
Preliminary Study on Species Diversity and Community Characteristics of Gamasid Mites on Small Mammals in Three Parallel Rivers Area of China

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Survey Sites
2.2. Gamasid Mites Collection and Identification
2.3. Species Diversity and Basic Community Structure Statistics
2.4. Species Abundance Distribution and Total Species Estimation
3. Results
3.1. Classification and Identification of Gamasid Mites and Their Small Mammal Hosts
3.2. Structural Characteristics and Species Diversity Changes of Gamasid Mite Communities
3.3. Species Abundance Distribution and Total Species Estimation of Gamasid Mite Community
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Taxonomic Checklist of Small Mammals Captured from Nine Survey Sites in Three Parallel Rivers Area of Northwest Yunnan, China (2001–2015)
| Orders, Families, Genera and Species of Small Mammal Hosts | Survey Sites | Individuals of Hosts |
| Order Rodentia | ||
| Ⅰ Family Muridae | ||
| 1 Rattus tanezumi (Temminck, 1844) | Gongshan, Jianchuan, Yulong, Weixi, Lushui, Fugong | 244 |
| 2 R. nitidus (Hodgson, 1845) | Gongshan, Jianchuan, Yulong, Weixi, Deqin, Lushui, Fugong | 129 |
| 3 R. norvegicus (Berkenhout, 1769) | Gongshan, Jianchuan, Yulong, Weixi | 136 |
| 4 R. andamanensis (Blyth, 1860) | Gongshan, Jianchuan, Deqin, Lushui | 37 |
| 5 Niviventer confucianus (Milne-Edwards, 1871) | Gongshan, Jianchuan, Xianggelila, Weixi, Deqin | 232 |
| 6 N. fulvescens (Gray, 1847) | Gongshan, Weixi, Fugong | 101 |
| 7 N. andersoni (Thomas, 1911) | Gongshan, Jianchuan, Lanping, Weixi | 35 |
| 8 N. excelsior (Thomas 1911) | Deqin | 2 |
| 9 Mus musculus Linnaeus, 1758 | Gongshan, Jianchuan | 9 |
| 10 M. caroli Bonhote, 1902 | Yulong | 1 |
| 11 Apodemus draco (Barrett-Hamiliton, 1900) | Gongshan, Weixi | 511 |
| 12 A. peninsulae (Thomas, 1907) | Gongshan, Weixi | 91 |
| 13 A. chevrieri (Milne-Edwards, 1868) | Gongshan, Jianchuan, Yulong, Lanping, Weixi, Xianggelila | 930 |
| 14 A. agrarius (Pallas, 1771) | Yulong, Xianggelila | 4 |
| 15 A. latronum Thomas, 1911 | Gongshan, Yulong, Xianggelila, Lanping, Deqin | 335 |
| 16 Berylmys bowersi (Anderson, 1879) | Gongshan, Weixi | 9 |
| 17 Micromys minutus (Pallas, 1771) | Jianchuan | 9 |
| 18 Vernaya fulva (G.M. Allen, 1927) | Weixi | 1 |
| 19 Bandicota indica (Bechstein, 1800) | Weixi | 1 |
| II Family Cricetidae | ||
| 20 Eothenomys miletus (Thomas, 1914) | Gongshan, Jianchuan, Yulong, Lanping, Weixi, Deqin | 552 |
| 21 E. eleusis (Thomas, 1911) | Gongshan, Lanping | 13 |
| 22 E. proditor Hinton, 1923 | Yulong, Xianggelila | 23 |
| 23 E. custos (Thomas, 1912) | Yulong, Lanping | 30 |
| Ⅲ Family Sciuridae | ||
| 24 Callosciurus quinquestriatus Anderson, 1871 | Lanping, Weixi | 6 |
| 25 Tamiops swinhoei (Milne-Edwards, 1874) | Gongshan | 3 |
| 26 Dremomys pernyi (Milne-Edwards, 1867) | Jianchuan, Weixi | 20 |
| 27 Rupestes forresti (Thomas, 1922) | Jianchuan | 13 |
| Ⅳ Family Pteromyidae | ||
| 28 Petaurista albiventer (Gray, 1834) | Gongshan, Weixi | 4 |
| 29 P. xanthotis (Milne-Edwards, 1872) | Weixi | 2 |
| 30 Trogopterus xanthipes (Milne-Edwards, 1867) | Gongshan, Weixi | 18 |
| 31 Belomys pearsonii (Gray, 1842) | Weixi | 1 |
| 32 Hylopetes alboniger (Hodgson, 1836) | Weixi | 4 |
| 33 Pteromys volans (Linnaeus, 1758) | Weixi | 11 |
| Order Eulipotyphla | ||
| Ⅴ Family Talpidae | ||
| 34 Scaptonyx fusicaudus (Milne-Edwrds, 1872) | Weixi | 1 |
| VI Family Soricidae | ||
| 35 Sorex excelsus (G.M. Allen, 1923) | Gongshan | 6 |
| 36 S. cylindricauda (Milne-Edwrds, 1872) | Weixi | 3 |
| 37 Episoriculus leucops (Hosfield, 1855) | Gongshan, Weixi | 7 |
| 38 Suncus murinus (Linnaeus, 1766) | Yulong, Xianggelila, Weixi | 14 |
| 39 Crocidura attenuata (Milne-Edwards, 1872) | Jianchuan, Yulong | 32 |
| 40 Anourosorex squamipes (Milne-Edwards, 1872) | Gongshan, Lanping, Weixi | 227 |
| Order Scandentia | ||
| VII Family Tupaiidae | ||
| 41 Tupaia belangeri (Wagner, 1841) | Yulong | 11 |
| Order Lagomorpha | ||
| VIII Family Ochotonidae | ||
| 42 Ochotona thibetana (Milne-Edwards, 1871) | Gongshan, Lanping, Weixi | 10 |
| 43 O. roylii (Ogilby, 1839) | Gongshan | 1 |
| Order Carnivora | ||
| IX Family Mustelidea | ||
| 44 Mustela kathiah Hodgson, 1835 | Yulong | 1 |
Appendix B. Taxonomic Checklist of Gamasid Mites Identified from Nine Survey Sites in Three Parallel Rivers Area of Northwest Yunnan, China (2001–2015)
| Families, Genera and Species of Gamasid Mites | Corresponding Small Mammal Hosts | Survey Sites |
| I Family Laelapidae Berlese 1892 | ||
| 1 Laelaps nuttalli Hirst, 1915 | Rattus tanezumi, Eothenomys miletus, R. nitidus, R. andamanensis, Apodemus chevrieri, R. norvegicus, A. draco, Niviventer confucianus, A. latronum, N. andersoni, Ochotona thibetana, E. custos, N. fulvescens, Mus musculus, Anourosorex squamipes, A. peninsulae, A. agrarius | Gongshan, Lushui, Fugong, Jianchuan, Yulong, Weixi, Xianggelila |
| 2 L. echidninus Berlese, 1887 | R. tanezumi, E. miletus, R. nitidus, R. andamanensis, N. confucianus, A. squamipes, R. norvegicus, N. fulvescens, Berylmys bowersi, A. chevrieri, Micromys minutus | Deqin, Lushui, Fugong, Gongshan, Jianchuan, Yulong, Weixi |
| 3 L. guizhouensis Gu et Wang, 1981 | A. chevrieri, M. musculus | Gongshan, Jianchuan, Yulong |
| 4 L. turkestanicus Lange, 1955 | N. confucianus, N. fulvescens, N. andersoni, A. chevrieri, N. excelsior, R. tanezumi, R. andamanensis, E. miletus, R. norvegicus, M. musculus, R. nitidus, Dremomys pernyi, M. caroli, B. bowersi, Petaurista albiventer, A. draco, A. peninsulae, Pteromys volans, P. xanthotis, Vernaya fulva, Trogopterus xanthipes | Deqin, Fugong, Gongshan, Jianchuan, Yulong, Lanping, Weixi, Xianggelila |
| 5 L. traubi Domrow, 1962 | N. andersoni, E. miletus, N. excelsior, R. nitidus, N. fulvescens, N. confucianus, A. chevrieri, A. latronum, A. draco, R. norvegicus, D. pernyi, P. volans, P. xanthotis, B. bowersi, Episoriculus leucops, Bandicota indica, | Deqin, Fugong, Gongshan, Jianchuan, Yulong, Lanping, Weixi, Xianggelila |
| 6 L. chini Wang et Li, 1965 | E. miletus, A. squamipes, N. confucianus, E. custos, E. eleusis, A. chevrieri, R. tanezumi, Suncus murinus, A. latronum, A. draco, E. leucops, M. minutus, R. norvegicus, E. proditor, A. peninsulae | Gongshan, Deqin, Jianchuan, Yulong, Lanping, Weixi, Xianggelila |
| 7 L. paucisetosa Gu et Wang, 1981 | N. fulvescens | Gongshan |
| 8 L. algericus Hirst, 1925 | M. caroli, | Yulong |
| 9 L. xingyiensis Gu et Wang, 1981 | M. minutus, | Jianchuan |
| 10 L. fukienensis Wang, 1963 | N. confucianus, N. fulvescens, N. andersoni, N. excelsior, M. musculus, R. nitidus, | Deqin, Fugong, Gongshan, Lanping |
| 11 L. liui Wang et Li, 1965 | B. bowersi | Gongshan |
| 12 L. jettmari Vitzthum, 1930 | A. chevrieri, E. miletus, M. minutus, R. norvegicus, A. latronum, E. eleusis, | Gongshan, Jianchuan, Yulong, Lanping, Xianggelila |
| 13 L. hongaiensis Grochovskaya et Nguen-Xuan-Xoe, 1961 | N. confucianus, | Gongshan |
| 14 L. jingdongensis Tian, Duan et Fang, 1990 | A. draco, E. miletus, A. chevrieri, A. latronum, E. eleusis, Callosciurus quinquestriatus, N. confucianus, A. peninsulae, R. norvegicus, D. pernyi, E. leucops | Gongshan, Deqin, Lanping, Weixi |
| 15 Haemolaelaps glasgowi (Ewing, 1925) | A. chevrieri, A. squamipes, N. fulvescens, P. albiventer, R. norvegicus, E. miletus, M. musculus, E. proditor | Gongshan, Jianchuan, Weixi, Xianggelila |
| 16 H. casalis (Berlese, 1887) | A. chevrieri, N. confucianus, E. miletus, T. xanthipes, P. volans, | Gongshan, Weixi |
| 17 H. petauristae Gu et Wang, 1980 | T. xanthipes | Gongshan |
| 18 H. anomalis Wang, Liao et Lin, 1981 | B. bowersi | Gongshan |
| 19 Androlaelaps singularis Wang et Li, 1965 | A. chevrieri, A. squamipes, R. nitidus, R. norvegicus, R. andamanensis, S. murinus, | Fugong, Gongshan, Jianchuan, Yulong, Xianggelila |
| 20 Dipolaelaps anourosorecis (Gu et Wang 1981) | E. miletus, R. norvegicus, Crocidura attenuata, A. squamipes, A. draco, Sorex excelsus, N. andersoni, R. nitidus, E. leucops, O. roylii, A. chevrieri, R. andamanensis, B. bowersi, P. albiventer, E. custos, | Gongshan, Yulong |
| 21 D. jiangkouensis Gu, 1985 | N. confucianus, N. fulvescens, | Gongshan |
| 22 D. longisetosus Huang, 1985 | A. latronum | Deqin |
| 23 Tricholaelaps myonysognathus (Grochovskaya et Nguen-Xuan-Xoe, 1961) | R. tanezumi, | Lushui |
| 24 Hypoaspis pavlovskii (Bregetova, 1956) | N. andersoni, E. miletus, A. draco, N. confucianus, A. squamipes, A. chevrieri, R. tanezumi, N. fulvescens, R. nitidus, R. andamanensis, S. murinus, R. norvegicus, A. latronum, A. peninsulae | Gongshan, Lushui, Fugong, Jianchuan, Yulong, Lanping, Weixi, Xianggelila |
| 25 H. miles (Berlese, 1892) | R. tanezumi, A. squamipes, E. miletus, R. nitidus, A. chevrieri, | Gongshan, Jianchuan, Yulong, Xianggelila |
| 26 H. lubrica Voigts et Oudemans, 1904 | R. norvegicus, R. nitidus, A. latronum, R. tanezumi, N. fulvescens, A. chevrieri, C. attenuata, | Gongshan, Deqin, Lushui, Fugong, Yulong |
| 27 H. chianensis Gu, 1990 | R. andamanensis, A. chevrieri | Gongshan |
| 28 H. praesternalis Willmann, 1949 | A. squamipes, R. tanezumi, R. nitidus, | Gongshan, Lushui, Fugong |
| 29 H. concinna (Teng, 1982) | R. nitidus, R. tanezumi, R. andamanensis, N. fulvescens, | Lushui, Fugong, Gongshan |
| 30 H. aculeifer (Canestrini, 1884) | R. nitidus | Fugong |
| 31 H. ovatus Ma, Ning et Wei, 2003 | R. norvegicus, A. draco, A. squamipes, R. nitidus | Gongshan |
| 32 H. digitalis Teng, 1981 | A. squamipes | Gongshan |
| 33 H. tengi Gu et Bai, 1991 | A. squamipes, | Gongshan |
| 34 Haemogamasus oliviformis Teng et Pan, 1964 | A. draco, A. squamipes, E. miletus, E. eleusis, O. thibetana, A. chevrieri, R. nitidus, R. andamanensis, N. fulvescens, A. latronum, N. confucianus, A. peninsulae, R. norvegicus, D. pernyi, C. quinquestriatus, E. custos, A. agrarius | Gongshan, Deqin, Jianchuan, Yulong, Lanping, Weixi, Xianggelila |
| 35 H. dorsalis Teng et Pan, 1964 | E. custos, A. chevrieri | Yulong, Lanping |
| 36 H. gongshanensis Tian et Gu, 1989 | E. miletus, A. squamipes, A. latronum, A. draco, N. confucianus, O. thibetana | Gongshan, Deqin, Weixi |
| 37 H. monticola Wang et Li, 1965 | A. draco, A. squamipes, R. andamanensis, N. fulvescens, R. nitidus, N. confucianus, S. murinus, A. chevrieri, E. custos, A. latronum, S. excelsus | Fugong, Gongshan, Yulong, Lanping, Weixi, Xianggelila |
| 38 H. sexsetosus Guo et Gu, 1993 | N. fulvescens, E. miletus, T. xanthipes | Gongshan, Jianchuan |
| 39 H. pontiger (Berlese, 1903) | R. norvegicus | Weixi |
| 40 H. multidentis Guo et Gu, 1997 | E. miletus | Weixi |
| 41 H. quadrisetatus Vitzthum, 1926 | A. draco, A. squamipes, N. confucianus, R. norvegicus | Gongshan, Weixi |
| 42 H. yunlongensis Gu et Fang, 1987 | E. miletus | Weixi |
| 43 H. paradauricus Teng et Pan, 1964 | A. chevrieri, N. confucianus | Lanping, Weixi |
| 44 H. sanxiaensis Liu et Ma, 2001 | A. squamipes, P. volans, T. xanthipes | Weixi |
| 45 H. hodosi Buiakova et Goncharova, 1961 | E. miletus, Scaptonyx fusicaudus | Weixi |
| 46 H. trifurcisetus Zhou et Jiang, 1987 | R. nitidus, | Gongshan |
| 47 Eulaelaps stabularis Koch, 1836 | R. nitidus, N. fulvescens, R. tanezumi, E. proditor | Fugong, Lushui, Xianggelila |
| 48 E. shanghaiensis Wen, 1976 | A. draco, E. miletus, N. confucianus, A. chevrieri, A. latronum | Weixi, Gongshan, Jianchuan, Yulong, Xianggelila |
| 49 E. dremomydis Gu et Wang, 1984 | Rupestes forresti, D. pernyi, E. miletus, A. latronum, N. confucianus, A. peninsulae | Jianchuan, Lanping, Weixi |
| 50 E. substabularis Yang et Gu, 1986 | A. draco, A. squamipes, R. tanezumi, A. latronum, A. peninsulae, E. miletus, R. norvegicus, R. nitidus, N. fulvescens, A. chevrieri, A. agrarius, E. eleusis, O. thibetana, N. confucianus, T. xanthipes, Hylopetes alboniger | Gongshan, Deqin, Fugong, Jianchuan, Yulong, Lanping, Weixi |
| 51 E. huzhuensis Yang et Gu, 1985 | A. latronum, R. norvegicus, R. nitidus, A. chevrieri, A. draco, E. miletus, N. confucianus, A. peninsulae, S. cylindricauda, E. leucops, P. volans, T. xanthipes, Belomys pearsonii | Deqin, Fugong, Gongshan, Lushui, Weixi |
| 52 E. silvestris Zhou, 1981 | R. tanezumi, | Fugong |
| 53 E. dongfangis Wen, 1976 | R. nitidus | Fugong |
| 54 Gymnolaelaps sinensis Wang, Zhou et Ji, 1991 | A. chevrieri | Weixi |
| 55 G. weishanensis Gu et Guo, 1997 | N. fulvescens, A. chevrieri, | Gongshan, Jianchuan, Yulong |
| 56 Cosmolaelaps retirugi (Ma, Yang et Zhang, 2004) | A. squamipes, R. norvegicus, E. miletus | Gongshan, Weixi |
| 57 C. yeruiyuae Ma, 1995 | A. squamipes, R. tanezumi, R. nitidus, N. fulvescens, | Gongshan, Lushui, Fugong |
| 58 Hirstionyssus sunci Wang, 1962 | E. miletus, A. squamipes, A. chevrieri, R. norvegicus, R. tanezumi, R. nitidus, N. confucianus, A. latronum, C. attenuata, Mustela kathiah, Tupaia belangeri, A. draco, A. agrarius, R. andamanensis, | Gongshan, Deqin, Fugong, Jianchuan, Yulong, Lanping, Weixi, Xianggelila |
| 59 H. callosciuri Bregetova et Grokhovskaya, 1961 | Tamiops swinhoei | Gongshan |
| 60 H. neosinicus Teng et Pan, 1962 | A. draco, E. miletus, N. confucianus | Weixi |
| 61 H. qinghaiensis Gu et Yang, 1986 | E. miletus | Weixi |
| 62 H. microti Hsu et Ma, 1964 | E. miletus | Weixi |
| 63 H. isabellinus (Oudemans, 1913) | A. draco, E. miletus, A. chevrieri | Weixi |
| 64 H. musculi (Johnston, 1849) | A. latronum | Deqin |
| II Family Dermanyssidae Kolenati 1859 | ||
| 65 Liponyssoides muris (Hirst, 1913) | R. norvegicus, T. belangeri | Jianchuan, Yulong |
| III Family Macronyssidae Oudemans 1996 | ||
| 66 Ornithonyssus bacoti (Hirst, 1913) | R. tanezumi, R. nitidus, R. norvegicus, R. andamanensis, A. draco, N. confucianus, A. chevrieri, A. peninsulae, N. andersoni | Deqin, Lushui, Fugong, Gongshan, Weixi |
| IV Family Aceosejidae Baker et Wharton 1952 | ||
| 67 Lasioseius medius Gu et Guo, 1994 | A. squamipes, E. miletus, E. custos | Gongshan, Yulong |
| 68 L. trifurcipilus Gu et Guo, 1996 | A. chevrieri | Xianggelila |
| 69 L. qinghaiensis Wang et Li, 2001 | A. squamipes, E. miletus, R. nitidus | Gongshan |
| 70 L. multispathus Gu et Huang, 1990 | A. squamipes | Gongshan |
| 71 L. liaohaorongae Ma, 1996 | A. squamipes | Gongshan |
| 72 L. chenpengi Ma et Yin, 1999 | A. squamipes | Gongshan |
| V Family Ameroseiidae Evans 1963 | ||
| 73 Ameroseius taoerhensis Ma, 1995 | A. latronum | Deqin |
| 74 Sinoseius lobatus Bai et Gu, 1995 | E. miletus, D. pernyi | Weixi |
| VI Family Parasitidae Oudemans 1901 | ||
| 75 Parasitus consanguineus Oudemans et Voigts, 1904 | A. draco | Gongshan |
| 76 P. wangdunqingi Ma, 1995 | A. squamipes | Gongshan |
| VII Family Parholaspidae Evans 1956 | ||
| 77 Gamasholaspis eothenomydis Gu, 1984 | A. squamipes | Gongshan |
| VIII Family Macrochelidae Vitzthum, 1930 | ||
| 78 Macrocheles liguizhenae Ma, 1996 | A. draco, A. squamipes, S. excelsus, E. miletus, R. nitidus | Gongshan, Fugong |
| 79 M. muscaedomesticae (Scopoli, 1772) | A. draco, A. squamipes, E. miletus, R. norvegicus | Gongshan |
| IX Family Pachylaelaptidae Berlese 1888 | ||
| 80 Pachylaelaps badongensis (Liu et Ma, 2003) | A. draco | Weixi |
| 81 P. nuditectus Ma et Yin, 2000 | A. latronum | Deqin |
| X Family Blattisocidae Garman, 1948 | ||
| 82 Proctolaelaps pygmaeus (Muller, 1859) | R. tanezumi, N. confucianus, N. fulvescens, A. draco, E. miletus, A. peninsulae, R. norvegicus, P. xanthotis, R. nitidus, A. chevrieri, M. musculus, C. attenuata, A. agrarius | Lushui, Fugong, Gongshan, Jianchuan, Yulong, Weixi, Xianggelila |
References
- Yin, S.G.; Bei, N.X.; Chen, W.P. Soil Gamasida from Northeast China; China Agricultural Press: Beijing, China, 2013. (In Chinese) [Google Scholar]
- Xiang, R.; Guo, X.G.; Zhao, C.F.; Fan, R.; Mao, K.Y.; Zhang, Z.W.; Huang, X.B. Infestation and distribution of gamasid mites on Himalayan field rat (Rattus nitidus) in Yunnan Province of Southwest China. Biologia 2021, 76, 1763–1773. [Google Scholar] [CrossRef]
- Huang, L.Q.; Guo, X.G.; Speakman, J.R.; Dong, W.G. Analysis of gamasid mites (Acari: Mesostigmata) associated with the Asian house rat, Rattus tanezumi (Rodentia: Muridae) in Yunnan Province, southwest China. Parasitol. Res. 2013, 112, 1967–1972. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.F.; Teng, K.F. Economic Insect Fauna of China Fasc. 40 Acari: Dermanyssoidese; Science Press: Beijing, China, 1993. (In Chinese) [Google Scholar]
- Rosen, S.; Yeruham, I.; Braverman, Y. Dermatitis in humans associated with the mites Pyemotes tritici, Dermanyssus gallinae, Ornithonyssus bacoti and Androlaelaps casalis in Israel. Med. Vet. Entomol. 2002, 16, 442–444. [Google Scholar] [CrossRef] [PubMed]
- George, D.R.; Finn, R.D.; Graham, K.M.; Mul, M.F.; Maurer, V.; Moro, C.V.; Sparagano, O.A. Should the poultry red mite Dermanyssus gallinae be of wider concern for veterinary and medical science? Parasite Vector 2015, 8, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, W.; Fölster-Holst, R. Tropical rat mites (Ornithonyssus bacoti)—Serious ectoparasites. J. Dtsch. Dermatol. Ges. 2009, 7, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Lopatina Iu, V.; Petrova, A.D.; Timoshkov, V.V. The gamasid mites (Parasitiformes: Mesostigmata) of small mammals from undeveloped land in Moscow. Parazitologiia 1998, 32, 118–128. [Google Scholar] [PubMed]
- Lucky, A.W.; Sayers, C.; Argus, J.D.; Lucky, A. Avian mite bites acquired from a new source--pet gerbils: Report of 2 cases and review of the literature. Arch. Dermatol. 2001, 137, 167–170. [Google Scholar]
- Goldman, L. Lichen urticatus syndrome as a manifestation of sensitivity to bites from various species of arthropods. Arch. Derm. Syphilol. 1948, 58, 74–79. [Google Scholar] [CrossRef]
- Yin, P.W.; Guo, X.G.; Jin, D.C.; Fan, R.; Zhao, C.F.; Zhang, Z.W.; Huang, X.B.; Mao, K.Y. Distribution and Host Selection of Tropical Rat Mite, Ornithonyssus bacoti, in Yunnan Province of Southwest China. Animals 2021, 11, 110. [Google Scholar] [CrossRef]
- Yin, P.W.; Guo, X.G.; Jin, D.C.; Song, W.Y.; Zhang, L.; Zhao, C.F.; Fan, R.; Zhang, Z.W.; Mao, K.Y. Infestation and Seasonal Fluctuation of Gamasid Mites (Parasitiformes: Gamasida) on Indochinese Forest Rat, Rattus andamanensis (Rodentia: Muridae) in Southern Yunnan of China. Biology 2021, 10, 1297. [Google Scholar] [CrossRef]
- Reeves, W.K.; Loftis, A.D.; Szumlas, D.E.; Abbassy, M.M.; Helmy, I.M.; Hanafi, H.A.; Dasch, G.A. Rickettsial pathogens in the tropical rat mite Ornithonyssus bacoti (Acari: Macronyssidae) from Egyptian rats (Rattus spp.). Exp. Appl. Acarol. 2007, 41, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.J.; Tesh, R.B. The role of mites in the transmission and maintenance of Hantaan virus (Hantavirus: Bunyaviridae). J. Infect. Dis. 2014, 210, 1693–1699. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.C.; Wang, L.; Wang, S.; Zhu, L.; Dong, L.Y.; Zhang, Z.T.; Hao, B.; Yang, F.; Liu, W.B.; Deng, Y.; et al. Meteorological factors affect the epidemiology of hemorrhagic fever with renal syndrome via altering the breeding and hantavirus-carrying states of rodents and mites: A 9 years’ longitudinal study. Emerg. Microbes Infect. 2017, 6, e104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhu, J.; Tao, K.; Wu, G.; Guo, H.; Wang, J.; Zhang, J.; Xing, A. Proliferation and location of Hantaan virus in gamasid mites and chigger mites, a molecular biological study. Natl. Med. J. China 2002, 82, 1415–1419. (In Chinese) [Google Scholar]
- Renz, A.; Wenk, P. Intracellular development of the cotton-rat filaria Litomosoides carinii in the vector mite Ornithonyssus bacoti. T. Roy. Soc. Trop. Med. H. 1981, 75, 166–168. [Google Scholar] [CrossRef]
- Xi, Y.F.; Xu, J.J.; Ren, Y.F.; Yuan, Y.Z.; Tao, G.Y.; Zhao, M.L.; Xu, Y.X. Drug screening with the cotton-rat model of filariasis. Acta Pharm. Sin. 1979, 14, 455–460. (In Chinese) [Google Scholar]
- Ming, Q.Z.; Shi, Z.T. The tentative inquiry on the formation time in the region of three parallel rivers. Yunnan Geog. Environ. Res. 2006, 18, 1–4. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, R.Z. Geological events and mammalian distribution in China. Acta Zool. Sin. 2002, 48, 141–153. [Google Scholar]
- Wu, S.H.; Dai, E.F.; He, D.M. Major Research Perspectives on Environmental and Developmental Issues for the Longitudinal Range-Gorge Region (LRGR) in Southwestern China. Prog. Geog. 2005, 24, 31–40. (In Chinese) [Google Scholar] [CrossRef]
- Ma, C.L.; Robert K, M.; Chen, W.Y.; Zhou, Z.K. Plant diversity and priority conservation areas of Northwestern Yunnan, China. Biodivers. Conserv. 2007, 16, 757–774. [Google Scholar] [CrossRef]
- Moseley, R.K. Historical Landscape Change in Northwestern Yunnan, China. Mt. Res. Dev. 2006, 26, 214–219. [Google Scholar] [CrossRef] [Green Version]
- Sherman, R.; Mullen, R.; Li, H.; Fang, Z.; Yi, W. Spatial patterns of plant diversity and communities in alpine ecosystems of the Hendguan Mountains, Northwest Yunnan, China. J. Plant Ecol. 2008, 1, 117–136. [Google Scholar] [CrossRef]
- Zhang, R.Z.; Zheng, D.; Yang, Q.Y.; Liu, Y.H. The Series of the Scientific Expedition to Hengduan Mountains, Qinghai-Xizang Plateau. Physical Geography of Hengduan Mountains; Science Press: Beijing, China, 1997. (In Chinese) [Google Scholar]
- Lin, S.; Wu, R.; Hua, C.; Ma, J.; Wang, W.; Yang, F.; Wang, J. Identifying local-scale wilderness for on-ground conservation actions within a global biodiversity hotspot. Sci. Rep. 2016, 6, 25898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diao, Y.; Wang, J.; Yang, F.; Wu, W.; Zhou, J.; Wu, R. Identifying optimized on-the-ground priority areas for species conservation in a global biodiversity hotspot. J. Environ. Manag. 2021, 290, 112630. [Google Scholar] [CrossRef] [PubMed]
- Sherman, R.; Mullen, R. Alpine Ecosystems of Northwest Yunnan, China: An Initial Assessment for Conservation. J. Mountain Sci. 2007, 4, 181–192. [Google Scholar] [CrossRef]
- Dahmana, H.; Granjon, L.; Diagne, C.; Davoust, B.; Fenollar, F.; Mediannikov, O. Rodents as Hosts of Pathogens and Related Zoonotic Disease Risk. Pathogens 2020, 9, 202. [Google Scholar] [CrossRef] [Green Version]
- Gong, Z.D.; Wu, H.Y.; Duan, X.D.; Feng, X.G.; Zhang, Y.Z.; Liu, Q. The species diversity and distribution trends of small mammals in Hengduan Mountains Yunnan. Biodivers. Sci. 2001, 9, 73–79. (In Chinese) [Google Scholar]
- Quan, S.Y.; Yue, R.P.; Zhang, L.Y.; Lian, H.Y.; Zang, Y.H.; Bian, C.L.; Li, D.; Ju, J.K.; Gong, Z.D. The composition and spatial distribution of small mammals in the Hengduan Mountains of Yunnan, China. Chin. J. Vector Biol. Control 2010, 21, 16–22. (In Chinese) [Google Scholar]
- Gao, G.; Wang, B.; He, C.X.; Luo, X. Biodiversity of birds and mammals in alpine habitat of Mt. Gaoligong, Lushui County, Yunnan. Biodivers. Sci. 2017, 25, 332–339. (In Chinese) [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.Z.; Li, X.Y.; Song, W.Y.; Li, Q.; Onditi, K.; Khanal, L.; Jiang, X.L. Small mammal species richness and turnover along elevational gradient in Yulong Mountain, Yunnan, Southwest China. Ecol. Evol. 2020, 10, 2545–2558. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.E.; Lacher, T.E.; Mittermeier, R.A. Handbook of the Mammals of the World, vol 7, Rodents II; Lynx Ediciones: Barcelona, Spain, 2017. [Google Scholar]
- Smith, A.T.; Xie, Y.; Hoffmann, R.S.; Lunde, D.; MacKinnon, J.; Wilson, D.E.; Wozencraft, W.C. A Guide to the Mammals of China; Princeton University Press: Princeton, NJ, USA, 2008. [Google Scholar]
- Huang, W.J.; Chen, Y.X.; Wen, Y.X. Glires of China (Zhong Guo Nie Chi Lei); Fudan University Press: Shanghai, China, 1995. (In Chinese) [Google Scholar]
- Pan, Z.W.; Deng, G.F. Economic Insect Fauna of China. Fasc 17, Acari: Gamasina; Science Press: Beijing, China, 1980. (In Chinese) [Google Scholar]
- Evans, G.O.; Till, W.M. Mesostigmatic mites of Britain and Ireland (Chelicerata: Acari-Parasitiformes): An introduction to their external morphology and classification. Trans. Zool. Soc. Lond. 1979, 35, 139–262. [Google Scholar] [CrossRef]
- Radovsky, F.J. The Macronyssidae and Laelapidae (Acarina: Mesostigmata) Parasitic on Bats; University of California Publications in Entomology: Berkeley, CA, USA; Los Angeles, CA, USA, 1967. [Google Scholar]
- Chen, Y.L.; Guo, X.G.; Ren, T.G.; Zhang, L.; Fan, R.; Zhao, C.F.; Zhang, Z.W.; Mao, K.Y.; Huang, X.B.; Qian, T.J. Infestation and distribution of chigger mites on Chevrieri’s field mouse (Apodemus chevrieri) in Southwest China. Int. J. Parasitol. Parasites Wildl. 2022, 17, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Peng, P.Y.; Guo, X.G.; Ren, T.G.; Song, W.Y.; Dong, W.G.; Fan, R. Species diversity of ectoparasitic chigger mites (Acari: Prostigmata) on small mammals in Yunnan Province, China. Parasitol. Res. 2016, 115, 3605–3618. [Google Scholar] [CrossRef] [PubMed]
- Legendre, P. Studying beta diversity: Ecological variation partitioning by multiple regression and canonical analysis. J. Plant Ecol. 2007, 31, 976–981. (In Chinese) [Google Scholar] [CrossRef] [Green Version]
- Legendre, P. Studying beta diversity: Ecological variation partitioning by multiple regression and canonical analysis. J. Plant Ecol. 2008, 1, 3–8. (In English) [Google Scholar] [CrossRef] [Green Version]
- Guo, X.G.; Dong, W.G.; Men, X.Y.; Qian, T.J.; Wu, D.; Ren, T.G.; Qin, F.; Song, W.Y.; Yang, Z.H.; Fletcher, Q.E. Species Abundance Distribution of Ectoparasites on Norway Rats (Rattus norvegicus) from a Localized Area in Southwest China. J. Arthropod Borne Dis. 2016, 10, 192–200. [Google Scholar] [PubMed]
- Peng, P.Y.; Guo, X.G.; Jin, D.C.; Dong, W.G.; Qian, T.J.; Qin, F.; Yang, Z.H. Species abundance distribution and ecological niches of chigger mites on small mammals in Yunnan province, southwest China. Biologia 2017, 72, 1031–1040. [Google Scholar] [CrossRef]
- Baltanás, A. On the Use of Some Methods for the Estimation of Species Richness. Oikos 1992, 65, 484–492. [Google Scholar] [CrossRef]
- Guo, X.G.; Speakman, J.R.; Dong, W.G.; Men, X.Y.; Qian, T.J.; Wu, D.; Qin, F.; Song, W.Y. Ectoparasitic insects and mites on Yunnan red-backed voles (Eothenomys miletus) from a localized area in southwest China. Parasitol. Res. 2013, 112, 3543–3549. [Google Scholar] [CrossRef]
- Chen, Z.; Yang, X.J.; Liu, J.Z. Research report of medical ticks and mites in Hebei province. J. Med. Pest Control 2006, 238–240. (In Chinese) [Google Scholar]
- Ji, H.Q.; Feng, S.Q.; Liu, N.; He, Y.M.; Li, H.; Zhu, B.; Du, J.; Zhou, C.B. Species and geographical distribution of fleas and gamasid mites on the rat-shape animals in Chongqing city. Chin. J. Hyg. Insectic. Equip. 2012, 18, 413–415. (In Chinese) [Google Scholar] [CrossRef]
- Lu, M.G.; Jiang, Q.L.; Gong, Z.Y.; Ni, Q.X.; Ma, L.M. A list of Gamasid mites (Acari: Gamasina) in Zhejiang province. Chin. J. Vector Biol. Control 2017, 28, 269–273. (In Chinese) [Google Scholar]
- Tao, J.W.; Liu, Y.R.; Yang, Z.Q. Analysis on Fauna of Gamasid Mites in Hubei Province, China. Chin. J. Vector Biol. Control 2005, 16, 294–296. (In Chinese) [Google Scholar]
- Zhou, G.Z.; Wang, Z.; Huang, W.C.; Li, P.; Yin, G.Q.; Wen, Y.; Cheng, X.H.; Xue, J. Survey of gamasid mite species and evaluation of gamasid mite control efficacy in military camps in Shandong province, China. Chin. J. Vector Biol. Control 2013, 24, 535–537. (In Chinese) [Google Scholar]
- Zhou, S.H.; Deng, Y.Q.; Li, S.Y.; Wang, L.L. Supplementary records of Dermanyssoid mites (Acari: Parasitiformes) in Fujian province. Chin. J. Vector Biol. Control 2012, 23, 467–470. (In Chinese) [Google Scholar]
- Mašán, P. A revision of the family Ameroseiidae (Acari, Mesostigmata), with some data on Slovak fauna. ZooKeys 2017, 704, 1–228. [Google Scholar] [CrossRef] [Green Version]
- Vinarski, M.V.; Korallo-Vinarskaya, N.P. An annotated catalogue of the gamasid mites associated with small mammals in Asiatic Russia. The family Hirstionyssidae (Acari: Mesostigmata: Gamasina). Zootaxa 2020, 4838, 102–118. [Google Scholar] [CrossRef]
- Salmane, I.; Heldt, S. Predatory soil mites (Acari, Mesostigmata, Gamasina) from the Western Baltic Coast of Latvia. Acarologia 2001, 41, 295–301. [Google Scholar]
- He, J.; Lin, S.; Li, J.; Yu, J.; Jiang, H. Evolutionary history of zoogeographical regions surrounding the Tibetan Plateau. Commun. Biol. 2020, 3, 415. [Google Scholar] [CrossRef]
- Kai, H.; Jiang, X. Sky islands of southwest China. I: An overview of phylogeographic patterns. Chin. Sci. Bull. 2014, 59, 1–13. [Google Scholar]
- Wang, Y.; Si Tu, Q.; Li, Z.H. Relationship Between Regional Geographic Environment Characteristics and Soil and Water Loss of Northwest Yunnan Province. Bull. Soil Water Conserv. 2003, 23, 1–5+57. (In Chinese) [Google Scholar] [CrossRef]
- Cavarzere, V.; Roper, J.J.; Marchi, V.; Silveira, L.F. Geographical drivers of altitudinal diversity of birds in the Atlantic Forest. Biologia 2021, 76, 3275–3285. [Google Scholar] [CrossRef]
- Ghimire, A.; Rokaya, M.B.; Timsina, B.; Bílá, K.; Kindlmann, P. Diversity of birds recorded at different altitudes in central Nepal Himalayas. Ecol. Indic. 2021, 127, 107730. [Google Scholar] [CrossRef]
- Heaney, L.R. Small mammal diversity along elevational gradients in the Philippines: An assessment of patterns and hypotheses. Global Ecol. Biogeogr. 2001, 10, 15–39. [Google Scholar] [CrossRef]
- Koh, C.N.; Lee, P.F.; Lin, R.S. Bird species richness patterns of northern Taiwan: Primary productivity, human population density, and habitat heterogeneity. Divers. Distrib. 2010, 12, 546–554. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, X.; Luo, Z.F.; Lan, J.J.; Liu, Y.H. Elevational diversity gradients across seed plant taxonomic levels in the Lancang River Nature Reserve: Role of temperature, water and the mid-domain effect. J. For. Res. 2018, 29, 1121–1127. [Google Scholar] [CrossRef]
- Christy, M.M. Global analysis of reptile elevational diversity. Global Ecol. Biogeogr. 2010, 19, 541–553. [Google Scholar] [CrossRef]
- Xiang, R.; Guo, X.G.; Ren, T.G.; Zhao, C.F.; Fan, R.; Zhang, Z.W.; Mao, K.Y.; Peng, P.Y.; Huang, X.B.; Qian, T.J. Infestation and distribution of mites on the Yunnan red-backed vole (Eothenomys miletus) in Yunnan Province of southwest China between 2001 and 2015. Biologia 2021, 77, 61–68. [Google Scholar] [CrossRef]
- Li, B.; Guo, X.G.; Zhao, C.F.; Zhang, Z.W.; Fan, R.; Peng, P.Y.; Song, W.Y.; Ren, T.G.; Zhang, L.; Qian, T.J. Infestation of chigger mites on Chinese mole shrew, Anourosorex squamipes, in Southwest China and ecological analysis. Parasite 2022, 29, 39. [Google Scholar] [CrossRef]
- Eslami, A.; Yousefi, A.; Dowling, A.P.G. Prevalence of ectoparasites in black rat (Rattus rattus) from Mangrove forests of Qeshm Island, Iran. Comp. Clin. Path. 2018, 27, 1583–1586. [Google Scholar] [CrossRef]
- Peng, P.Y.; Guo, X.G.; Song, W.Y.; Hou, P.; Zou, Y.J.; Fan, R. Ectoparasitic chigger mites on large oriental vole (Eothenomys miletus) across southwest, China. Parasitol. Res. 2016, 115, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Fecchio, A.; Martins, T.F.; Bell, J.A.; De La Torre, G.M.; Pinho, J.B.; Weckstein, J.D.; Tkach, V.V.; Labruna, M.B.; Dias, R.I. Low host specificity and lack of parasite avoidance by immature ticks in Brazilian birds. Parasitol. Res. 2020, 119, 2039–2045. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Guo, X.G.; Song, W.Y.; Fan, R.; Zhao, C.F.; Mao, K.Y.; Zhang, Z.W.; Peng, P.Y.; Lin, H.; Dong, W.G.; et al. Infestation and distribution of chigger mites on Brown rat (Rattus norvegicus) in Yunnan Province, Southwest China. Trop. Biomed. 2021, 38, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Bhuyan, P.J.; Nath, A.J. Record of Tropical Rat Mite, Ornithonyssus bacoti (Acari: Mesostigmata: Macronyssidae) from Domestic and Peridomestic Rodents (Rattus rattus) in Nilgiris, Tamil Nadu, India. J. Arthropod Borne Dis. 2015, 10, 98–101. [Google Scholar] [PubMed]
- Heukelbach, J.; Feldmeier, H. Ectoparasitic Infestations. Curr. Infect. Dis. Rep. 2005, 7, 373–380. [Google Scholar] [CrossRef]
- McGill, B.J.; Etienne, R.S.; Gray, J.S.; Alonso, D.; Anderson, M.J.; Benecha, H.K.; Dornelas, M.; Enquist, B.J.; Green, J.L.; He, F.; et al. Species abundance distributions: Moving beyond single prediction theories to integration within an ecological framework. Ecol. Lett. 2007, 10, 995–1015. [Google Scholar] [CrossRef]
- Guo, X.G.; Qian, T.J.; Meng, X.Y.; Dong, W.G.; Shi, W.X.; Wu, D. Preliminary ananlysis of chigger communities associated with house rats (Rattus flavipectus) from six counties in Yunnan, China. Syst. Appl. Acarol. 2006, 11, 13–21. [Google Scholar]
- Engen, S. Heterogeneous communities with lognormal species abundance distribution: Species–area curves and sustainability. J. Theor. Biol. 2007, 249, 791–803. [Google Scholar] [CrossRef]
- Ding, F.; Jiang, W.L.; Guo, X.G.; Fan, R.; Zhao, C.F.; Zhang, Z.W.; Mao, K.Y.; Xiang, R. Infestation and Related Ecology of Chigger Mites on the Asian House Rat (Rattus tanezumi) in Yunnan Province, Southwest China. Korean J. Parasitol. 2021, 59, 377–392. [Google Scholar] [CrossRef]
- Matthews, T.J.; Whittaker, R.J. On the species abundance distribution in applied ecology and biodiversity management. J. Appl. Ecol. 2015, 52, 443–454. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order Names of Small Mammal Hosts | Individuals of Hosts | Number of Host Families | Number of Host Genera | Number of Host Species | Constituent Ratios Cr (%) |
|---|---|---|---|---|---|
| Rodentia | 3517 | 4 | 18 | 33 | 91.83 |
| Eulipotyphla | 290 | 2 | 6 | 7 | 7.57 |
| Scandentia | 11 | 1 | 1 | 1 | 0.29 |
| Lagomorpha | 11 | 1 | 1 | 2 | 0.29 |
| Carnivora | 1 | 1 | 1 | 1 | 0.03 |
| Total | 3830 | 9 | 27 | 44 | 100.00 |
| Family Names of Gamasid Mites | Number of Gamasid Mite Genera | Number of Gamasid Mite Species | Individuals of Gamasid Mites | Constituent Ratios of Gamasid Mite Individuals Cr (%) |
|---|---|---|---|---|
| Laelapidae | 11 | 64 | 24,853 | 95.41 |
| Dermanyssidae | 1 | 1 | 16 | 0.06 |
| Macronyssidae | 1 | 1 | 928 | 3.56 |
| Aceosejidae | 1 | 6 | 31 | 0.12 |
| Ameroseiidae | 2 | 2 | 4 | 0.02 |
| Parasitidae | 1 | 2 | 3 | 0.01 |
| Parholaspidae | 1 | 1 | 1 | 0.01 |
| Macrochelidae | 1 | 2 | 48 | 0.18 |
| Pachylaelaptidae | 1 | 2 | 11 | 0.04 |
| Blattisocidae | 1 | 1 | 153 | 0.58 |
| Total | 21 | 82 | 26,048 | 100.00 |
| Names of Dominant Hosts | Number of Hosts | Cr of Hosts | Individuals of Gamasid Mites | S | H | E | D |
|---|---|---|---|---|---|---|---|
| Apodemus chevrieri (Milne-Edwards, 1868) | 930 | 24.28% | 1775 | 30 | 2.50 | 0.73 | 0.12 |
| Eothenomys miletus (Thomas, 1914) | 552 | 14.41% | 877 | 34 | 1.79 | 0.51 | 0.36 |
| Apodemus draco (Barrett-Hamiliton, 1900) | 511 | 13.34% | 558 | 24 | 2.24 | 0.70 | 0.18 |
| Names of Dominant Host Species | Names of Dominant Gamasid Mite Species | Individuals of Total Gamasid Mites | Individuals of Dominant Gamasid Mite Species | Cr of Dominant Gamasid Mite Species |
|---|---|---|---|---|
| Apodemus chevrieri | Eulaelaps shanghaiensis Wen, 1976 | 1775 | 493 | 27.77% |
| Eothenomys miletus | Laelaps chini Wang et Li, 1965 | 877 | 509 | 58.04% |
| Apodemus draco | Laelaps jingdongensis Tian, Duan et Fang, 1990 | 558 | 207 | 37.10% |
| Altitude Gradients (m) | Individuals of Small Mammal Hosts | Individuals of Gamasid Mites | Statistics on Community Structure of Small Mammal Hosts | Statistics on Community Structure of Gamasid Mites | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| S | H | E | D | S | H | E | D | |||
| <1000 | 182 | 3702 | 3 | 0.39 | 0.36 | 0.81 | 13 | 0.85 | 0.33 | 0.55 |
| 1000–1500 | 214 | 7315 | 8 | 1.47 | 0.71 | 0.30 | 38 | 1.84 | 0.51 | 0.22 |
| 1500–2000 | 400 | 8710 | 16 | 1.91 | 0.69 | 0.20 | 41 | 1.16 | 0.31 | 0.52 |
| 2000–2500 | 367 | 1498 | 20 | 2.18 | 0.73 | 0.18 | 24 | 2.02 | 0.64 | 0.21 |
| 2500–3000 | 735 | 1304 | 22 | 1.67 | 0.54 | 0.33 | 28 | 2.59 | 0.78 | 0.10 |
| 3000–3500 | 1799 | 2954 | 32 | 2.17 | 0.63 | 0.16 | 42 | 2.61 | 0.70 | 0.10 |
| >3500 | 133 | 565 | 13 | 1.69 | 0.66 | 0.26 | 23 | 2.07 | 0.66 | 0.19 |
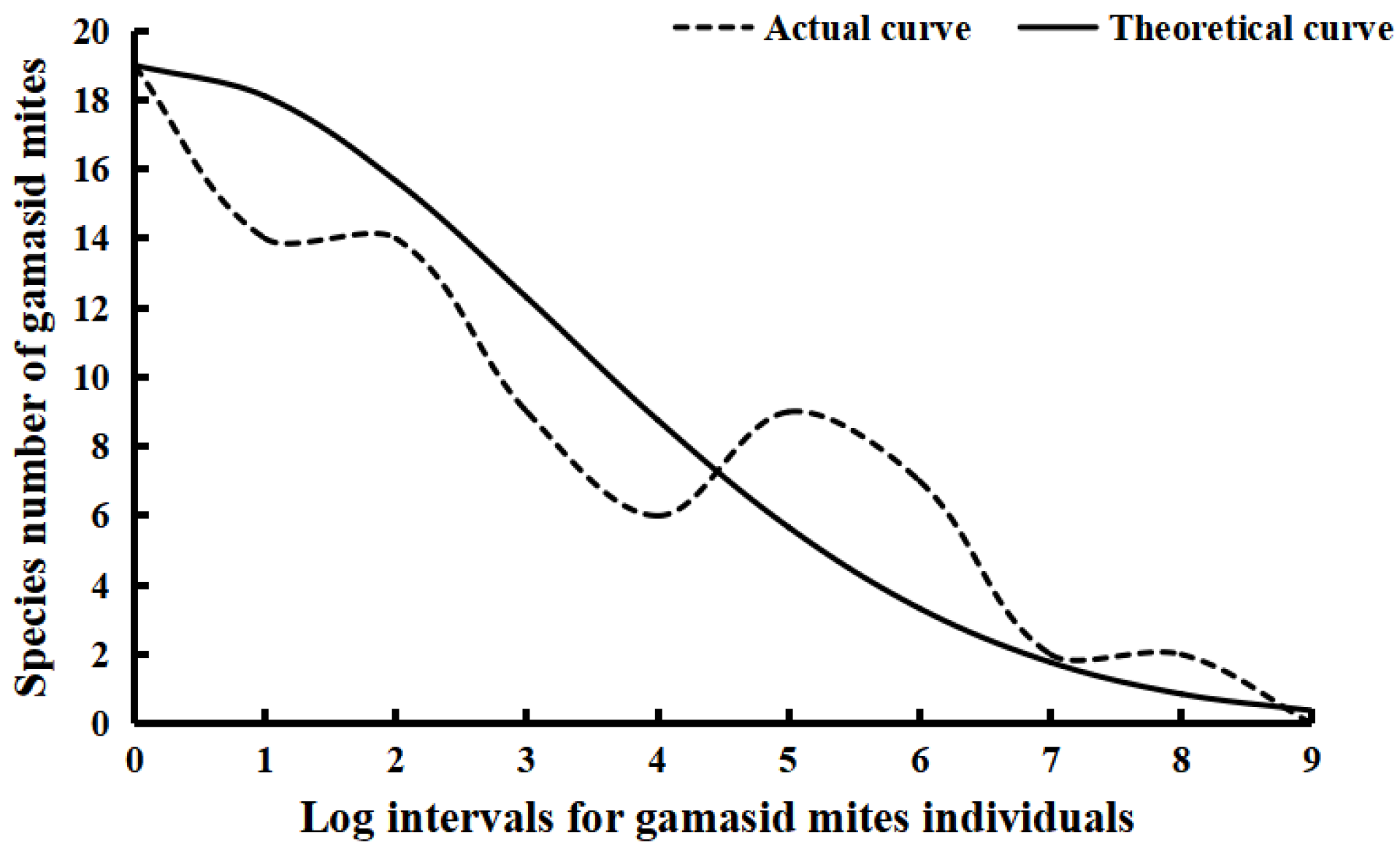
| Log Intervals Based on log3M | Individual Ranges of Gamasid Mites in Each Log Interval | Midpoint Values of Each Individual Range | Actual Gamasid Mite Species | Theoretical Gamasid Mite Species |
|---|---|---|---|---|
| 0 | 0–1 | 1 | 19 | 19.00 |
| 1 | 2–4 | 3 | 14 | 18.10 |
| 2 | 5–13 | 9 | 14 | 15.66 |
| 3 | 14–40 | 27 | 9 | 12.29 |
| 4 | 41–121 | 81 | 6 | 8.76 |
| 5 | 122–364 | 243 | 9 | 5.67 |
| 6 | 365–1093 | 729 | 7 | 3.33 |
| 7 | 1094–3280 | 2187 | 2 | 1.77 |
| 8 | 3281–9841 | 6561 | 2 | 0.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.-X.; Guo, X.-G.; Song, W.-Y.; Zhao, C.-F.; Zhang, Z.-W.; Fan, R.; Chen, T.; Lv, Y.; Yin, P.-W.; Jin, D.-C. Preliminary Study on Species Diversity and Community Characteristics of Gamasid Mites on Small Mammals in Three Parallel Rivers Area of China. Animals 2022, 12, 3217. https://doi.org/10.3390/ani12223217
Zhou J-X, Guo X-G, Song W-Y, Zhao C-F, Zhang Z-W, Fan R, Chen T, Lv Y, Yin P-W, Jin D-C. Preliminary Study on Species Diversity and Community Characteristics of Gamasid Mites on Small Mammals in Three Parallel Rivers Area of China. Animals. 2022; 12(22):3217. https://doi.org/10.3390/ani12223217
Chicago/Turabian StyleZhou, Juan-Xiu, Xian-Guo Guo, Wen-Yu Song, Cheng-Fu Zhao, Zhi-Wei Zhang, Rong Fan, Ting Chen, Yan Lv, Peng-Wu Yin, and Dao-Chao Jin. 2022. "Preliminary Study on Species Diversity and Community Characteristics of Gamasid Mites on Small Mammals in Three Parallel Rivers Area of China" Animals 12, no. 22: 3217. https://doi.org/10.3390/ani12223217