
The Upregulation of Cathepsin G Is Associated with Resistance to Bovine Paratuberculosis

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. MAP Strain, Bacterial Culture, and Preparation of Bacterial Suspensions
2.3. Peripheral Blood Mononuclear Cells’ (PBMCs) Purification, CD14+ Monocyte Selection, Ex Vivo Differentiation to Monocyte-Derived Macrophages (MDMs), and MAP Infection
2.4. Mycobacterial Growth-Inhibition Assay (MGIA) Assessment Using the BACTEC MGIT 960 System
2.5. Bovine CTSG ELISA
2.6. Genotyping, Case–Control Population, and Odds Ratio (OR)
3. Results
3.1. The Presence of the Minor Allele in the rs41976219 (AC) Resulted in Increased CTSG Protein Levels in Supernatants of MAP-Infected CD14+-MDMs after 2 h of Infection
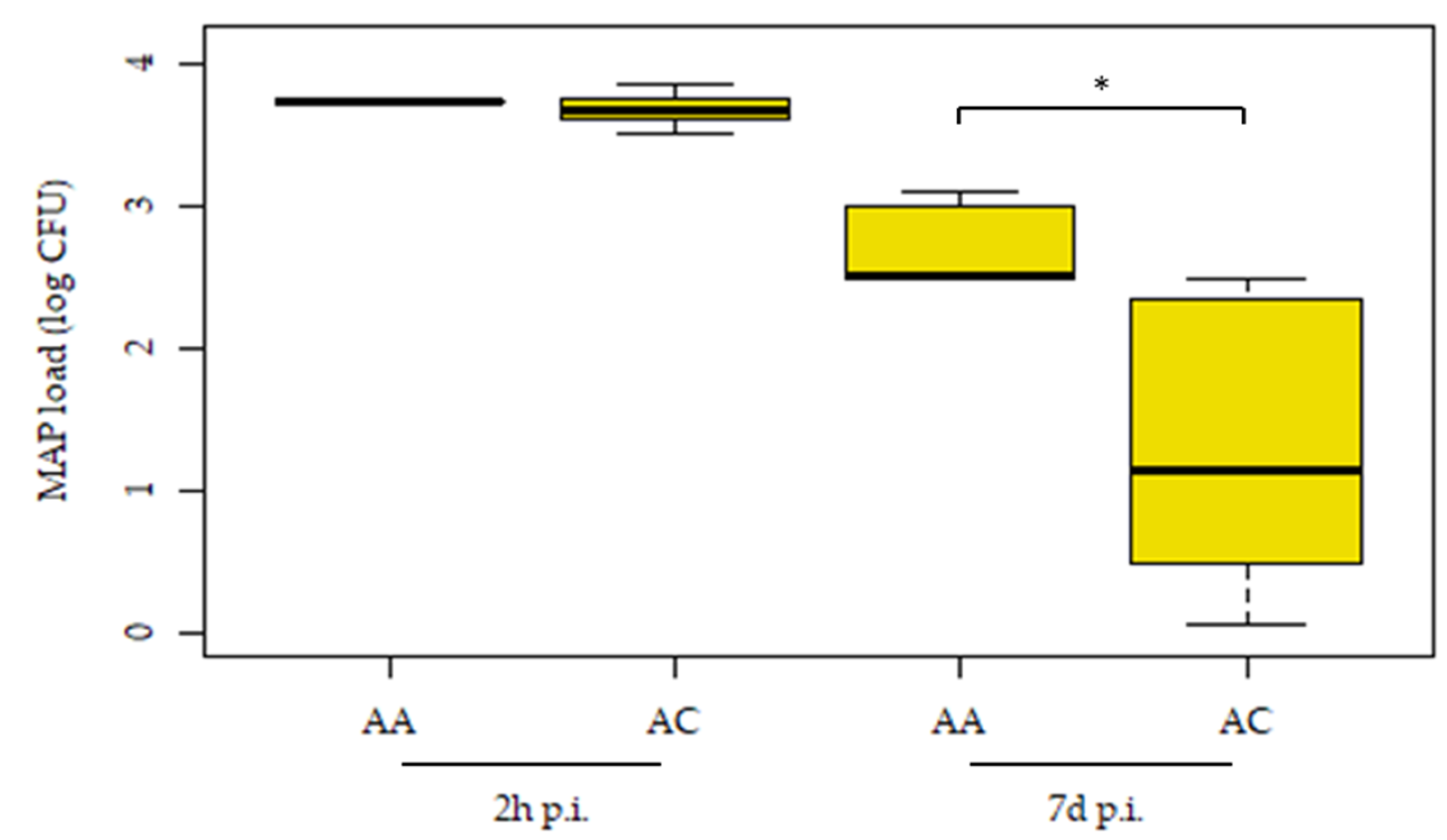
3.2. Association between the Inhibition of MAP Growth within MDMs and the cis-eQTL-rs41976219 AC Genotype Using an MGIA Assay
3.3. The cis-eQTL-rs41976219 T/T Genotype Correlated with PTB Control
3.4. The Presence of Two Minor Alleles in the cis-eQTL-rs41976219 (CC) Increased CTSG Protein Levels in the Plasmas of Healthy Cows
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rasmussen, P.; Barkema, H.W.; Mason, S.; Beaulieu, E.; Hall, D.C. Economic losses due to Johne’s disease (paratuberculosis) in dairy cattle. J. Dairy Sci. 2021, 104, 3123–3143. [Google Scholar] [CrossRef]
- González, J.; Geijo, M.V.; García-Pariente, C.; Verna, A.; Corpa, J.M.; Reyes, L.E.; Ferreras, M.C.; Juste, R.A.; García Marín, J.F.; Pérez, V. Histopathological classification of lesions associated with natural paratuberculosis infection in cattle. J. Comp. Pathol. 2005, 133, 184–196. [Google Scholar] [CrossRef]
- Balseiro, A.; Perez, V.; Juste, R.A. Chronic regional intestinal inflammatory disease: A trans-species slow infection? Comp. Immunol. Microbiol. Infect. Dis. 2019, 62, 88–100. [Google Scholar] [CrossRef]
- Juste, R.A.; Elguezabal, N.; Pavón, A.; Garrido, J.M.; Geijo, M.; Sevilla, I.; Cabriada, J.L.; Tejada, A.; García-Campos, F.; Casado, R.; et al. Association between Mycobacterium avium subsp. paratuberculosis DNA in blood and cellular and humoral immune response in inflammatory bowel disease patients and controls. Int. J. Infect. Dis. 2009, 13, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Pierce, E.S. Could Mycobacterium avium subspecies paratuberculosis cause Crohn’s disease, ulcerative colitis and colorectal cancer? Infect. Agents Cancer 2018, 13, 1. [Google Scholar] [CrossRef]
- Mameli, G.; Cocco, E.; Frau, J.; Marrosu, M.G.; Sechi, L.A. Epstein Barr Virus and Mycobacterium avium subsp. paratuberculosis peptides are recognized in sera and cerebrospinal fluid of MS patients. Sci. Rep. 2016, 6, 22401. [Google Scholar] [CrossRef] [Green Version]
- Niegowska, M.; Rapini, N.; Piccinini, S.; Mameli, G.; Caggiu, E.; Bitti, M.L.M.; Sechi, L.A. Type 1 Diabetes at-risk children highly recognize Mycobacterium avium subspecies paratuberculosis epitopes homologous to human Znt8 and Proinsulin. Sci. Rep. 2016, 6, 22266. [Google Scholar] [CrossRef] [Green Version]
- Bo, M.; Erre, G.L.; Niegowska, M.; Piras, M.; Taras, L.; Longu, M.G.; Passiu, G.; Sechi, L.A. Interferon regulatory factor 5 is a potential target of autoimmune response triggered by Epstein-Barr virus and Mycobacterium avium subsp. paratuberculosis in rheumatoid arthritis: Investigating a mechanism of molecular mimicry. Clin. Exp. Rheumatol. 2018, 36, 376–381. [Google Scholar]
- Naito, T.; Yokoyama, N.; Kakuta, Y.; Ueno, K.; Kawai, Y.; Onodera, M.; Moroi, R.; Kuroha, M.; Kanazawa, Y.; Kimura, T.; et al. Clinical and genetic risk factors for decreased bone mineral density in Japanese patients with inflammatory bowel disease. J. Gastroenterol. Hepatol. 2018, 33, 1873–1881. [Google Scholar] [CrossRef]
- Garrido, J.M.; Vazquez, P.; Molina, E.; Plazaola, J.M.; Sevilla, I.A.; Geijo, M.V.; Alonso-Hearn, M.; Juste, R.A. Paratuberculosis vaccination causes only limited cross-reactivity in the skin test for diagnosis of bovine tuberculosis. PLoS ONE 2013, 8, e80985. [Google Scholar] [CrossRef]
- Garry, F. Control of Paratuberculosis in Dairy Herds. Vet. Clin. Food Anim. Pract. 2011, 27, 599–607. [Google Scholar] [CrossRef]
- Groenendaal, H.; Nielen, M.; Jalvingh, A.W.; Horst, S.H.; Galligan, D.T.; Hesselink, J.W. A simulation of Johne’s disease control. Prev. Vet. Med. 2002, 54, 225–245. [Google Scholar] [CrossRef]
- Alonso-Hearn, M.; Badia-Bringué, G.; Canive, M. Genome-wide association studies for the identification of cattle susceptible and resilient to paratuberculosis. Front. Vet. Sci. 2022, 9, 935133. [Google Scholar] [CrossRef]
- Kiser, J.N.; White, S.N.; Johnson, K.A.; Hoff, J.L.; Taylor, J.F.; Neibergs, H.L. Identification of loci associated with susceptibility to Mycobacterium avium subspecies paratuberculosis (Map) tissue infection in cattle. J. Anim. Sci. 2017, 95, 1080–1091. [Google Scholar] [CrossRef] [Green Version]
- McGovern, S.P.; Purfield, D.C.; Ring, S.C.; Carthy, T.R.; Graham, D.A.; Berry, D.P. Candidate genes associated with the heritable humoral response to Mycobacterium avium ssp. paratuberculosis in dairy cows have factors in common with gastrointestinal diseases in humans. J. Dairy Sci. 2019, 102, 4249–4263. [Google Scholar] [CrossRef] [Green Version]
- Canive, M.; Badia-Bringué, G.; Vázquez, P.; González-Recio, O.; Fernández, A.; Garrido, J.M.; Juste, R.A.; Alonso-Hearn, M. Identification of loci associated with pathological outcomes in Holstein cattle infected with Mycobacterium avium subsp. paratuberculosis using whole-genome sequence data. Sci. Rep. 2021, 11, 20177. [Google Scholar] [CrossRef]
- Canive, M.; González-Recio, O.; Fernández, A.; Vázquez, P.; Badia-Bringué, G.; Lavín, J.L.; Garrido, J.M.; Juste, R.A.; Alonso-Hearn, M. dentification of loci associated with susceptibility to Mycobacterium avium subsp. paratuberculosis infection in Holstein cattle using combinations of diagnostic tests and imputed whole-genome sequence data. PLoS ONE 2021, 16, e0256091. [Google Scholar] [CrossRef]
- Aryngaziyev, B.; Beltramo, C.; Dondo, A.; Karymsakov, T.; Varello, K.; Goria, M.; Di Blasio, A.; Nodari, S.; Colussi, S.; Modesto, P.; et al. Polymorphisms associated to bovine paratuberculosis: Investigation of their role in dna-protein interactions and transcriptional regulation. Vet. Ital. 2020, 56, 109–114. [Google Scholar] [CrossRef]
- Kiser, J.N.; Wang, Z.; Zanella, R.; Scraggs, E.; Neupane, M.; Cantrell, B.; Van Tassell, C.P.; White, S.N.; Taylor, J.F.; Neibergs, H.L. Functional Variants Surrounding Endothelin 2 Are Associated with Mycobacterium avium Subspecies paratuberculosis Infection. Front. Vet. Sci. 2021, 8, 280. [Google Scholar] [CrossRef]
- Jansen, R.C.; Nap, J.P. Genetical genomics: The added value from segregation. Trends Genet. 2001, 17, 388–391. [Google Scholar] [CrossRef] [Green Version]
- Canive, M.; Fernandez-Jimenez, N.; Casais, R.; Vázquez, P.; Lavín, J.L.; Bilbao, J.R.; Blanco-Vázquez, C.; Garrido, J.M.; Juste, R.A.; Alonso-Hearn, M. Identification of loci associated with susceptibility to bovine paratuberculosis and with the dysregulation of the MECOM, eEF1A2, and U1 spliceosomal RNA expression. Sci. Rep. 2021, 11, 313. [Google Scholar] [CrossRef] [PubMed]
- Conus, S.; Simon, H.U. Cathepsins and their involvement in immune responses. Swiss Med. Wkly 2010, 140, w13042. [Google Scholar] [CrossRef] [PubMed]
- Turk, V. NEW EMBO MEMBERS’ REVIEW: Lysosomal cysteine proteases: Facts and opportunities. EMBO J. 2001, 20, 4629–4633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, A.; Deveraux, Q.; Turk, B.; Sali, A. Comprehensive search for cysteine cathepsins in the human genome. Biol. Chem. 2004, 385, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Chwieralski, C.E.; Welte, T.; Bühling, F. Cathepsin-regulated apoptosis. Apoptosis 2006, 11, 143–149. [Google Scholar] [CrossRef]
- Turk, V.; Stoka, V.; Vasiljeva, O.; Renko, M.; Sun, T.; Turk, B.; Turk, D. Cysteine cathepsins: From structure, function and regulation to new frontiers. Biochim. Biophys. Acta Proteins Proteom. 2012, 1824, 68–88. [Google Scholar] [CrossRef] [Green Version]
- Szulc-Dąbrowska, L.; Bossowska-Nowicka, M.; Struzik, J.; Toka, F.N. Cathepsins in Bacteria-Macrophage Interaction: Defenders or Victims of Circumstance? Front. Cell. Infect. Microbiol. 2020, 10, 601072. [Google Scholar] [CrossRef]
- Vázquez, P.; Ruiz-Larrañaga, O.; Garrido, J.M.; Iriondo, M.; Manzano, C.; Agirre, M.; Estonba, A.; Juste, R.A. Genetic association analysis of paratuberculosis forms in Holstein-Friesian cattle. Vet. Med. Int. 2014, 2014, 321327. [Google Scholar] [CrossRef] [Green Version]
- Juste, R.A.; Alonso-Hearn, M.; Garrido, J.M.; Abendaño, N.; Sevilla, I.A.; Gortazar, C.; De La Fuente, J.; Dominguez, L. Increased lytic efficiency of bovine macrophages trained with killed mycobacteria. PLoS ONE 2016, 11, e0165607. [Google Scholar] [CrossRef] [Green Version]
- Abendaño, N.; Sevilla, I.; Prieto, J.M.; Garrido, J.M.; Juste, R.A.; Alonso-Hearn, M. Quantification of Mycobacterium avium subsp. paratuberculosis strains representing distinct genotypes and isolated from domestic and wildlife animal species by use of an automatic liquid culture system. J. Clin. Microbiol. 2012, 50, 2609–2617. [Google Scholar] [CrossRef] [Green Version]
- Canive, M.; Badia-Bringué, G.; Vazquez, P.; Garrido, J.M.; Juste, R.A.; Fernandez, A.; Gonzalez-Recio, Ó.; Alonso-Hearn, M. A Genome-Wide Association Study for Tolerance to Paratuberculosis Identifies Candidate Genes Involved in DNA Packaging, DNA Damage Repair, Innate Immunity, and Pathogen Persistence. Front. Immunol. 2022, 13, 820965. [Google Scholar] [CrossRef]
- Abendaño, N.; Juste, R.A.; Alonso-Hearn, M. Anti-inflammatory and antiapoptotic responses to infection: A common denominator of human and bovine macrophages infected with Mycobacterium avium subsp. paratuberculosis. BioMed Res. Int. 2013, 2013, 908348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abendaño, N.; Tyukalova, L.; Barandika, J.F.; Balseiro, A.; Sevilla, I.A.; Garrido, J.M.; Juste, R.A.; Alonso-Hearn, M. Mycobacterium avium subsp. paratuberculosis isolates induce in vitro granuloma formation and show successful survival phenotype, common anti-inflammatory and antiapoptotic responses within ovine macrophages regardless of genotype or host of origin. PLoS ONE 2014, 9, e104238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, L.E.; Abendaño, N.; Juste, R.A.; Alonso-Hearn, M. Three-dimensional in vitro models of granuloma to study bacteria-host interactions, drug-susceptibility, and resuscitation of dormant mycobacteria. BioMed Res. Int. 2014, 2014, 623856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Hearn, M.; Canive, M.; Blanco-Vazquez, C.; Torremocha, R.; Balseiro, A.; Amado, J.; Varela-Martinez, E.; Ramos, R.; Jugo, B.M.; Casais, R. RNA-Seq analysis of ileocecal valve and peripheral blood from Holstein cattle infected with Mycobacterium avium subsp. paratuberculosis revealed dysregulation of the CXCL8/IL8 signaling pathway. Sci. Rep. 2019, 9, 14845. [Google Scholar] [CrossRef] [Green Version]
- Badia-Bringué, G.; Canive, M.; Casais-Goyos, R.; Blanco-Vázquez, C.; Amado, J.; Iglesias, N.; González, A.; Bascones, M.; Juste, R.A.; Alonso-Hearn, M. Evaluation of a droplet digital PCR (ddPCR) assay for quantification of Mycobacterium avium subsp. paratuberculosis (MAP) DNA in whole-blood and fecal samples from MAP-infected Holstein cattle. Front. Vet. Sci. 2022, 9, 944189. [Google Scholar] [CrossRef]
- Blanco-Vázquez, C.; Alonso-Hearn, M.; Iglesias, N.; Vázquez, P.; Juste, R.A.; Garrido, J.M.; Balseiro, A.; Canive, M.; Amado, J.; Queipo, M.A.; et al. Use of ATP-binding cassette subfamily A member 13 (ABCA13) for sensitive detection of focal pathological forms of subclinical bovine paratuberculosis. Front. Vet. Sci. 2022, 9, 816135. [Google Scholar] [CrossRef]
- Arranz-Trullén, J.; Lu, L.; Pulido, D.; Bhakta, S.; Boix, E. Host antimicrobial peptides: The promise of new treatment strategies against tuberculosis. Front. Immunol. 2017, 8, 1499. [Google Scholar] [CrossRef] [Green Version]
- Cirone, K.M.; Lahiri, P.; Holani, R.; Tan, Y.L.; Arrazuria, R.; De Buck, J.; Barkema, H.W.; Cobo, E.R. Synthetic cathelicidin LL-37 reduces Mycobacterium avium subsp. paratuberculosis internalization and pro-inflammatory cytokines in macrophages. Cell Tissue Res. 2020, 379, 207–217. [Google Scholar] [CrossRef]
- Odeberg, H.; Olsson, I. Antibacterial activity of cationic proteins from human granulocytes. J. Clin. Investig. 1975, 56, 1118–1124. [Google Scholar] [CrossRef]
- Bangalore, N.; Travis, J.; Onunka, V.C.; Pohl, J.; Shafer, W.M. Identification of the primary antimicrobial domains in human neutrophil Cathepsin G. J. Biol. Chem. 1990, 265, 13584–13588. [Google Scholar] [CrossRef]
- Kretschmer, D.; Breitmeyer, R.; Gekeler, C.; Lebtig, M.; Schlatterer, K.; Nega, M.; Stahl, M.; Stapels, D.; Rooijakkers, S.; Peschel, A. Staphylococcus aureus Depends on Eap Proteins for Preventing Degradation of Its Phenol-Soluble Modulin Toxins by Neutrophil Serine Proteases. Front. Immunol. 2021, 12, 701093. [Google Scholar] [CrossRef] [PubMed]
- Reeves, E.P.; Lu, H.; Jacobs, H.L.; Messina, C.G.M.; Bolsover, S.; Gabellall, G.; Potma, E.O.; Warley, A.; Roes, J.; Segal, A.W. Killing activity of neutrophils is mediated through activation of proteases by K+ flux. Nature 2002, 416, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Burster, T.; Macmillan, H.; Hou, T.; Boehm, B.O.; Mellins, E.D. Cathepsin G: Roles in antigen presentation and beyond. Mol. Immunol. 2010, 47, 658–665. [Google Scholar] [CrossRef] [Green Version]
- Pires, D.; Marques, J.; Pombo, J.P.; Carmo, N.; Bettencourt, P.; Neyrolles, O.; Lugo-Villarino, G.; Anes, E. Role of Cathepsins in Mycobacterium tuberculosis Survival in Human Macrophages. Sci. Rep. 2016, 6, 32247. [Google Scholar] [CrossRef] [Green Version]
- Danelishvili, L.; Everman, J.L.; McNamara, M.J.; Bermudez, L.E. Inhibition of the plasma-membrane-associated serine protease Cathepsin G by Mycobacterium tuberculosis Rv3364c suppresses caspase-1 and pyroptosis in macrophages. Front. Microbiol. 2012, 2, 281. [Google Scholar] [CrossRef] [Green Version]
- Walter, K.; Steinwede, K.; Aly, S.; Reinheckel, T.; Bohling, J.; Maus, U.A.; Ehlers, S. Cathepsin G in Experimental Tuberculosis: Relevance for Antibacterial Protection and Potential for Immunotherapy. J. Immunol. 2015, 195, 3325–3333. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Marrero, C.A.; Stewart, J.; Shafer, W.M.; Roman, J. The down-regulation of Cathepsin G in THP-1 monocytes after infection with Mycobacterium tuberculosis is associated with increased intracellular survival of bacilli. Infect. Immun. 2004, 72, 5712–5721. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, M.; Meinders, A.; Steinwede, K.; Maus, R.; Lucke, N.; Bühling, F.; Ehlers, S.; Welte, T.; Maus, U.A. Mediator responses of alveolar macrophages and kinetics of mononuclear phagocyte subset recruitment during acute primary and secondary mycobacterial infections in the lungs of mice. Cell. Microbiol. 2007, 9, 738–752. [Google Scholar] [CrossRef]
- Steinwede, K.; Maus, R.; Bohling, J.; Voedisch, S.; Braun, A.; Ochs, M.; Schmiedl, A.; Länger, F.; Gauthier, F.; Roes, J.; et al. Cathepsin G and Neutrophil Elastase Contribute to Lung-Protective Immunity against Mycobacterial Infections in Mice. J. Immunol. 2012, 188, 4476–4487. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| No Lesion (N) | Frequency (%) | Lesion (N) | Frequency (%) | OR (95% CI) | |
|---|---|---|---|---|---|
| Codominant | |||||
| AA | 422 | 84.2 | 369 | 83.5 | 1 |
| AC | 68 | 13.6 | 67 | 15.2 | 1.13 (0.78–1.62) |
| CC | 11 | 2.2 | 6 | 1.4 | 0.62 (0.23–1.7) |
| Dominant | |||||
| AA | 422 | 84.2 | 369 | 83.5 | 1 |
| AC-CC | 79 | 15.8 | 73 | 16.5 | 1.06 (0.75–1.5) |
| Recessive | |||||
| AA-AC | 490 | 97.8 | 436 | 98.6 | 1 |
| C/C | 11 | 2.2 | 6 | 1.4 | 0.61 (0.22–1.67) |
| Overdominant | |||||
| AA-CC | 433 | 86.4 | 375 | 84.8 | 1 |
| AC | 68 | 13.6 | 67 | 15.2 | 1.14 (0.79–1.64) |
| log-Additive | |||||
| 0, 1, 2 | 501 | 53.1 | 442 | 46.9 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Canive, M.; Badia-Bringué, G.; Alonso-Hearn, M. The Upregulation of Cathepsin G Is Associated with Resistance to Bovine Paratuberculosis. Animals 2022, 12, 3038. https://doi.org/10.3390/ani12213038
Canive M, Badia-Bringué G, Alonso-Hearn M. The Upregulation of Cathepsin G Is Associated with Resistance to Bovine Paratuberculosis. Animals. 2022; 12(21):3038. https://doi.org/10.3390/ani12213038
Chicago/Turabian StyleCanive, Maria, Gerard Badia-Bringué, and Marta Alonso-Hearn. 2022. "The Upregulation of Cathepsin G Is Associated with Resistance to Bovine Paratuberculosis" Animals 12, no. 21: 3038. https://doi.org/10.3390/ani12213038