
Brain Organoids to Evaluate Cellular Therapies

 , , , , , and
, , , , , and 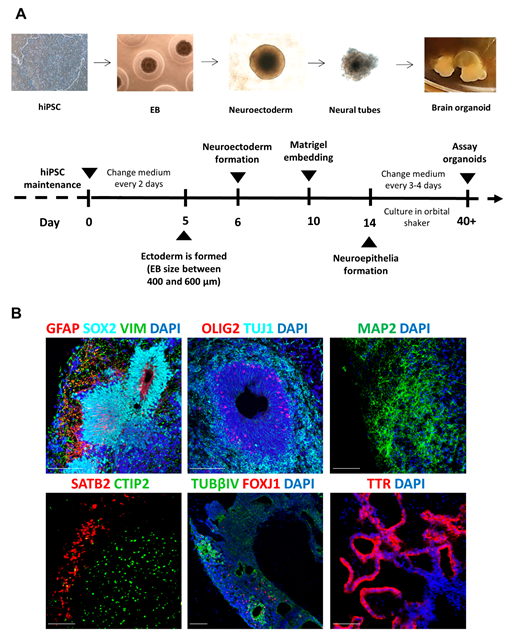{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Organoid Generation
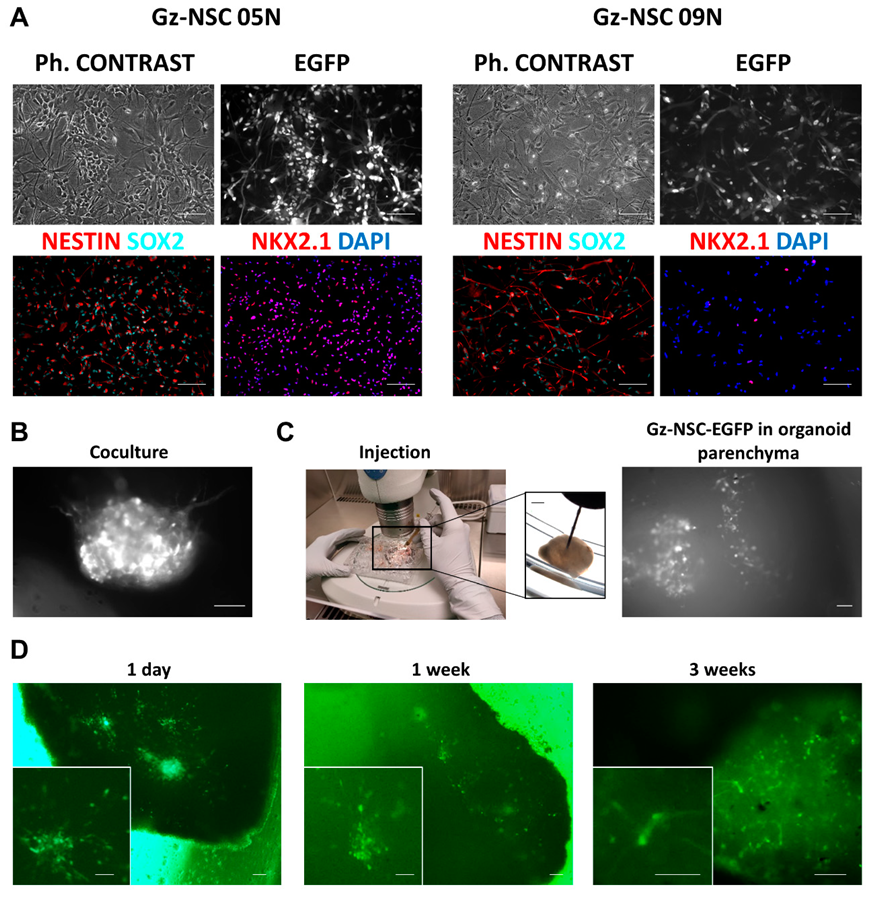
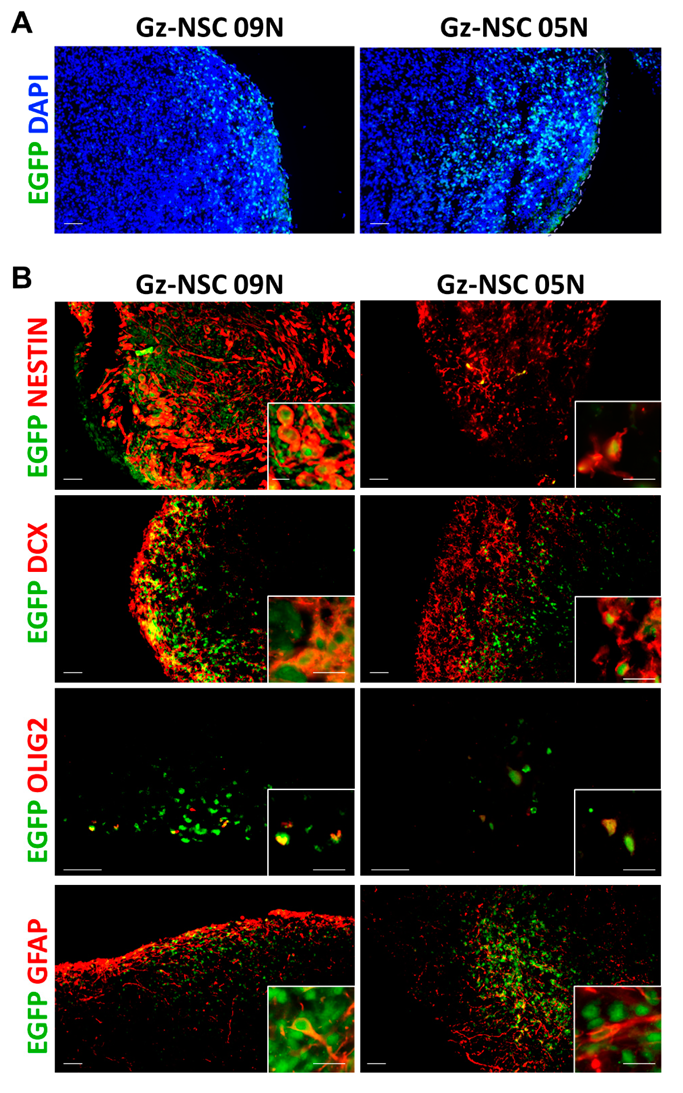
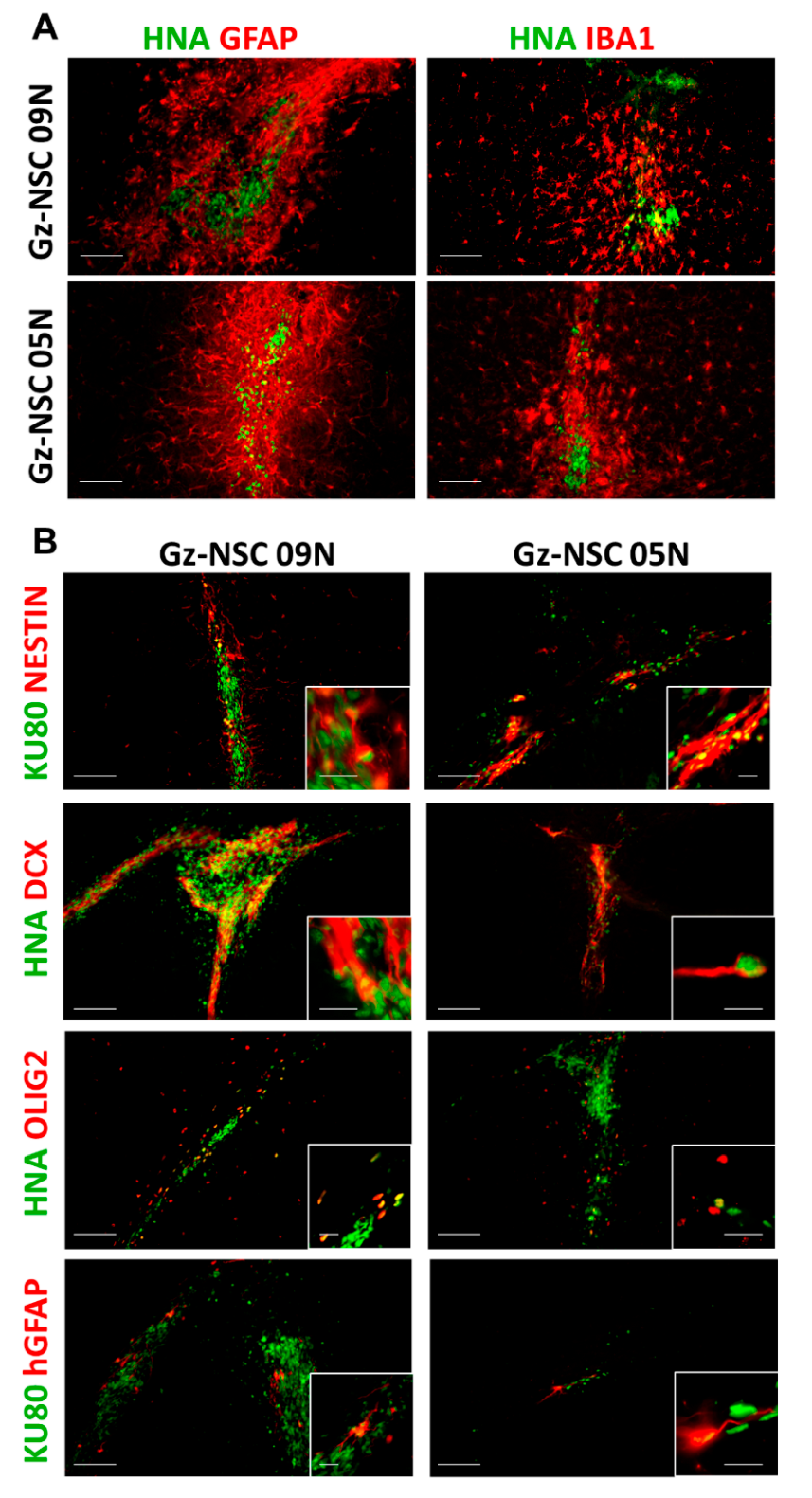
2.2. Transplantation into Brain Organoids
2.3. Transplantation into the Brain of Immunodeficient Animals
2.4. Immunofluorescence
2.5. Statistics
3. Results
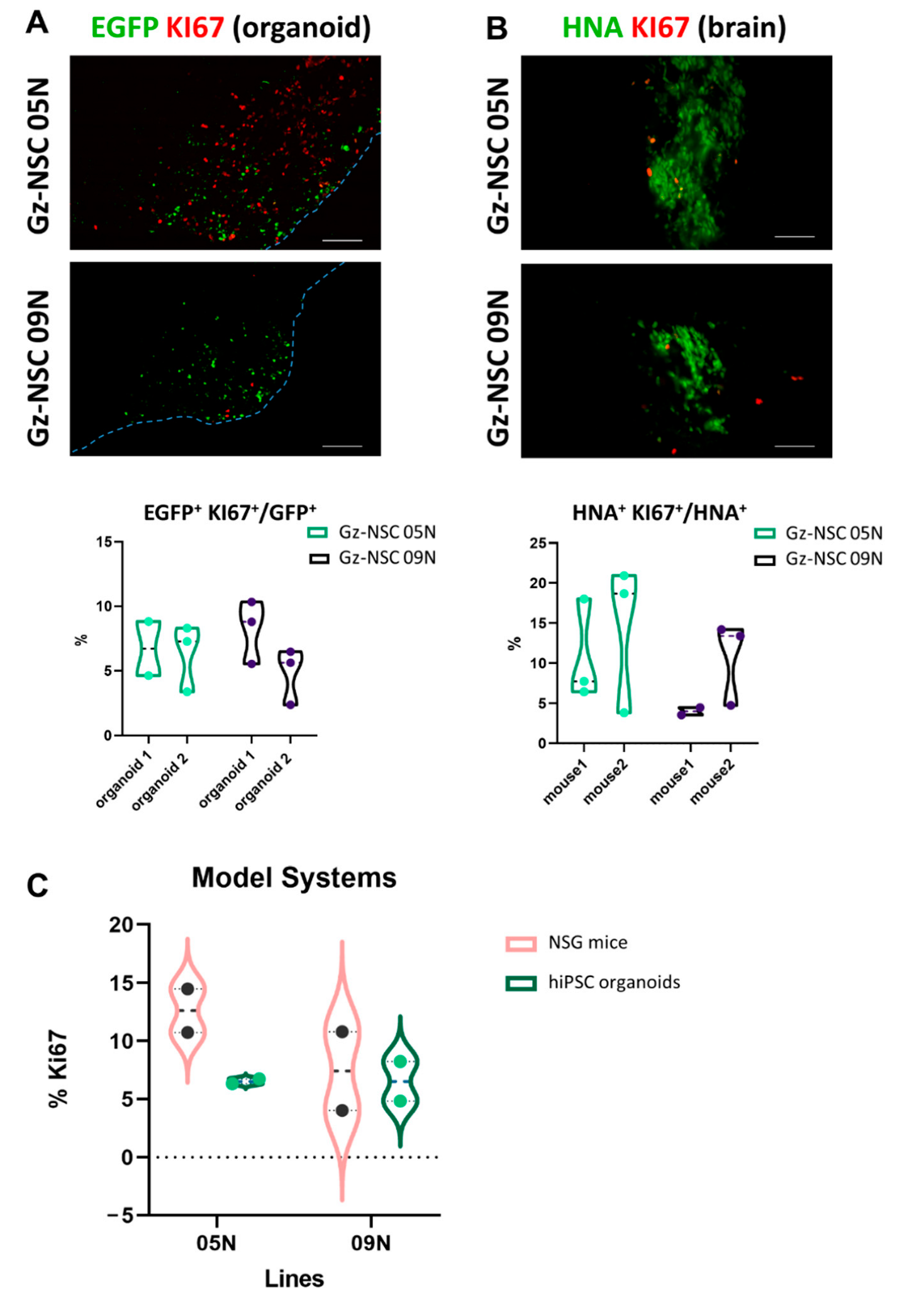
3.1. Transplantation into Human Brain Organoids
3.2. Transplantation into Immunodeficient Mouse Brains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bashor, C.J.; Hilton, I.B.; Bundukwala, H.; Smith, D.M.; Veiseh, O. Engineering the next generation of cell-based therapeutics. Nat. Rev. Drug Discov. 2022, 21, 655–675. [Google Scholar] [CrossRef]
- Mak, I.W.Y.; Evaniew, N.; Ghert, M. Lost in translation: Animal models and clinical trials in cancer treatment. Am. J. Transl. Res. 2014, 6, 114. [Google Scholar]
- Ransohoff, R.M. All (animal) models (of neurodegeneration) are wrong. Are they also useful? J. Exp. Med. 2018, 215, 2955–2958. [Google Scholar] [CrossRef] [Green Version]
- Donega, V.; Nijboer, C.H.; Van Velthoven, C.T.J.; Youssef, S.A.; De Bruin, A.; Van Bel, F.; Kavelaars, A.; Heijnen, C.J. Assessment of long-term safety and efficacy of intranasal mesenchymal stem cell treatment for neonatal brain injury in the mouse. Pediatr. Res. 2015, 78, 520–526. [Google Scholar] [CrossRef] [Green Version]
- Nabar, N.R.; Yuan, F.; Lin, X.; Wang, L.; Bai, G.; Mayl, J.; Li, Y.; Zhou, S.F.; Wang, J.; Cai, J.; et al. Cell Therapy: A Safe and Efficacious Therapeutic Treatment for Alzheimer’s Disease in APP+PS1 Mice. PLoS ONE 2012, 15, e0243343. [Google Scholar] [CrossRef]
- Rosati, J.; Ferrari, D.; Altieri, F.; Tardivo, S.; Ricciolini, C.; Fusilli, C.; Zalfa, C.; Profico, D.C.; Pinos, F.; Bernardini, L.; et al. Establishment of stable iPS-derived human neural stem cell lines suitable for cell therapies. Cell Death Dis. 2018, 9, 937. [Google Scholar] [CrossRef]
- Aguilera, Y.; Mellado-Damas, N.; Olmedo-Moreno, L.; López, V.; Panadero-Morón, C.; Benito, M.; Guerrero-Cázares, H.; Márquez-Vega, C.; Martín-Montalvo, A.; Capilla-González, V. Preclinical safety evaluation of intranasally delivered human mesenchymal stem cells in juvenile mice. Cancers 2021, 13, 1169. [Google Scholar] [CrossRef]
- Barrett, D.M.; Zhao, Y.; Liu, X.; Jiang, S.; Carpenito, C.; Kalos, M.; Carroll, R.G.; June, C.H.; Grupp, S.A. Treatment of advanced leukemia in mice with mRNA engineered T cells. Hum. Gene Ther. 2011, 22, 157–1586. [Google Scholar] [CrossRef] [Green Version]
- Van Norman, G.A. Limitations of Animal Studies for Predicting Toxicity in Clinical Trials: Is it Time to Rethink Our Current Approach? JACC Basic to Transl. Sci. 2019, 4, 845–854. [Google Scholar] [CrossRef]
- Dawson, T.M.; Golde, T.E.; Tourenne, C.L.; Programs, S.C.; Sciences, M.; Snyder, S.H.; Malvin, H.; Orleans, N.; Hospital, M.G. Animal Models of Neurodegenerative Diseases. Nat. Neurosci. 2018, 21, 1370–1379. [Google Scholar] [CrossRef] [PubMed]
- Azkona, G.; Sanchez-Pernaute, R. Mice in translational neuroscience: What R we doing? Prog. Neurobiol. 2022, 217, 102330. [Google Scholar] [CrossRef]
- Knight, E.; Przyborski, S. Advances in 3D cell culture technologies enabling tissue-like structures to be created in vitro. J. Anat. 2015, 14, 384–395. [Google Scholar] [CrossRef] [Green Version]
- Vergani, L.; Grattarola, M.; Nicolini, C. Modifications of chromatin structure and gene expression following induced alterations of cellular shape. Int. J. Biochem. Cell Biol. 2004, 36, 1447–1461. [Google Scholar] [CrossRef]
- Doss, M.; Sachinidis, A. Current Challenges of iPSC-Based Disease Modeling and Therapeutic Implications. Cells 2019, 8, 403. [Google Scholar] [CrossRef] [Green Version]
- de Leeuw, S.M.; Davaz, S.; Wanner, D.; Milleret, V.; Ehrbar, M.; Gietl, A.; Tackenberg, C. Increased maturation of iPSC-derived neurons in a hydrogel-based 3D culture. J. Neurosci. Methods 2021, 360, 109254. [Google Scholar] [CrossRef]
- Morata Tarifa, C.; López Navas, L.; Azkona, G.; Sánchez Pernaute, R. Chimeras for the twenty-first century. Crit. Rev. Biotechnol. 2020, 40, 283–291. [Google Scholar] [CrossRef]
- Bai, J.; Wang, C. Organoids and Microphysiological Systems: New Tools for Ophthalmic Drug Discovery. Front. Pharmacol. 2020, 11, 407. [Google Scholar] [CrossRef] [Green Version]
- Rabeling, A.; Goolam, M. Cerebral organoids as an in vitro model to study autism spectrum disorders. Gene Ther. 2022, 2022, 1–11. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Renner, M.; Martin, C.A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral organoids model human brain development and microcephaly. Nature 2013, 493, 385–388. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.; Lancaster, M.A.; Castanon, R.; Nery, J.R.; Knoblich, J.A.; Ecker, J.R. Cerebral Organoids Recapitulate Epigenomic Signatures of the Human Fetal Brain. Cell Rep. 2016, 17, 3369–3384. [Google Scholar] [CrossRef] [Green Version]
- Shou, Y.; Liang, F.; Xu, S.; Li, X. The Application of Brain Organoids: From Neuronal Development to Neurological Diseases. Front. Cell Dev. Biol. 2020, 8, 579659. [Google Scholar] [CrossRef]
- Trujillo, C.A.; Adams, J.W.; Negraes, P.D.; Carromeu, C.; Tejwani, L.; Acab, A.; Tsuda, B.; Thomas, C.A.; Sodhi, N.; Fichter, K.M.; et al. Pharmacological reversal of synaptic and network pathology in human MECP2-KO neurons and cortical organoids. EMBO Mol. Med. 2021, 13, e12523. [Google Scholar] [CrossRef]
- Samarasinghe, R.A.; Miranda, O.A.; Buth, J.E.; Mitchell, S.; Ferando, I.; Watanabe, M.; Allison, T.F.; Kurdian, A.; Fotion, N.N.; Gandal, M.J.; et al. Identification of neural oscillations and epileptiform changes in human brain organoids. Nat. Neurosci. 2021, 24, 1488–1500. [Google Scholar] [CrossRef]
- Fernandez-Muñoz, B.; Garcia-Delgado, A.B.; Arribas-Arribas, B.; Sanchez-Pernaute, R. Human neural stem cells for cell-based medicinal products. Cells 2021, 10, 2377. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Knoblich, J.A. Generation of cerebral organoids from human pluripotent stem cells. Nat. Protoc. 2014, 9, 2329–2340. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Muñoz, B.; Rosell-Valle, C.; Ferrari, D.; Alba-Amador, J.; Montiel, M.Á.; Campos-Cuerva, R.; Lopez-Navas, L.; Muñoz-Escalona, M.; Martín-López, M.; Profico, D.C.; et al. Retrieval of germinal zone neural stem cells from the cerebrospinal fluid of premature infants with intraventricular hemorrhage. Stem Cells Transl. Med. 2020, 9, 1085–1101. [Google Scholar] [CrossRef]
- Paxinos, G.F.K. The Mouse Brain in Stereotaxic Coordinates: Compact Second Edition; Acadademic Press: San Diego, CA, USA, 2004. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Ormel, P.R.; Vieira de Sá, R.; van Bodegraven, E.J.; Karst, H.; Harschnitz, O.; Sneeboer, M.A.M.; Johansen, L.E.; van Dijk, R.E.; Scheefhals, N.; Berdenis van Berlekom, A.; et al. Microglia innately develop within cerebral organoids. Nat. Commun. 2018, 9, 4167. [Google Scholar] [CrossRef] [Green Version]
- Cakir, B.; Tanaka, Y.; Kiral, F.R.; Xiang, Y.; Dagliyan, O.; Wang, J.; Lee, M.; Greaney, A.M.; Yang, W.S.; duBoulay, C.; et al. Expression of the transcription factor PU.1 induces the generation of microglia-like cells in human cortical organoids. Nat. Commun. 2022, 13, 430. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Corsini, N.S.; Wolfinger, S.; Gustafson, E.H.; Phillips, A.W.; Burkard, T.R.; Otani, T.; Livesey, F.J.; Knoblich, J.A. Guided self-organization and cortical plate formation in human brain organoids. Nat. Biotechnol. 2017, 35, 659–666. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Sullivan, G.J.; Park, I.H. How well do brain organoids capture your brain? iScience 2021, 24, 102063. [Google Scholar] [CrossRef] [PubMed]
- Emmett, S.R.; Dove, B.; Mahoney, L.; Wurm, T.; Hiscox, J.A. The cell cycle and virus infection. Methods Mol. Biol. 2005, 296, 197–218. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Delgado, A.B.; Campos-Cuerva, R.; Rosell-Valle, C.; Martin-López, M.; Casado, C.; Ferrari, D.; Márquez-Rivas, J.; Sánchez-Pernaute, R.; Fernández-Muñoz, B. Brain Organoids to Evaluate Cellular Therapies. Animals 2022, 12, 3150. https://doi.org/10.3390/ani12223150
García-Delgado AB, Campos-Cuerva R, Rosell-Valle C, Martin-López M, Casado C, Ferrari D, Márquez-Rivas J, Sánchez-Pernaute R, Fernández-Muñoz B. Brain Organoids to Evaluate Cellular Therapies. Animals. 2022; 12(22):3150. https://doi.org/10.3390/ani12223150
Chicago/Turabian StyleGarcía-Delgado, Ana Belén, Rafael Campos-Cuerva, Cristina Rosell-Valle, María Martin-López, Carlos Casado, Daniela Ferrari, Javier Márquez-Rivas, Rosario Sánchez-Pernaute, and Beatriz Fernández-Muñoz. 2022. "Brain Organoids to Evaluate Cellular Therapies" Animals 12, no. 22: 3150. https://doi.org/10.3390/ani12223150