
Characterizing the Gut Microbiota of Eurasian Otter (Lutra lutra chinensis) and Snub-Nosed Monkey (Rhinopithecus roxellana) to Enhance Conservation Practices in the Foping National Nature Reserve of China
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
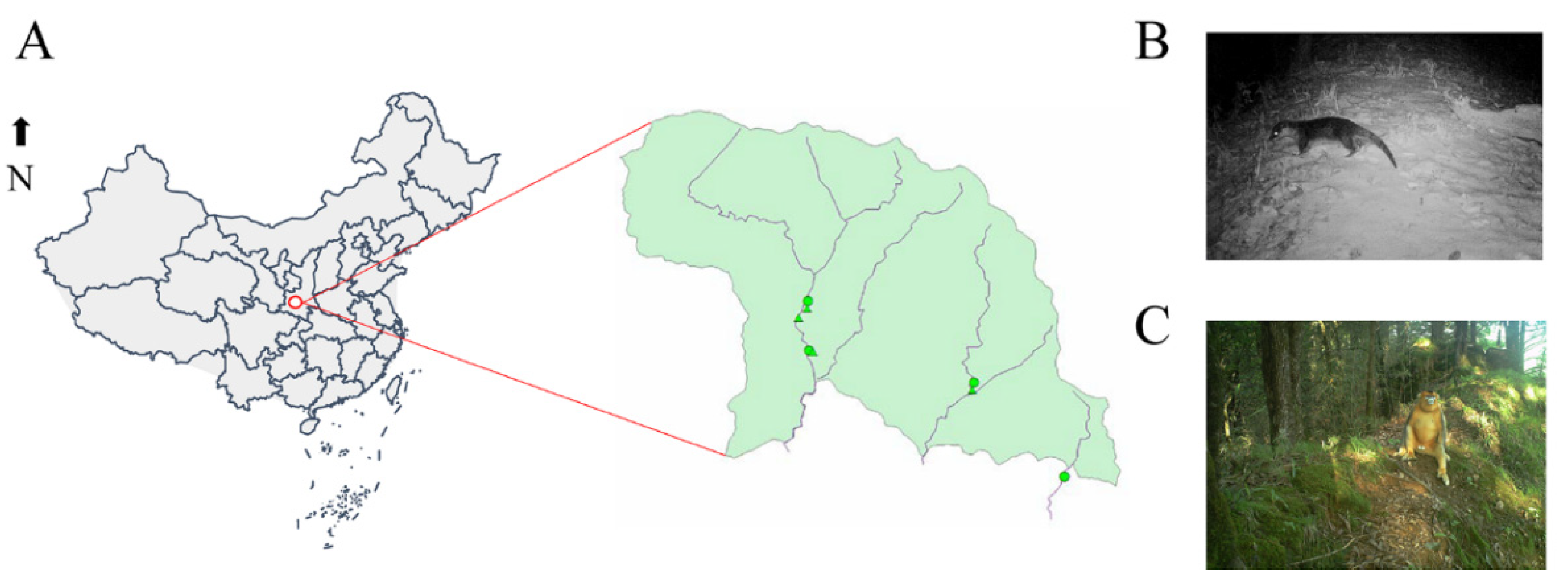
2.1. Overview of Collected Samples
2.2. Microbial Community Analysis
2.3. Short-Chain Fatty Acid Determination
2.4. Statistical Analysis
3. Results
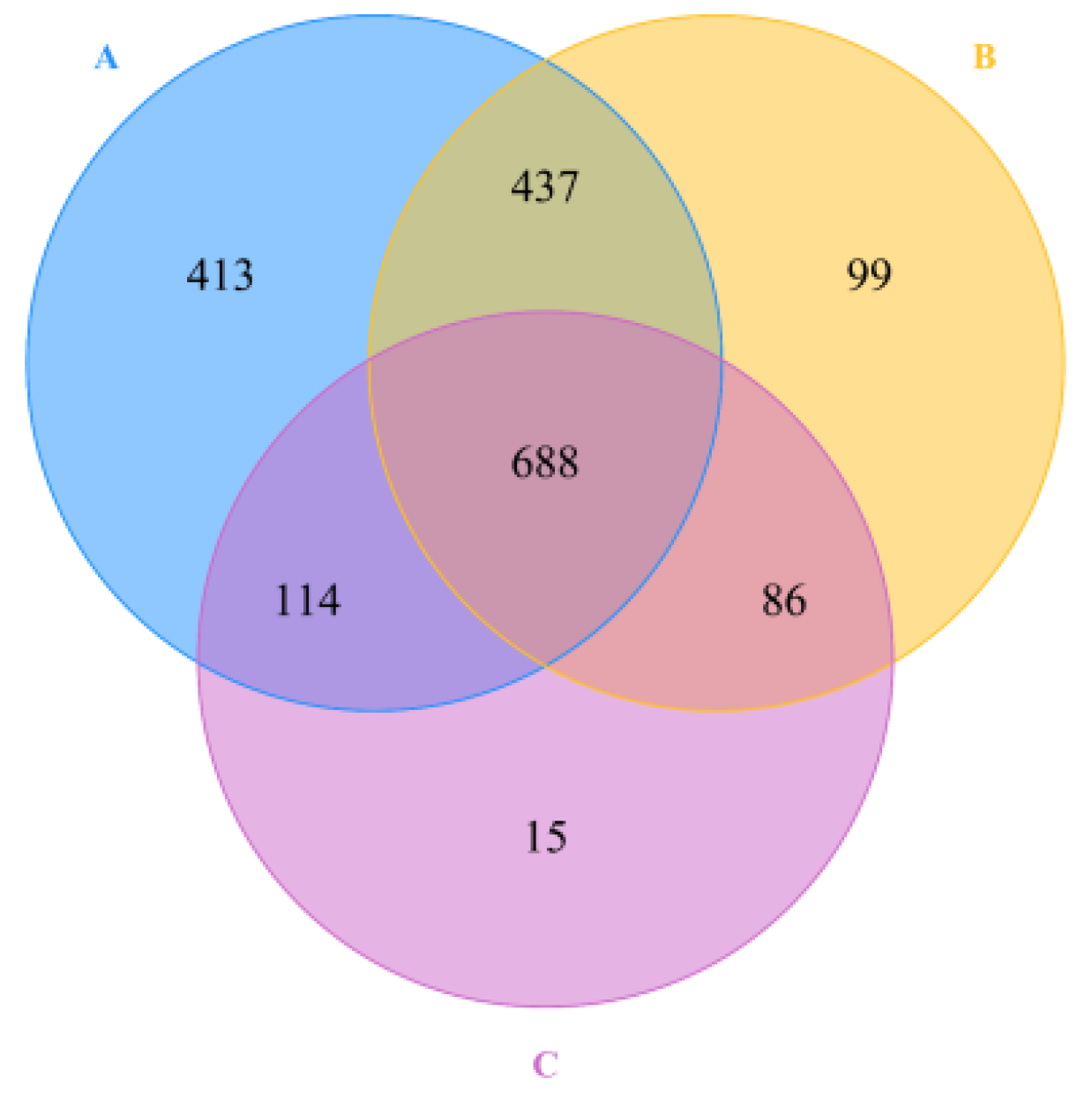
3.1. Sequencing Statistics
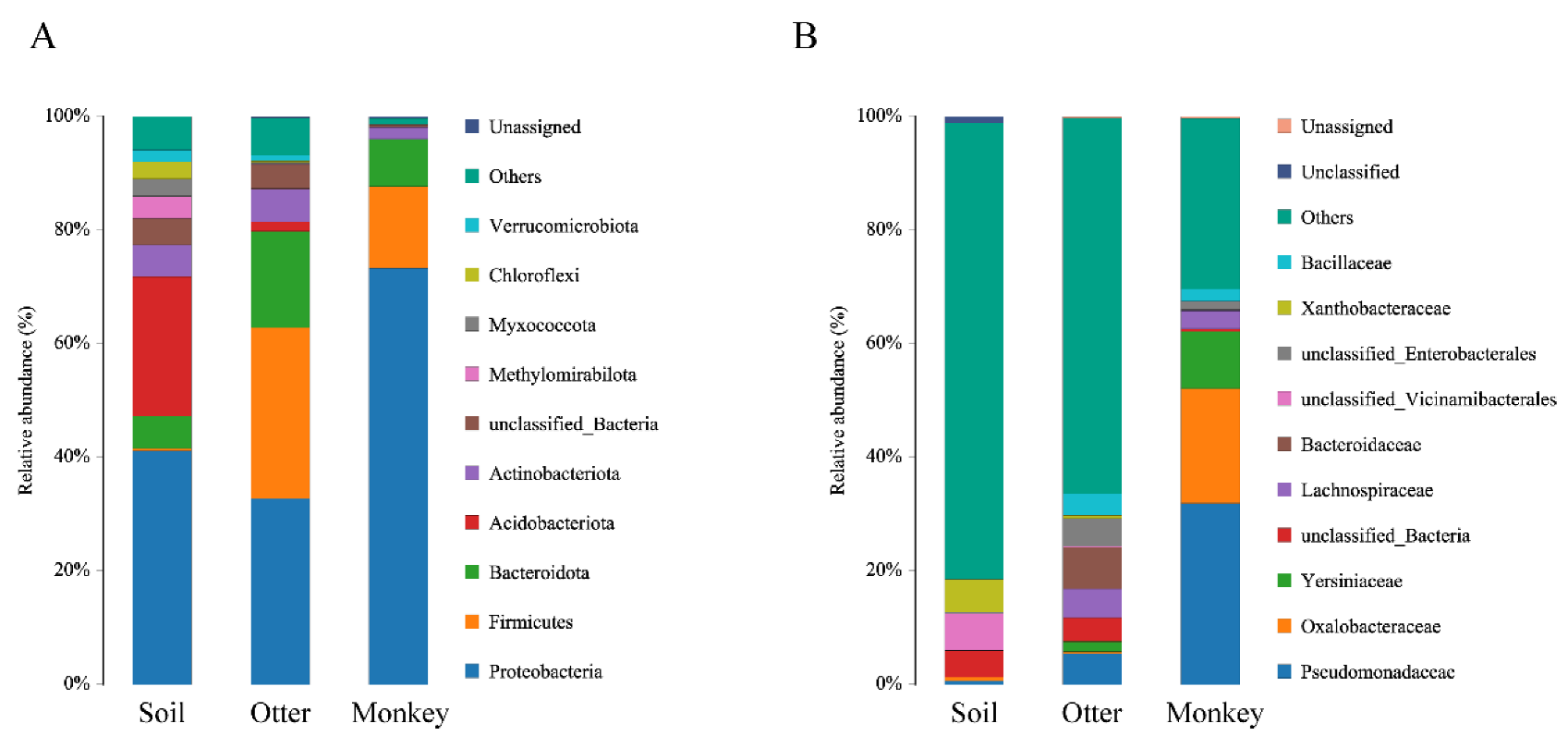
3.2. Composition of Fecal Microbial Communities
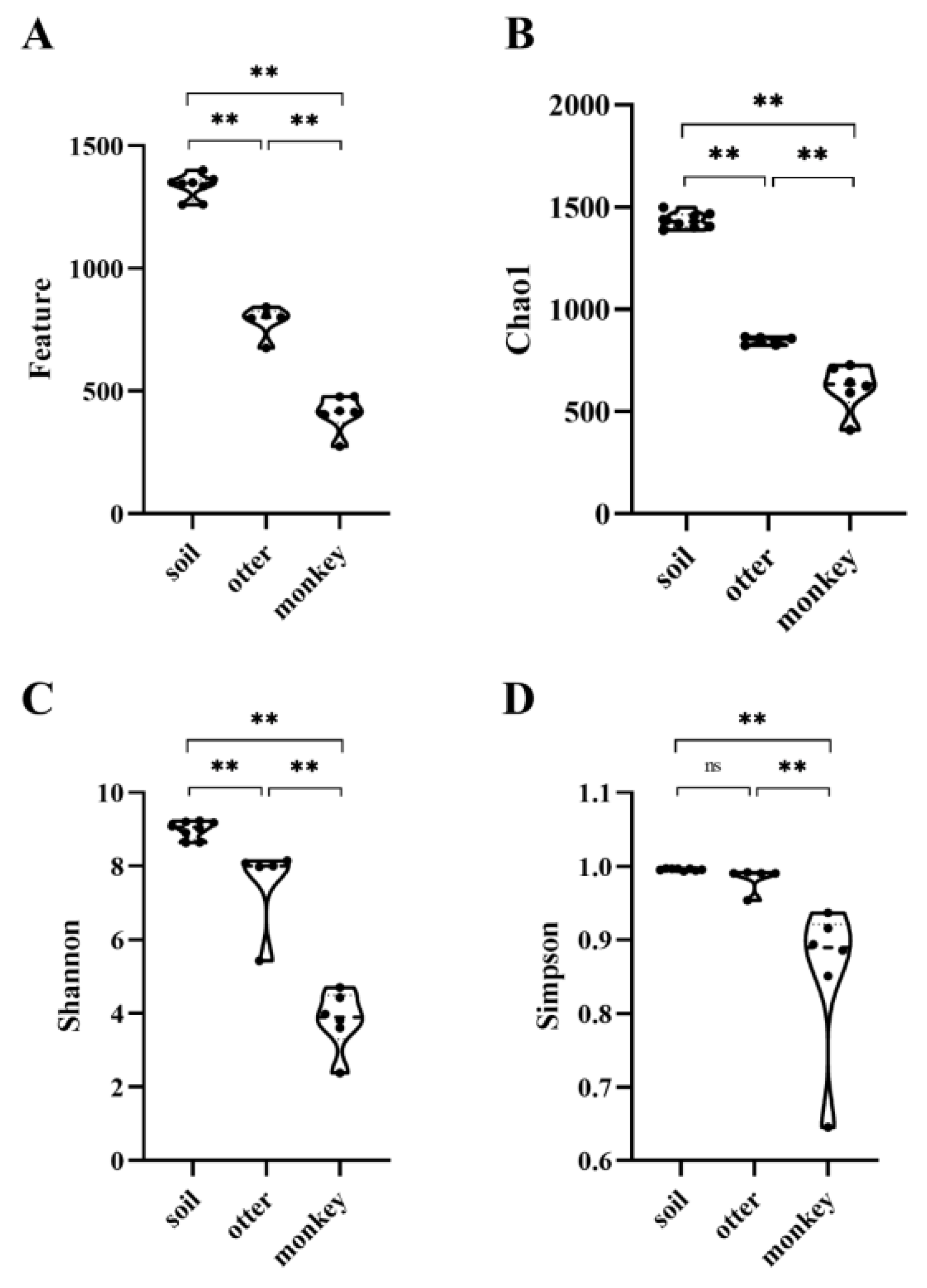
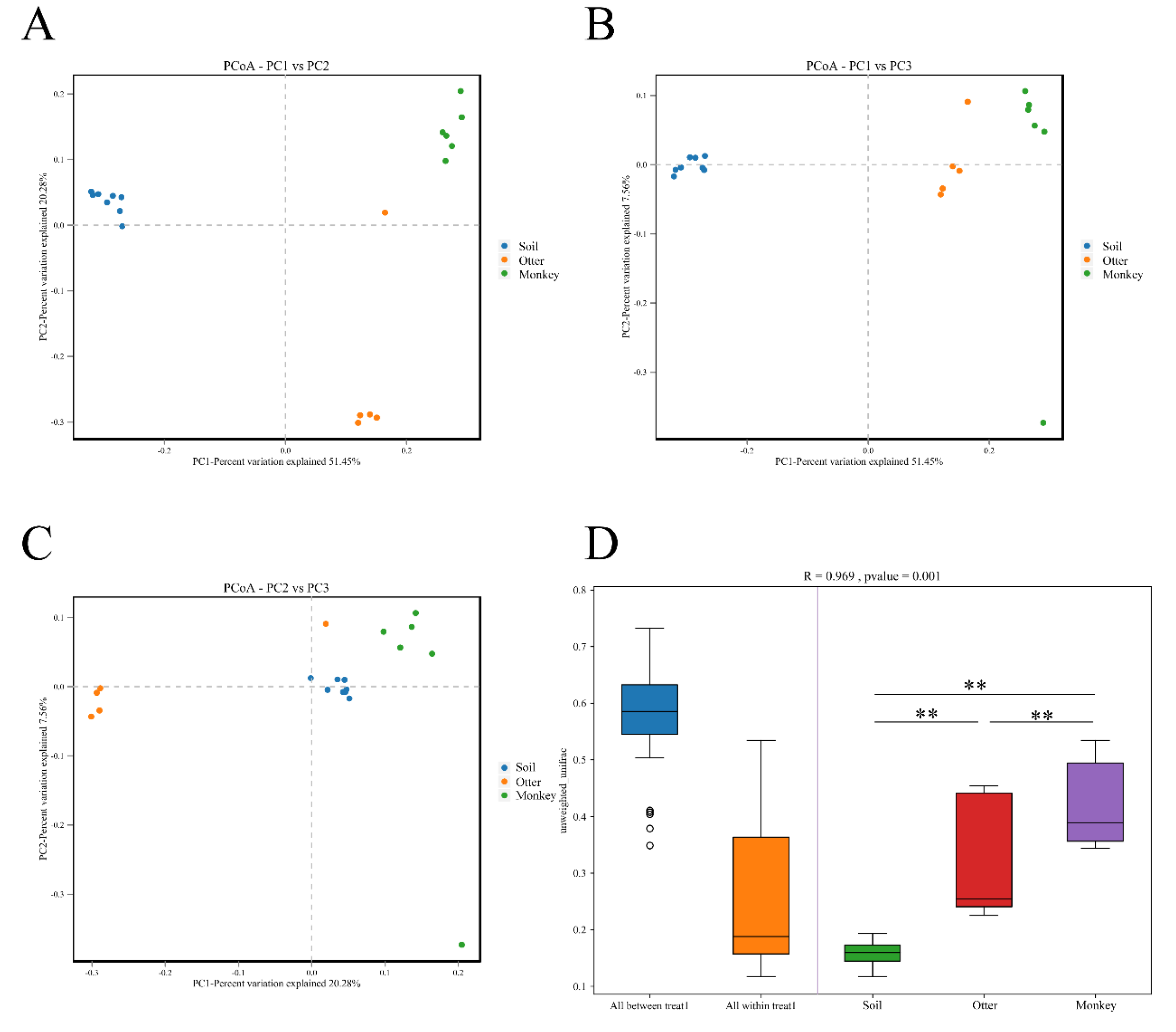
3.3. Diversity
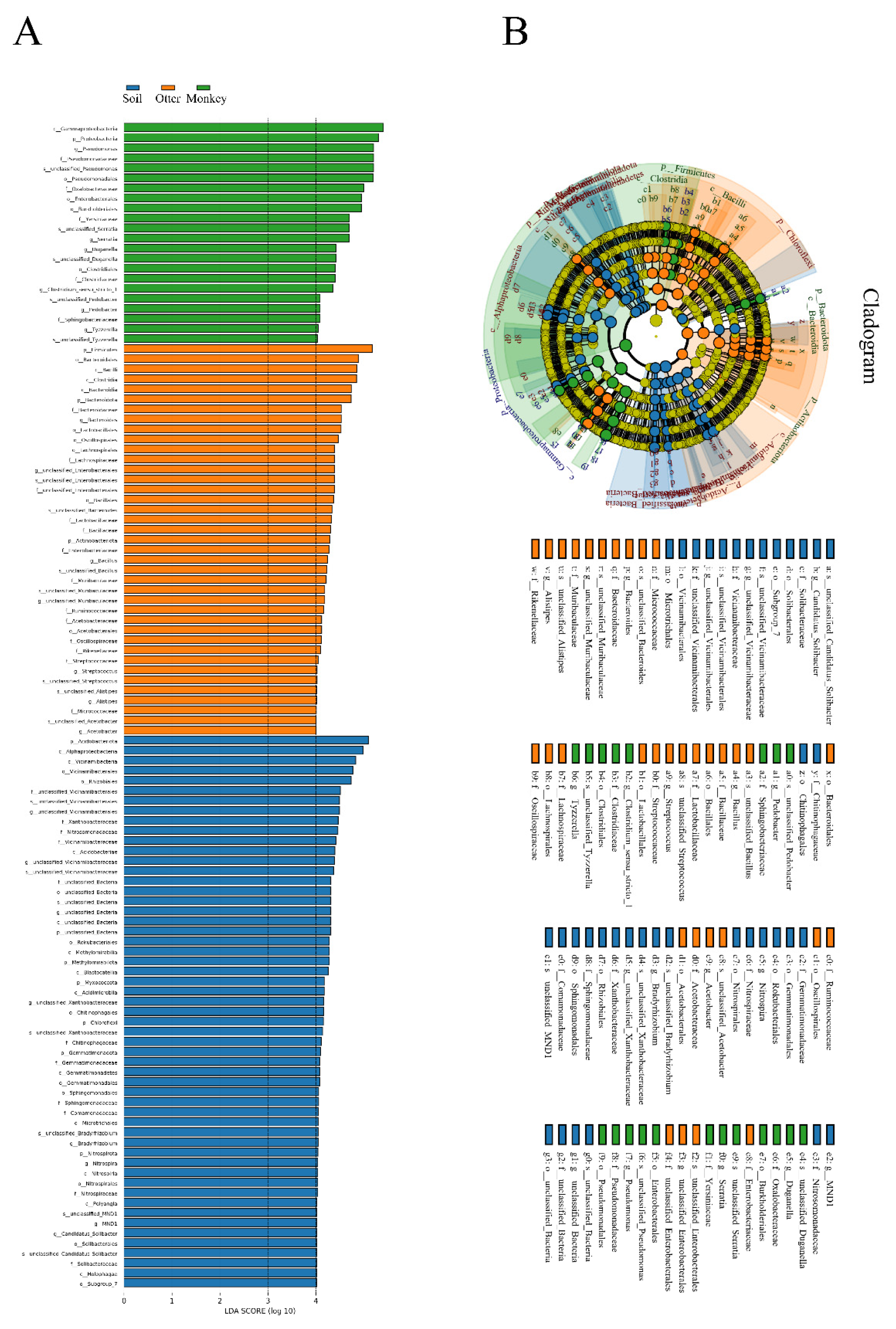
3.4. Difference Analysis
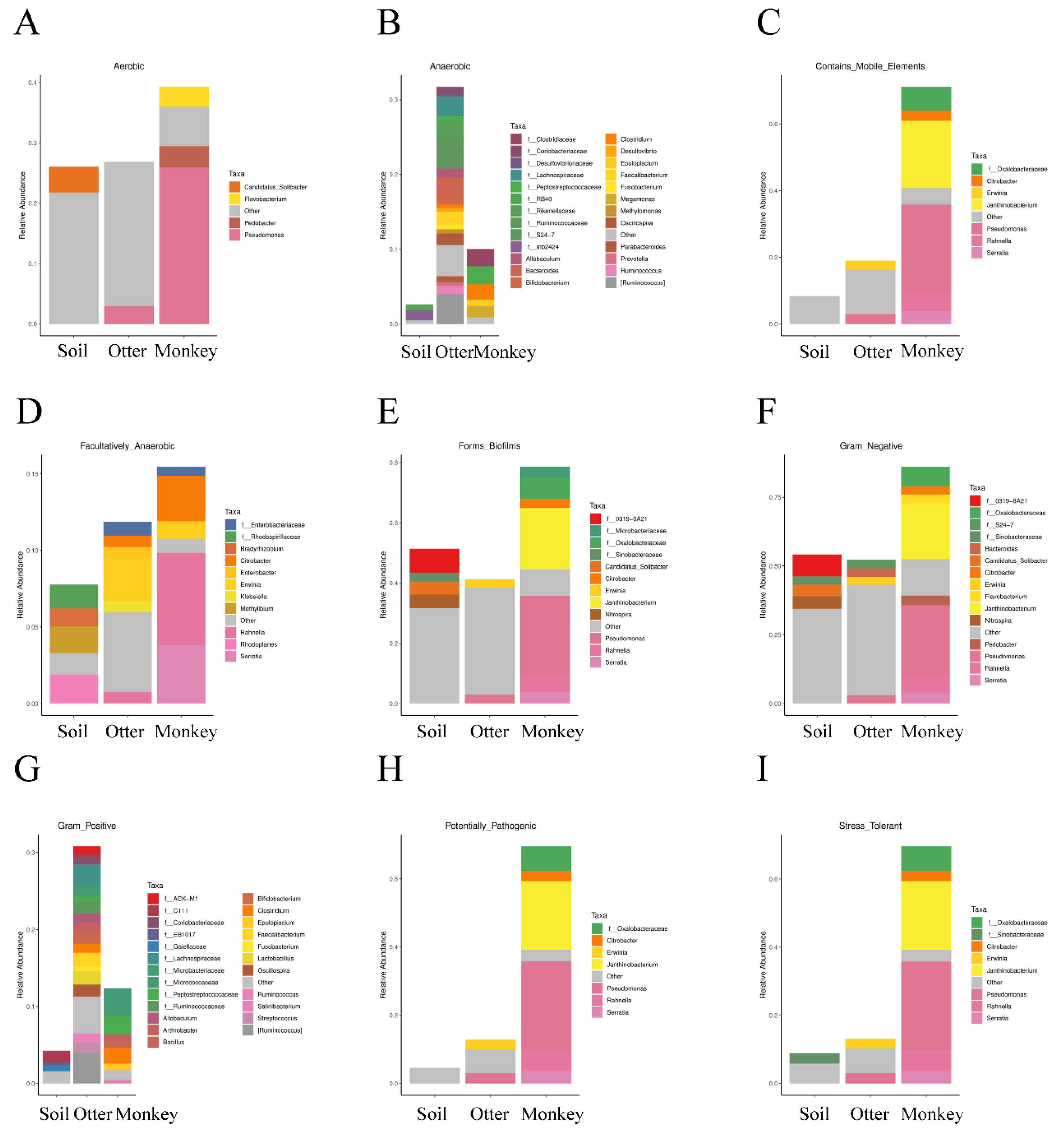
3.5. Prediction of Microbial Phenotype
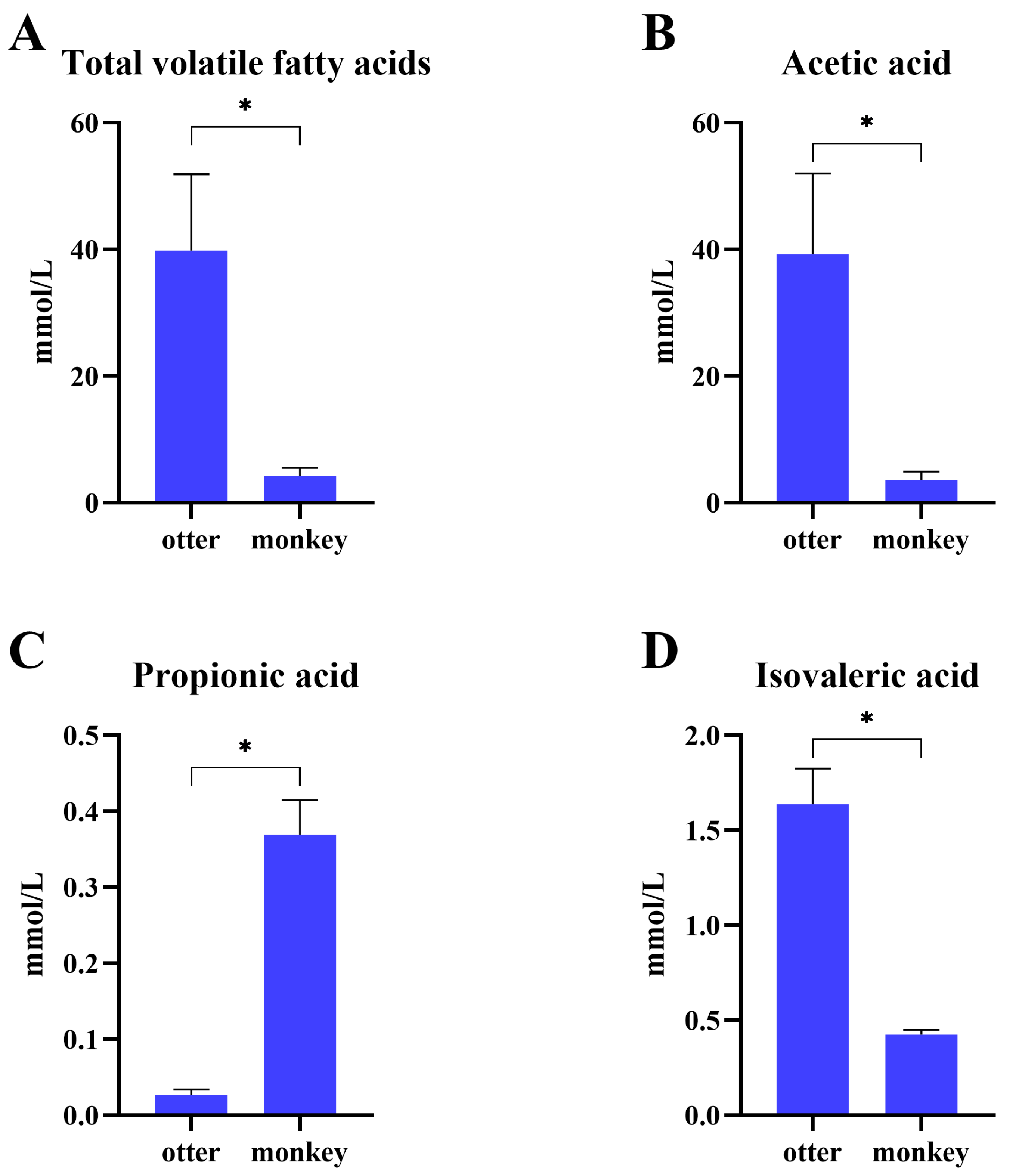
3.6. Concentration of Short-Chain Fatty Acids
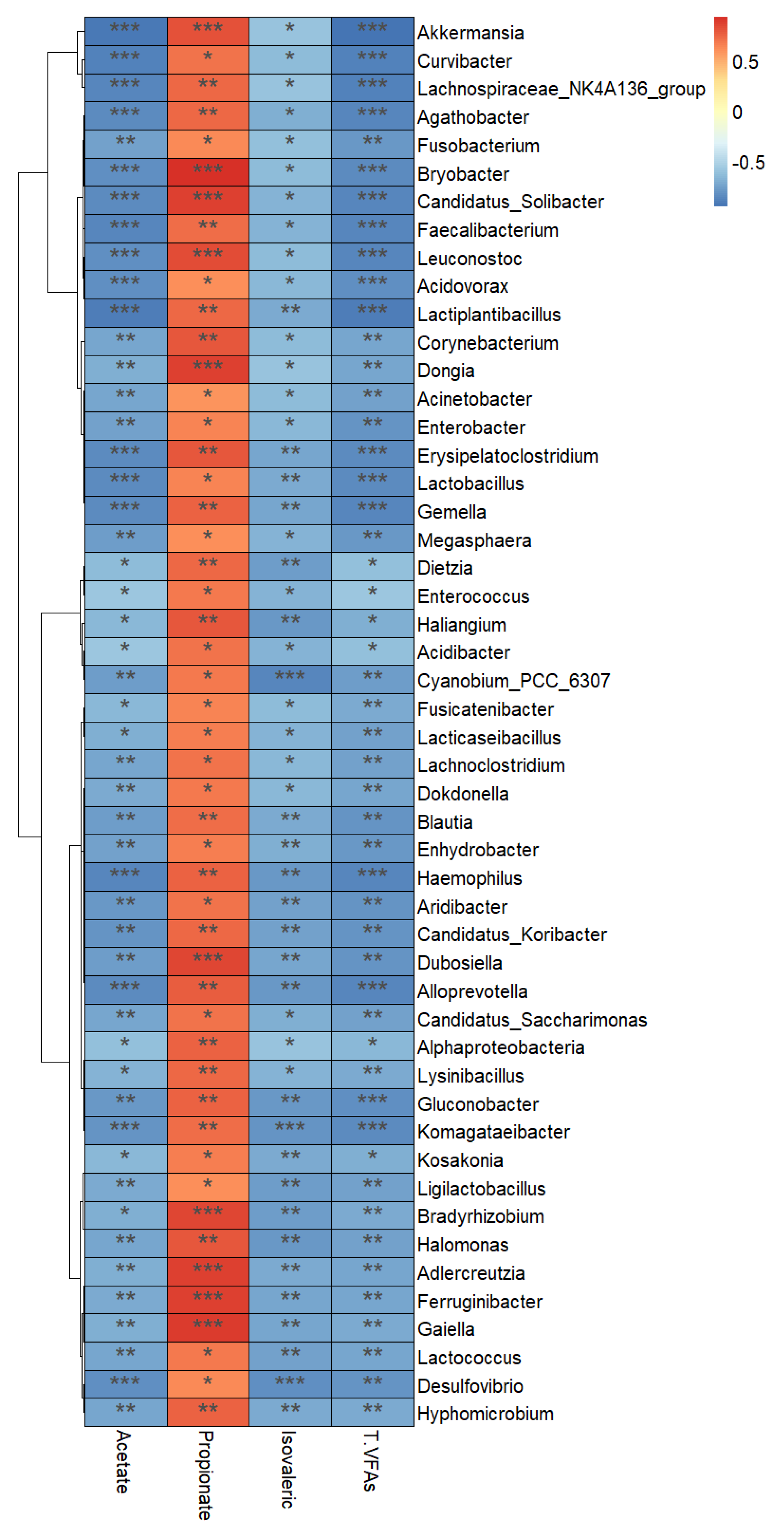
3.7. Correlations between the Fecal Microbiota and Short-Chain Fatty Acid Profile
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nishida, A.H.; Ochman, H. Rates of gut microbiome divergence in mammals. Mol. Ecol. 2018, 27, 1884–1897. [Google Scholar] [CrossRef]
- Dudek, N.K.; Switzer, A.D.; Costello, E.K.; Murray, M.J.; Tomoleoni, J.A.; Staedler, M.M.; Tinker, M.T.; Relman, D.A. Characterizing the oral and distal gut microbiota of the threatened southern sea otter (Enhydra lutris nereis) to enhance conservation practice. Conserv. Sci. Pract. 2022, 4, e12640. [Google Scholar] [CrossRef]
- Karmacharya, D.; Manandhar, P.; Manandhar, S.; Sherchan, A.M.; Sharma, A.N.; Joshi, J.; Bista, M.; Bajracharya, S.; Awasthi, N.P.; Sharma, N.; et al. Gut microbiota and their putative metabolic functions in fragmented Bengal tiger population of Nepal. PLoS ONE 2019, 14, e0221868. [Google Scholar] [CrossRef]
- Smith, C.C.; Snowberg, L.K.; Gregory Caporaso, J.; Knight, R.; Bolnick, D.I. Dietary input of microbes and host genetic variation shape among-population differences in stickleback gut microbiota. ISME J. 2015, 9, 2515–2526. [Google Scholar] [CrossRef]
- Frankel, J.S.; Mallott, E.K.; Hopper, L.M.; Ross, S.R.; Amato, K.R. The effect of captivity on the primate gut microbiome varies with host dietary niche. Am. J. Primatol. 2019, 81, e23061. [Google Scholar] [CrossRef]
- Bornbusch, S.L.; Greene, L.K.; Rahobilalaina, S.; Calkins, S.; Rothman, R.S.; Clarke, T.A.; Lafleur, M.; Drea, C.M. Gut microbiota of ring-tailed lemurs (Lemur catta) vary across natural and captive populations and correlate with environmental microbiota. Anim. Microbiome 2022, 4, 29. [Google Scholar] [CrossRef]
- Kim, I.S.; Sim, J.H.; Cho, J.W.; Kim, B.; Lee, Y.; Ahn, D. Osteoporosis in an Asian small-clawed otter (Aonyx cinereus Illiger, 1815). J. Vet. Med. Sci. 2020, 82, 376–378. [Google Scholar] [CrossRef] [Green Version]
- Day, C.C.; Westover, M.; Mcmillan, B.R. Seasonal diet of the northern river otter (Lontra canadensis): What drives prey selection? Can. J. Zool. 2015, 93, 197–205. [Google Scholar] [CrossRef]
- Qi, X.G.; Garber, P.A.; Ji, W.; Huang, Z.P.; Huang, K.; Zhang, P.; Guo, S.T.; Wang, X.W.; He, G.; Zhang, P.; et al. Satellite telemetry and social modeling offer new insights into the origin of primate multilevel societies. Nat. Commun. 2014, 5, 5296. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.P.; Bian, K.; Liu, Y.; Pan, R.L.; Qi, X.G.; Li, B.G. Male dispersal pattern in golden snub-nosed monkey (Rhinopithecus roxellana) in Qinling mountains and its conservation implication. Sci. Rep. 2017, 7, 46217. [Google Scholar] [CrossRef]
- Mcfall-Ngai, M.; Hadfield, M.G.; Bosch, T.C.; Carey, H.V.; Domazet-Lošo, T.; Douglas, A.E.; Dubilier, N.; Eberl, G.; Fukami, T.; Gilbert, S.F.; et al. Animals in a bacterial world, a new imperative for the life sciences. Proc. Natl. Acad. Sci. USA 2013, 110, 3229–3236. [Google Scholar] [CrossRef] [Green Version]
- Peng, Z.; Zhang, C.; Shen, M.; Bao, H.; Hou, Z.; He, S.; Hua, Y. Baylisascaris schroederi infection in giant pandas (Ailuropoda melanoleuca) in Foping National Nature Reserve, China. J. Wildl. Dis. 2017, 53, 854–858. [Google Scholar] [CrossRef]
- Yu, Z.; Morrison, M. Comparisons of different hypervariable regions of rrs genes for use in fingerprinting of microbial community by PCR-denaturing gradient gel electrophoresis. Appl. Environ. Microbiol. 2004, 70, 4800–4806. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wang, Z.; Dong, C.; Li, F.; Wang, W.; Yuan, Z.; Mo, F.; Weng, X. Rumen bacteria communities and performances of fattening lambs with a lower or greater subacute ruminal acidoss risk. Front. Microbiol. 2017, 8, 2506. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, A.; Harty, S.; Johnson, K.V.; Moeller, A.H.; Archie, E.A.; Schell, L.D.; Carmody, R.N.; Clutton-Brock, T.H.; Dunbar, R.I.M.; Burnet, P.W.J. Microbial transmission in animal social networks and the social microbiome. Nat. Ecol. Evol. 2020, 4, 1020–1035. [Google Scholar] [CrossRef]
- Sharma, A.K.; Petrzelkova, K.; Pafco, B.; Jost Robinson, C.A.; Fuh, T.; Wilson, B.A.; Stumpf, R.M.; Torralba, M.G.; Blekhman, R.; White, B.; et al. Traditional human populations and nonhuman primates show parallel gut microbiome adaptations to analogous ecological conditions. mSystems 2020, 5, e00815-20. [Google Scholar] [CrossRef]
- Clayton, J.B.; Gomez, A.; Amato, K.; Knights, D.; Travis, D.A.; Blekhman, R.; Knight, R.; Leigh, S.; Stumpf, R.; Wolf, T. The gut microbiome of nonhuman primates: Lessons in ecology and evolution. Am. J. Primatol. 2020, 80, e22867. [Google Scholar] [CrossRef]
- Moeller, A.H.; Li, Y.; Mpoudi Ngole, E.; Ahuka-Mundeke, S.; Lonsdorf, E.V.; Pusey, A.E.; Peeters, M.; Hahn, B.H.; Ochman, H. Rapid changes in the gut microbiome during human evolution. Proc. Natl. Acad. Sci. USA 2014, 111, 16431–16435. [Google Scholar] [CrossRef] [Green Version]
- Xia, W.; Liu, G.; Wang, D.; Chen, H.; Zhu, L.; Li, D. Functional convergence of Yunnan snub-nosed monkey and bamboo-eating panda gut microbiomes revealing the driving by dietary flexibility on mammal gut microbiome. Comput. Struct. Biotechnol. J. 2022, 20, 685–699. [Google Scholar] [CrossRef]
- Guo, G.; Eccles, K.M.; Mcmillan, M.; Thomas, P.J.; Chan, H.M.; Poulain, A.J. The gut microbial community structure of the north American river otter (Lontra canadensis) in the Alberta oil sands region in Canada: Relationship with local environmental variables and metal body burden. Environ. Toxicol. Chem. 2020, 39, 2516–2526. [Google Scholar] [CrossRef]
- Okamoto, Y.; Ichinohe, N.; Woo, C.; Han, S.Y.; Kim, H.H.; Ito, S.; Nakamura, C.; Kumura, J.; Nagaoka, K.; Yamamoto, N. Contrasting gut microbiota in captive Eurasian otters (Lutra lutra) by age. Arch. Microbiol. 2021, 203, 5405–5416. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, M.; Zhu, J.; Gao, Y.; Sha, W.; Ding, H.; Jiang, W.; Wu, S. Age, gender, and feeding environment influence fecal microbial diversity in spotted hyenas (Crocuta crocuta). Curr. Microbiol. 2020, 77, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Liu, D.; Zhai, J.; Zhang, L.; Ma, Y.; Xu, Y.; Rong, K.; Ma, J. Metagenomic analysis revealed the effects of goat milk feeding and breast feeding on the gut microbiome of Amur tiger cubs. Biochem. Biophys. Res. Commun. 2018, 503, 2590–2596. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.; Sales-Luís, T.; Duarte, A.; Nunes, S.F.; Carneiro, C.; Tenreiro, T.; Tenreiro, R.; Santos-Reis, M.; Tavares, L.; Vilela, C.L. First assessment of microbial diversity in faecal microflora of Eurasian otter (Lutra lutra Linnaeus, 1758) in Portugal. Eur. J. Wildl. Res. 2008, 54, 245–252. [Google Scholar] [CrossRef]
- Amato, K.R.; Yeoman, C.J.; Cerda, G.; Schmitt, C.A.; Cramer, J.D.; Miller, M.E.; Gomez, A.; Turner, T.R.; Wilson, B.A.; Stumpf, R.M.; et al. Variable responses of human and non-human primate gut microbiomes to a Western diet. Microbiome 2015, 3, 53. [Google Scholar] [CrossRef] [Green Version]
- Sang, A.G.P.; Guharat, S.; Wananukul, W. A mass cyanide poisoning from pickling bamboo shoots. Clin. Toxicol. 2011, 49, 834–839. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, T.; Lu, G.; Zhu, L. Lessons from bamboo-eating pandas and their gut microbiome: Gut microbiome flow and applications. Evol. Appl. 2020, 13, 615–619. [Google Scholar] [CrossRef] [Green Version]
- Selway, C.A.; Mills, J.G.; Weinstein, P.; Skelly, C.; Yadav, S.; Lowe, A.; Breed, M.F.; Weyrich, L.S. Transfer of environmental microbes to the skin and respiratory tract of humans after urban green space exposure. Environ. Int. 2020, 145, 106084. [Google Scholar] [CrossRef]
- Blum, W.E.H.; Zechmeister-Boltenstern, S.; Keiblinger, K.M. Does soil contribute to the human gut microbiome? Microorganisms 2019, 7, 287. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Zhang, H.; Bai, Z.; Zhang, A.; Bai, F.; Luo, X.; Hou, Y.; Ding, X.; Sun, B.; Sun, X.; et al. Exposure to soil, house dust and decaying plants increases gut microbial diversity and decreases serum immunoglobulin E levels in BALB/c mice. Environ. Microbiol. 2016, 18, 1326–1337. [Google Scholar] [CrossRef]
- Senghor, B.; Sokhna, C.; Ruimy, R.; Lagier, J.C. Gut microbiota diversity according to dietary habits and geographical provenance. Hum. Microb. J. 2018, 7–8, 1–9. [Google Scholar] [CrossRef]
- Grieneisen, L.E.; Charpentier, M.J.E.; Alberts, S.C.; Blekhman, R.; Bradburd, G.; Tung, J.; Archie, E.A. Genes, geology and germs: Gut microbiota across a primate hybrid zone are explained by site soil properties, not host species. Proc. Biol. Sci. 2019, 286, 20190431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loo, Y.T.; Howell, K.; Chan, M.; Zhang, P.; Ng, K. Modulation of the human gut microbiota by phenolics and phenolic fiber-rich foods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1268–1298. [Google Scholar] [CrossRef] [PubMed]
- Puddu, A.; Sanguineti, R.; Montecucco, F.; Viviani, G.L. Evidence for the gut microbiota short-chain fatty acids as key pathophysiological molecules improving diabetes. Mediat. Inflamm. 2014, 2014, 162021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowd, S.E.; Callaway, T.R.; Wolcott, R.D.; Sun, Y.; Mckeehan, T.; Hagevoort, R.G.; Edrington, T.S. Evaluation of the bacterial diversity in the feces of cattle using 16S rDNA bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP). BMC Microbiol. 2008, 8, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuhashi, T.; Sugitate, K.; Nakai, T.; Jikumaru, Y.; Ishihara, G. Rapid profiling method for mammalian feces short chain fatty acids by GC-MS. Anal. Biochem. 2018, 543, 51–54. [Google Scholar] [CrossRef]
- Guo, W.; Chen, Y.; Wang, C.; Ning, R.; Zeng, B.; Tang, J.; Li, C.; Zhang, M.; Li, Y.; Ni, Q.; et al. The carnivorous digestive system and bamboo diet of giant pandas may shape their low gut bacterial diversity. Conserv. Physiol. 2020, 8, coz104. [Google Scholar] [CrossRef]
- Compo, N.R.; Gomez, D.E.; Tapscott, B.; Weese, J.S.; Turner, P.V. Fecal bacterial microbiota of Canadian commercial mink (Neovison vison): Yearly, life stage, and seasonal comparisons. PLoS ONE 2018, 13, e0207111. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yang, Y.; Ishii, M.; Nagata, M.; Aw, W.; Obana, N.; Tomita, M.; Nomura, N.; Fukuda, S. Does the gut microbiota modulate host physiology through polymicrobial biofilms? Microbes Environ. 2020, 35, ME20037. [Google Scholar] [CrossRef]
- Brevik, E.C.; Pereg, L. History of soils in relation to animal and human health. In The Nexus of Soils, Plants, Animals and Human Hearth; Catena Soil Sciences: Stuttgart, Germeny, 2017. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, D.; Xie, T.; Du, R.; Guo, L. Characterizing the Gut Microbiota of Eurasian Otter (Lutra lutra chinensis) and Snub-Nosed Monkey (Rhinopithecus roxellana) to Enhance Conservation Practices in the Foping National Nature Reserve of China. Animals 2022, 12, 3097. https://doi.org/10.3390/ani12223097
Zhu D, Xie T, Du R, Guo L. Characterizing the Gut Microbiota of Eurasian Otter (Lutra lutra chinensis) and Snub-Nosed Monkey (Rhinopithecus roxellana) to Enhance Conservation Practices in the Foping National Nature Reserve of China. Animals. 2022; 12(22):3097. https://doi.org/10.3390/ani12223097
Chicago/Turabian StyleZhu, Dapeng, Tongtong Xie, Ruifang Du, and Long Guo. 2022. "Characterizing the Gut Microbiota of Eurasian Otter (Lutra lutra chinensis) and Snub-Nosed Monkey (Rhinopithecus roxellana) to Enhance Conservation Practices in the Foping National Nature Reserve of China" Animals 12, no. 22: 3097. https://doi.org/10.3390/ani12223097