |
| Sources of Uncertainty | Male | Female | Assumed Distribution |
|---|---|---|---|
| Fecundity | Triangle (8, 12.5, 18) | Triangle (8, 12.5, 18) | Triangular distribution |
| Natural mortality | ln (mean, SD) * | ln (mean, SD) * | Lognormal |
| Age at maturity | 7–10 years | 19–21 years | Uniform |
| Longevity | 22–31 years | 36–45 years | Uniform |
| Area | Model/Method | Sex | amat | amax | λ (yr−1) | Reference |
|---|---|---|---|---|---|---|
| Northwestern Atlantic | Age-structured matrix population model | F | 6, 7, 8 | 17 | 1.141 a (1.098–1.181) t | [52] |
| Atlantic | Age-structured matrix population model | F | 6, 7, 8 | 17 | * 1.076 a | [94] |
| North Atlantic | Bayesian surplus production model | F | 18 | 32 | * 1.060–1.061 y | [95] |
| South Atlantic | Bayesian surplus production model | F | 18 | 32 | * 1.060–1.064 y | [95] |
| North Pacific | Two-sex stage structured matrix population model | C | - | - | 1.078 b(2yr), 1.051 b(3yr) | [19] |
| M | 11–15 | 24–31 | ||||
| F | 18–21 | 31–41 | ||||
| North Pacific | Two-sex stage-structured matrix population model with polyandrous and polygynous | C | - | - | 1.010–1.082 c,y | [45] |
| M | 11–15 | 24–31 | ||||
| F | 18–21 | 31–41 | ||||
| Global | Two-sex age-structured matrix population model | C | - | - | * 1.107 d (1.007–1.374) x | [53] |
| M | 3 | 23 | ||||
| F | 17 | 38 | ||||
| Northwestern Pacific | Stage-based models | F | 19–22 | 42 | 1.059 e (1.023–1.098) z | [20] |
| Indian Ocean | Two-sex age-structured matrix population model | C | - | - | * 1.120 f (1.062–1.141) x | [21] |
| M | 7 | 23–29 | ||||
| F | 15 | 32–38 | ||||
| South Pacific | Two-sex stage-structured matrix population model | C | - | - | 1.098 g (2yr) (1.036–1.165) t 1.063 g (3yr) (1.008–1.119) t | This study |
| M | 7–10 | 22–31 | ||||
| F | 19–21 | 36–45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huynh, H.H.; Hung, C.-Y.; Tsai, W.-P. Demographic Analysis of Shortfin Mako Shark (Isurus oxyrinchus) in the South Pacific Ocean. Animals 2022, 12, 3229. https://doi.org/10.3390/ani12223229
Huynh HH, Hung C-Y, Tsai W-P. Demographic Analysis of Shortfin Mako Shark (Isurus oxyrinchus) in the South Pacific Ocean. Animals. 2022; 12(22):3229. https://doi.org/10.3390/ani12223229
Chicago/Turabian StyleHuynh, Hoang Huy, Chun-Yi Hung, and Wen-Pei Tsai. 2022. "Demographic Analysis of Shortfin Mako Shark (Isurus oxyrinchus) in the South Pacific Ocean" Animals 12, no. 22: 3229. https://doi.org/10.3390/ani12223229