
Dietary Strategies to Modulate the Health Condition and Immune Responses in Gilthead Seabream (Sparus aurata) Juveniles Following Intestinal Inflammation

 , , , and
, , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
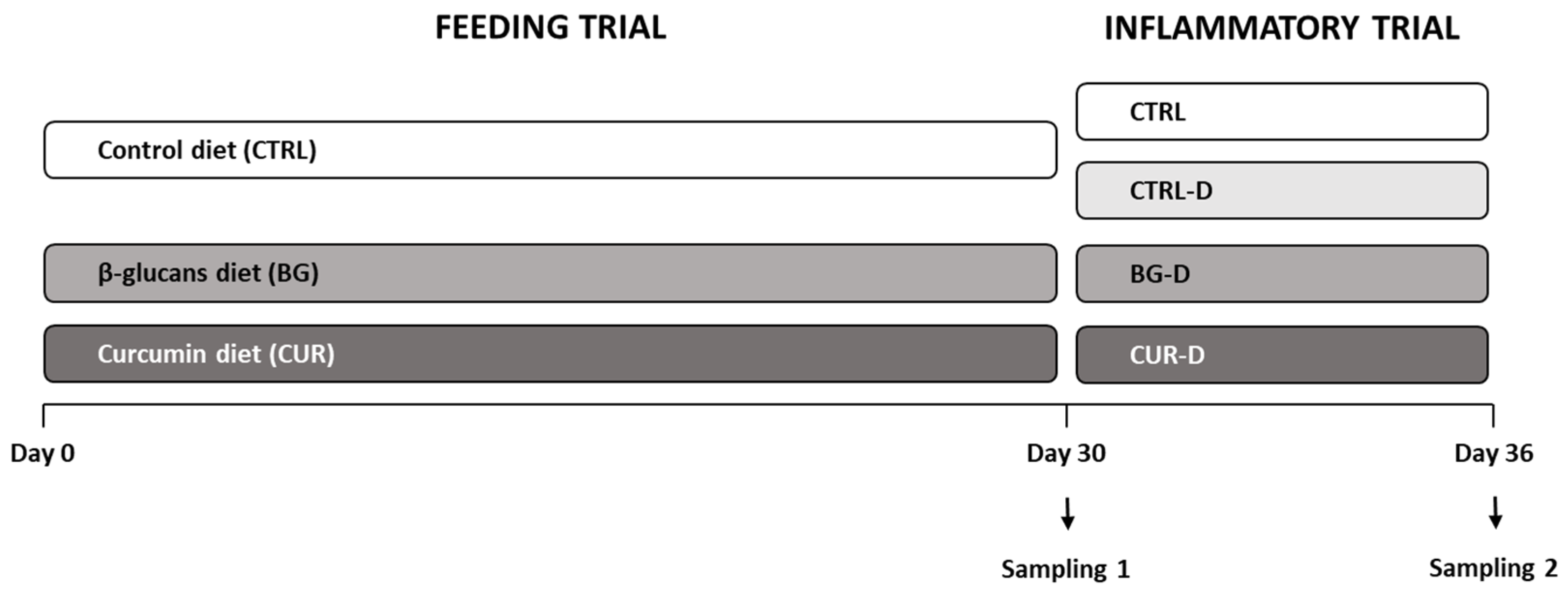
2.2. Fish and Experimental Design
2.3. Sample Collection
2.4. Hematological Profile
2.5. Plasma and Intestinal Humoral Immune Parameters
2.6. Liver and Intestinal Oxidative Stress
2.7. Intestinal Gene Expression
2.8. Statistical Analysis
3. Results
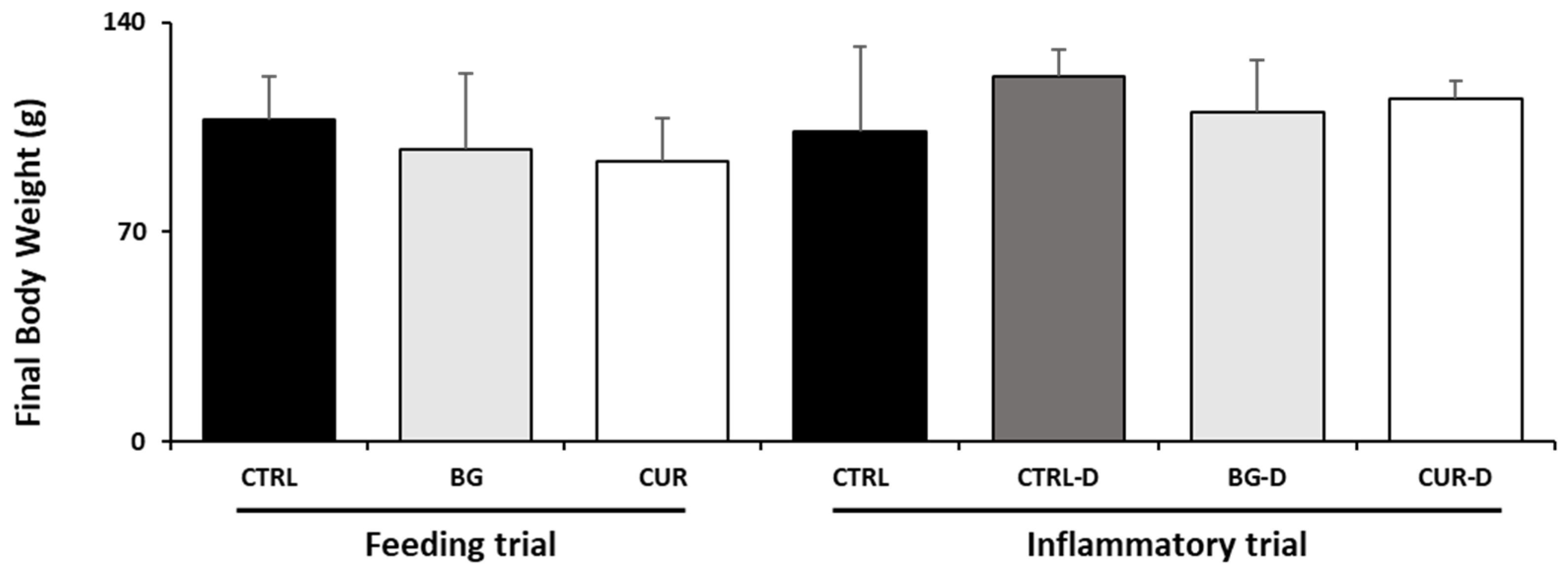
3.1. Growth Performance
3.2. Hematological Profile
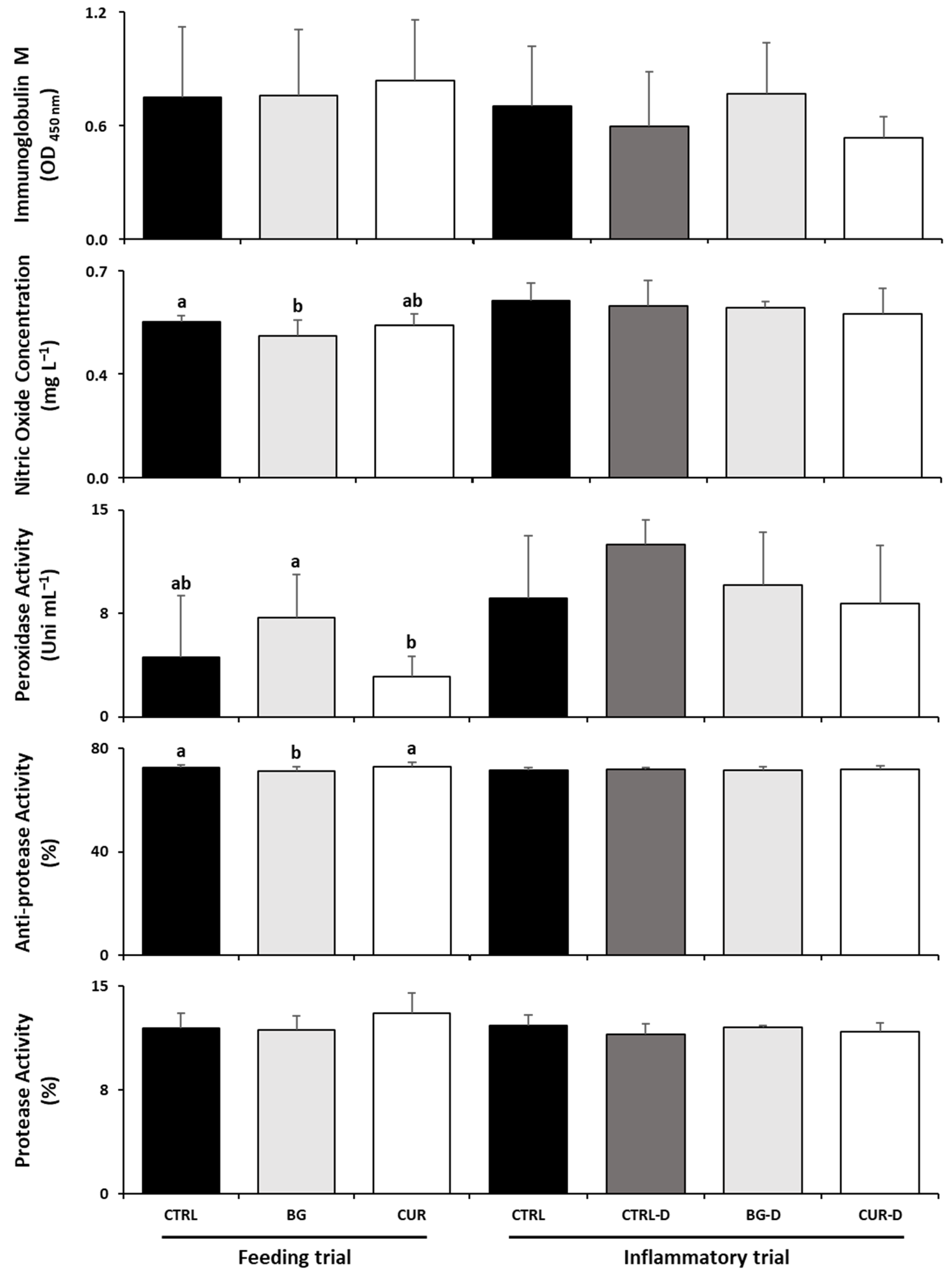
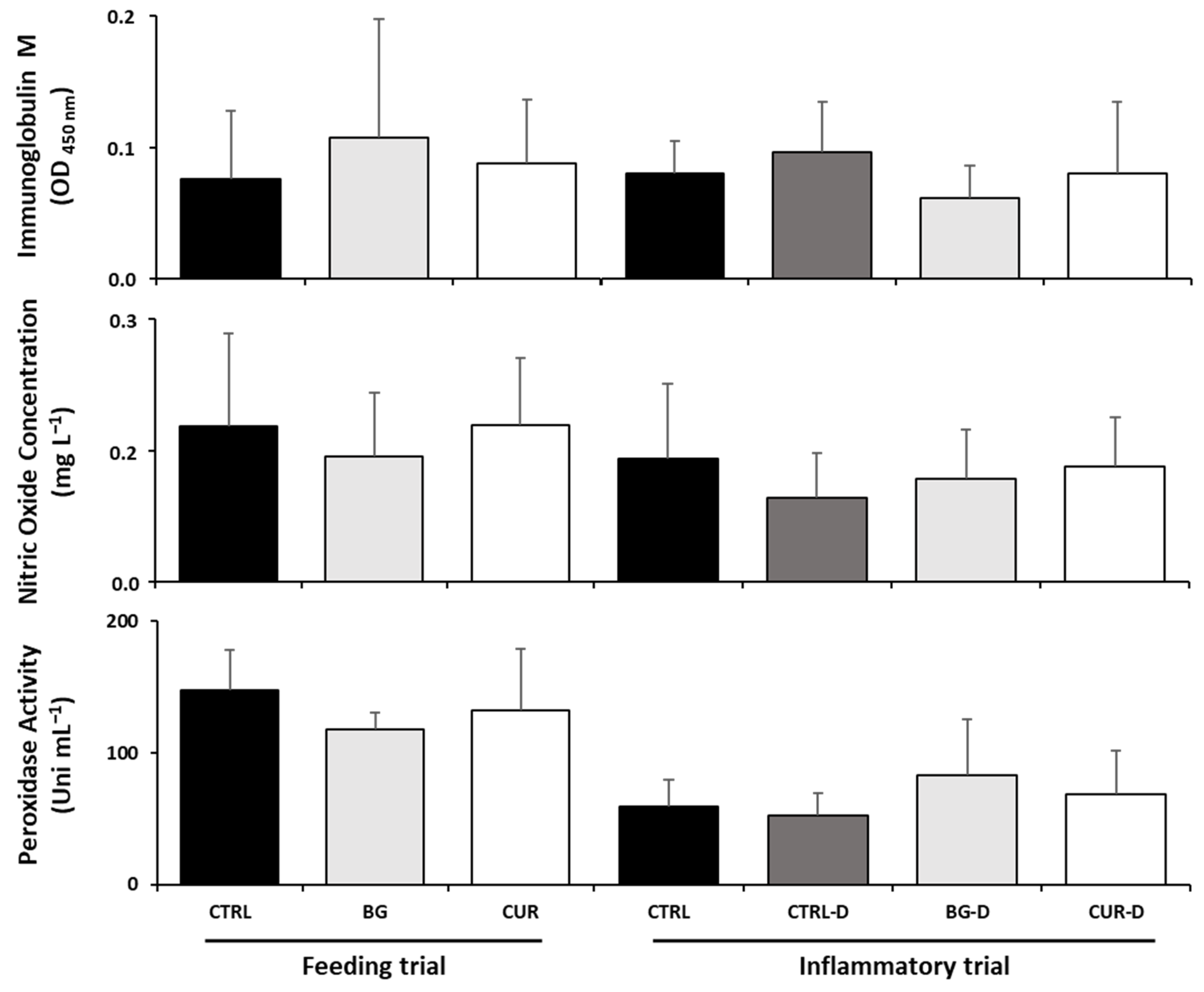
3.3. Plasma Humoral Immunological Parameters
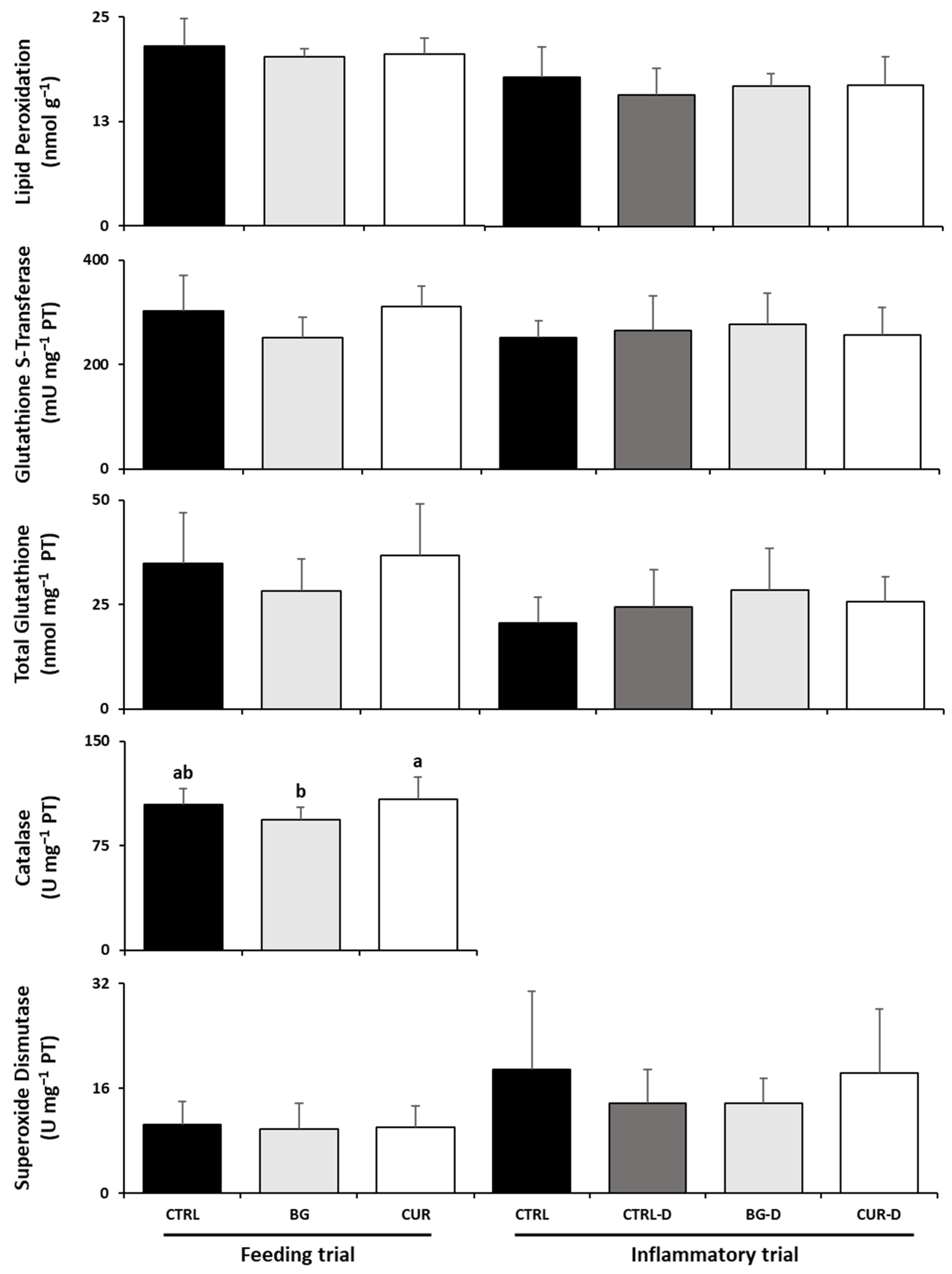
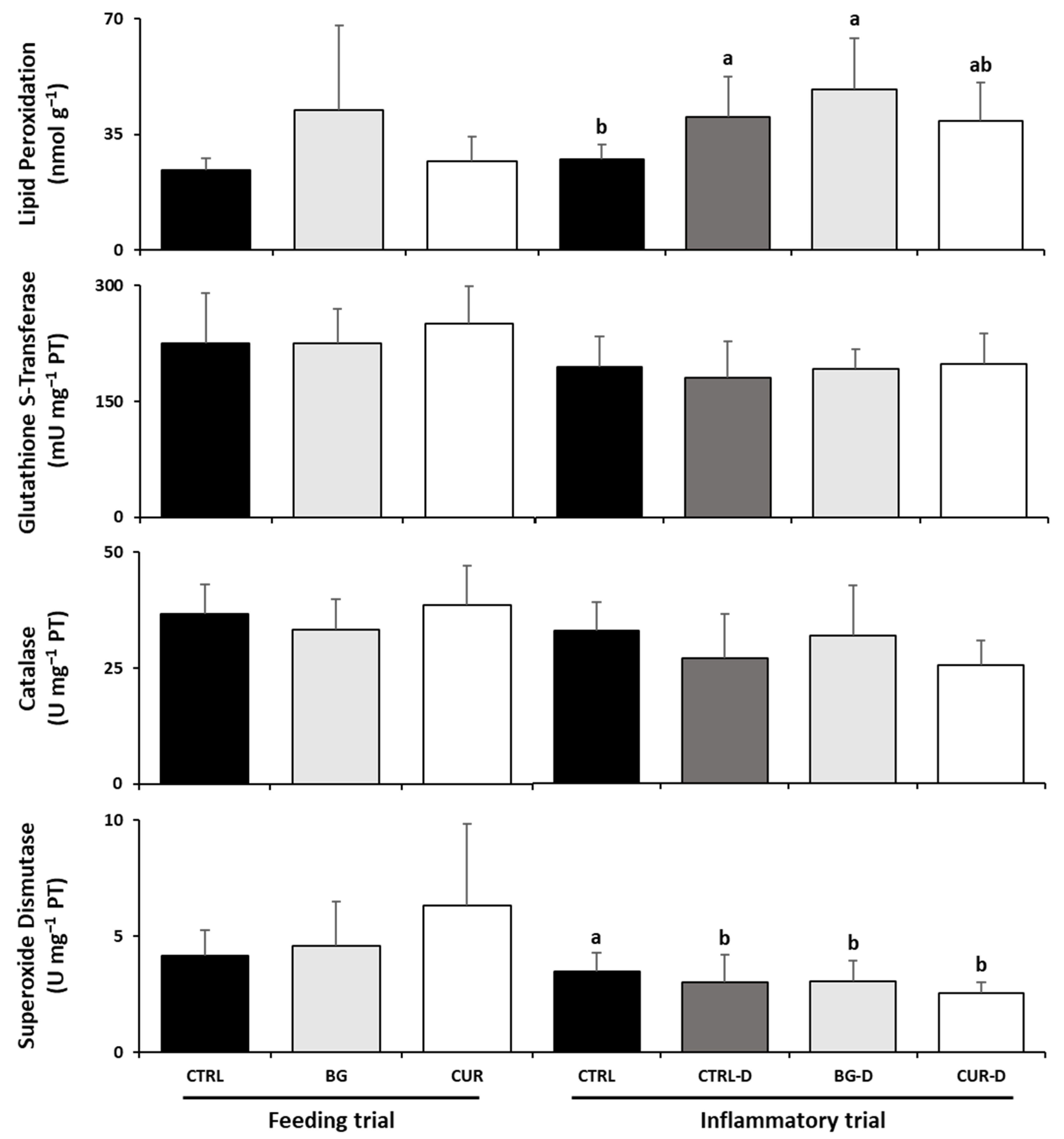
3.4. Liver Oxidative Stress
3.5. Intestinal Humoral Immunological Parameters
3.6. Intestinal Oxidative Stress
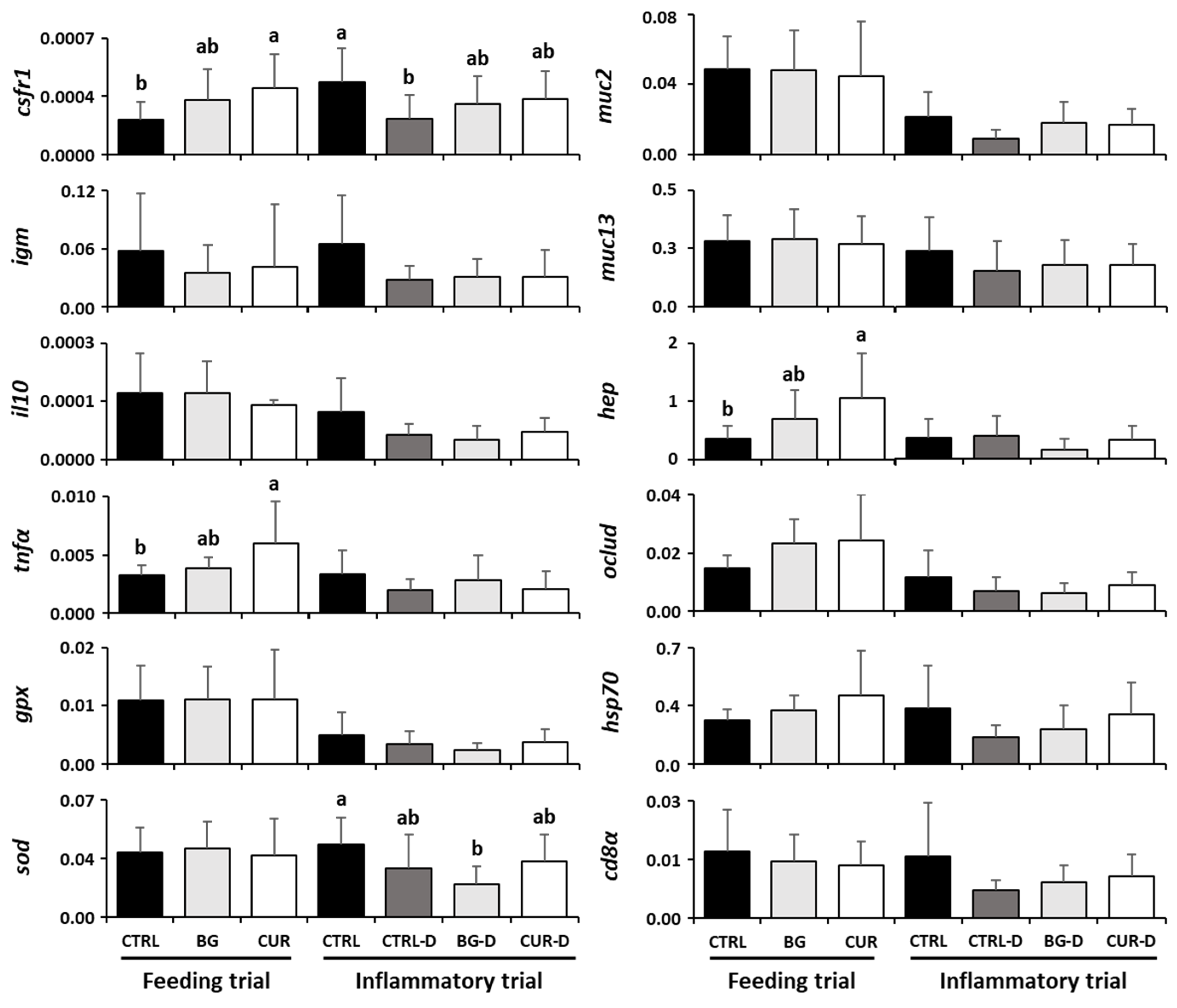
3.7. Intestinal Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vetvicka, V.; Vannucci, L.; Sima, P. The effects of β—Glucan on fish immunity. N. Am. J. Med. Sci. 2013, 5, 580–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ching, J.J.; Shuib, A.S.; Majid, N.A.; Taufek, N.M. Immunomodulatory activity of β-glucans in fish: Relationship between β-glucan administration parameters and immune response induced. Aquac. Res. 2020, 52, 1824–1845. [Google Scholar] [CrossRef]
- Ashry, A.; Hassan, A.; Habiba, M.; El-Zayat, A.; El-Sharnouby, M.; Sewilam, H.; Dawood, M. The Impact of Dietary Curcumin on the Growth Performance, Intestinal Antibacterial Capacity, and Haemato-Biochemical Parameters of Gilthead Seabream (Sparus aurata). Animals 2021, 11, 1779. [Google Scholar] [CrossRef] [PubMed]
- Meena, D.K.; Das, P.; Kumar, S.; Mandal, S.C.; Prusty, A.; Singh, S.K.; Akhtar, M.S.; Behera, B.K.; Kumar, K.; Pal, A.K.; et al. Beta-glucan: An ideal immunostimulant in aquaculture (a review). Fish Physiol. Biochem. 2012, 39, 431–457. [Google Scholar] [CrossRef]
- Tacon, A.G.J.; Metian, M. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects. Aquaculture 2008, 285, 146–158. [Google Scholar] [CrossRef]
- Hamid, S.N.I.N.; Yusof, S.J.H.M.; Zakaria, Z.; Abdullah, R. Evaluation of Potential Alternative Ingredients for Formulation of Fish Feed. Appl. Mech. Mater. 2015, 754–755, 1081–1086. [Google Scholar] [CrossRef]
- FAO. Use of Fishmeal and Fish Oil in Aquafeeds: Further Thoughts on the Fishmeal Trap; FAO Fisheries Circular No. 975; FAO: Rome, Italy, 2002; Available online: https://www.fao.org/3/y3781e/y3781e.pdf (accessed on 1 September 2021).
- Estruch, G.; Collado, M.C.; Monge-Ortiz, R.; Tomás-Vidal, A.; Jover-Cerdá, M.; Peñaranda, D.S.; Martínez, G.P.; Martínez-Llorens, S. Long-term feeding with high plant protein based diets in gilthead seabream (Sparus aurata, L.) leads to changes in the inflammatory and immune related gene expression at intestinal level. BMC Vet. Res. 2018, 14, 302. [Google Scholar] [CrossRef]
- Egerton, S.; Wan, A.; Murphy, K.; Collins, F.; Ahern, G.; Sugrue, I.; Busca, K.; Egan, F.; Muller, N.; Whooley, J.; et al. Replacing fishmeal with plant protein in Atlantic salmon (Salmo salar) diets by supplementation with fish protein hydrolysate. Sci. Rep. 2020, 10, 4194. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Sakhawat Hossain, M.; Ragaza, J.A.; Rubio Benito, M. The potential impacts of soy protein on fish gut health. In Soybean for Human Consumption and Animal Feed; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran Sulfate Sodium (DSS)-Induced Colitis in Mice. Curr. Protoc. Immunol. 2014, 104, 15.25.1–15.25.14. [Google Scholar] [CrossRef]
- Jialing, L.; Yangyang, G.; Jing, Z.; Xiaoyi, T.; Ping, W.; Liwei, S.; Simin, C. Changes in serum inflammatory cytokine levels and intestinal flora in a self-healing dextran sodium sulfate-induced ulcerative colitis murine model. Life Sci. 2020, 263, 118587. [Google Scholar] [CrossRef]
- Arslan, G.; Erichsen, K.; Milde, A.M.; Helgeland, L.; Bjørkkjær, T.; Frøyland, L.; Berstad, A. No Protection against DSS-induced Colitis by Short-term Pretreatment with Seal or Fish Oils in Rats. Integr. Med. Insights 2007, 2, 25–34. [Google Scholar] [CrossRef]
- Munyaka, P.M.; Rabbi, M.F.; Khafipour, E.; Ghia, J.-E. Acute dextran sulfate sodium (DSS)-induced colitis promotes gut microbial dysbiosis in mice. J. Basic Microbiol. 2016, 56, 986–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.-S.; Choi, J.; Hwang, S.-H.; Kim, J.-K.; Kim, E.-K.; Lee, S.-Y.; Lee, B.-I.; Park, S.-H.; Cho, M.-L. Cottonseed Oil Protects Against Intestinal Inflammation in Dextran Sodium Sulfate-Induced Inflammatory Bowel Disease. J. Med. Food 2019, 22, 672–679. [Google Scholar] [CrossRef]
- Silvestri, C.; Pagano, E.; Lacroix, S.; Venneri, T.; Cristiano, C.; Calignano, A.; Parisi, O.A.; Izzo, A.A.; Di Marzo, V.; Borrelli, F. Fish Oil, Cannabidiol and the Gut Microbiota: An Investigation in a Murine Model of Colitis. Front. Pharmacol. 2020, 11, 585096. [Google Scholar] [CrossRef] [PubMed]
- Daskalaki, M.; Axarlis, K.; Aspevik, T.; Orfanakis, M.; Kolliniati, O.; Lapi, I.; Tzardi, M.; Dermitzaki, E.; Venihaki, M.; Kousoulaki, K.; et al. Fish Sidestream-Derived Protein Hydrolysates Suppress DSS-Induced Colitis by Modulating Intestinal Inflammation in Mice. Mar. Drugs 2021, 19, 312. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.-W.; Zhou, X.-L.; Wang, R.; Shu, C.-H.; Zhou, Y.-F.; Ying, X.-G.; Zheng, B. Protective Effect of Tuna Bioactive Peptide on Dextran Sulfate Sodium-Induced Colitis in Mice. Mar. Drugs 2021, 19, 127. [Google Scholar] [CrossRef]
- Oehlers, S.; Flores, M.V.; Hall, C.; Crosier, K.E.; Crosier, P.S. Retinoic acid suppresses intestinal mucus production and exacerbates experimental enterocolitis. Dis. Model. Mech. 2012, 5, 457–467. [Google Scholar] [CrossRef] [Green Version]
- Oehlers, S.; Flores, M.V.; Hall, C.; Okuda, K.S.; Sison, J.O.; Crosier, K.E.; Crosier, P.S. Chemically Induced Intestinal Damage Models in Zebrafish Larvae. Zebrafish 2013, 10, 184–193. [Google Scholar] [CrossRef]
- Wang, P.; Lu, Y.-Q.; Wen, Y.; Yu, D.-Y.; Ge, L.; Dong, W.-R.; Xiang, L.-X.; Shao, J.-Z. IL-16 Induces Intestinal Inflammation via PepT1 Upregulation in a Pufferfish Model: New Insights into the Molecular Mechanism of Inflammatory Bowel Disease. J. Immunol. 2013, 191, 1413–1427. [Google Scholar] [CrossRef]
- Marjoram, L.; Bagnat, M. Infection, Inflammation and Healing in Zebrafish: Intestinal Inflammation. Curr. Pathobiol. Rep. 2015, 3, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Fenero, C.M.; Idelí, C.; Fé, M. Inflammatory diseases modelling in zebrafish. World J. Exp. Med. 2016, 6, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Hanyang, L.; Xuanzhe, L.; Xuyang, C.; Yujia, Q.; Jiarong, F.; Jun, S.; Zhihua, R. Application of Zebrafish Models in Inflammatory Bowel Disease. Front. Immunol. 2017, 8, 501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.-W.; Ho, Y.-J.; Ciou, S.-C.; Gong, Z. Innovative Disease Model: Zebrafish as an In Vivo Platform for Intestinal Disorder and Tumors. Biomedicines 2017, 5, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, L.-S.; Morrison, J.; Hsu, N.-Y.; Labrias, P.R.; Nayar, S.; Chen, E.; Villaverde, N.; Facey, J.A.; Boschetti, G.; Giri, M.; et al. Zebrafish modeling of intestinal injury, bacterial exposures, and medications defines epithelial in vivo responses relevant to human inflammatory bowel disease. Dis. Model. Mech. 2019, 12, dmm037432. [Google Scholar] [CrossRef] [Green Version]
- Perše, M.; Cerar, A. Dextran Sodium Sulphate Colitis Mouse Model: Traps and Tricks. J. Biomed. Biotechnol. 2012, 2012, 718617. [Google Scholar] [CrossRef] [Green Version]
- Gannam, A.L.; Schrock, R.M. Immunostimulants in Fish Diets. J. Appl. Aquac. 1999, 9, 53–89. [Google Scholar] [CrossRef]
- Sakai, M. Current research status of fish immunostimulants. Aquaculture 1999, 172, 63–92. [Google Scholar] [CrossRef]
- Ringø, E.; Olsen, R.E.; Wadsworth, S.; Vecino, J.L.G. Use of Immunostimulants and Nucleotides in Aquaculture: A Review. J. Mar. Sci. Res. Dev. 2011, 2, 104. [Google Scholar] [CrossRef] [Green Version]
- Vallejos-Vidal, E.; Reyes-López, F.; Teles, M.; MacKenzie, S. The response of fish to immunostimulant diets. Fish Shellfish Immunol. 2016, 56, 34–69. [Google Scholar] [CrossRef]
- Dawood, M.A.; Koshio, S.; Esteban, M. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2017, 10, 950–974. [Google Scholar] [CrossRef]
- Koch, J.F.A.; de Oliveira, C.A.F.; Zanuzzo, F.S. Dietary β-glucan (MacroGard®) improves innate immune responses and disease resistance in Nile tilapia regardless of the administration period. Fish Shellfish Immunol. 2021, 112, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.V.; Zanuzzo, F.S.; Koch, J.F.A.; De Oliveira, C.A.F.; Sima, P.; Vetvicka, V. Development of Fish Immunity and the Role of β-Glucan in Immune Responses. Molecules 2020, 25, 5378. [Google Scholar] [CrossRef]
- Granum, E.; Myklestad, S.M. A simple combined method for determination of β-1,3-glucan and cell wall polysaccharides in diatoms. Hydrobiologia 2002, 477, 155–161. [Google Scholar] [CrossRef]
- Chiovitti, A.; Molino, P.; Crawford, S.A.; Teng, R.; Spurck, T.; Wetherbee, R. The glucans extracted with warm water from diatoms are mainly derived from intracellular chrysolaminaran and not extracellular polysaccharides. Eur. J. Phycol. 2004, 39, 117–128. [Google Scholar] [CrossRef]
- Carballo, C.; Chronopoulou, E.G.; Letsiou, S.; Maya, C.; Labrou, N.E.; Infante, C.; Power, D.; Manchado, M. Antioxidant capacity and immunomodulatory effects of a chrysolaminarin-enriched extract in Senegalese sole. Fish Shellfish Immunol. 2018, 82, 1–8. [Google Scholar] [CrossRef]
- Reis, B.; Gonçalves, A.T.; Santos, P.; Sardinha, M.; Conceição, L.E.C.; Serradeiro, R.; Pérez-Sánchez, J.; Calduch-Giner, J.; Schmid-Staiger, U.; Frick, K.; et al. Immune Status and Hepatic Antioxidant Capacity of Gilthead Seabream Sparus aurata Juveniles Fed Yeast and Microalga Derived β-glucans. Mar. Drugs 2021, 19, 653. [Google Scholar] [CrossRef]
- Skjermo, J.; Størseth, T.R.; Hansen, K.; Handå, A.; Øie, G. Evaluation of β-(1→3, 1→6)-glucans and High-M alginate used as immunostimulatory dietary supplement during first feeding and weaning of Atlantic cod (Gadus morhua L.). Aquaculture 2006, 261, 1088–1101. [Google Scholar] [CrossRef]
- Ali, B.H.; Marrif, H.; Noureldayem, S.A.; Bakheit, A.O.; Blunden, G. Some Biological Properties of Curcumin: A Review. Nat. Prod. Commun. 2006, 1, 1934578X0600100613. [Google Scholar] [CrossRef] [Green Version]
- Sahu, S.; Das, B.K.; Mishra, B.K.; Pradhan, J.; Samal, S.K.; Sarangi, N. Effect of dietary Curcuma longa on enzymatic and immunological profiles of rohu, Labeo rohita (Ham.), infected with Aeromonas hydrophila. Aquac. Res. 2008, 39, 1720–1730. [Google Scholar] [CrossRef]
- Behera, T.; Swain, P.; Sahoo, S.K.; Mohapatra, D.; Das, B.K. Immunostimulatory effects of curcumin in fish, Labeo rohita (H.). Indian J. Nat. Prod. Resour. 2011, 2, 184–188. [Google Scholar]
- Elgendy, M.; Hakim, A.; Ibrahim, T.; Soliman, W.; Ali, S. Immunomodulatory Effects of Curcumin on Nile Tilapia, Oreochromis niloticus and its Antimicrobial Properties against Vibrio alginolyticus. J. Fish. Aquat. Sci. 2016, 11, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Wu, X.-Y.; Zhou, X.-Q.; Feng, L.; Liu, Y.; Jiang, W.-D.; Wu, P.; Zhao, Y. Effects of dietary curcumin supplementation on growth performance, intestinal digestive enzyme activities and antioxidant capacity of crucian carp Carassius auratus. Aquaculture 2016, 463, 174–180. [Google Scholar] [CrossRef]
- Mahmoud, H.K.; Al-Sagheer, A.A.; Reda, F.M.; Mahgoub, S.A.; Ayyat, M.S. Dietary curcumin supplement influence on growth, immunity, antioxidant status, and resistance to Aeromonas hydrophila in Oreochromis niloticus. Aquaculture 2017, 475, 16–23. [Google Scholar] [CrossRef]
- Baldissera, M.D.; Souza, C.F.; Zeppenfeld, C.C.; Descovi, S.; Machado, V.S.; Santos, R.C.; Baldisserotto, B. Efficacy of dietary curcumin supplementation as bactericidal for silver catfish against Streptococcus agalactiae. Microb. Pathog. 2018, 116, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Yonar, M.E.; Yonar, S.M.; Ispir, U.; Ural, M. Effects of curcumin on haematological values, immunity, antioxidant status and resistance of rainbow trout (Oncorhynchus mykiss) against Aeromonas salmonicida subsp. achromogenes. Fish Shellfish Immunol. 2019, 89, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zhou, X.; Jiang, W.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.; Tang, L.; Shi, H.; Feng, L. Dietary curcumin supplementation enhanced growth performance, intestinal digestion, and absorption and amino acid transportation abilities in on-growing grass carp (Ctenopharyngodon idella). Aquac. Res. 2020, 51, 4863–4873. [Google Scholar] [CrossRef]
- Zheng, D.; Huang, C.; Huang, H.; Zhao, Y.; Khan, M.R.U.; Zhao, H.; Huang, L. Antibacterial Mechanism of Curcumin: A Review. Chem. Biodivers. 2020, 17, e2000171. [Google Scholar] [CrossRef]
- Mohamed, A.A.-R.; El-Houseiny, W.; El-Murr, A.E.; Ebraheim, L.L.; Ahmed, A.I.; El-Hakim, Y.M.A. Effect of hexavalent chromium exposure on the liver and kidney tissues related to the expression of CYP450 and GST genes of Oreochromis niloticus fish: Role of curcumin supplemented diet. Ecotoxicol. Environ. Saf. 2019, 188, 109890. [Google Scholar] [CrossRef]
- Alagawany, M.; Farag, M.R.; Abdelnour, S.A.; Dawood, M.A.; Elnesr, S.S.; Dhama, K. Curcumin and its different forms: A review on fish nutrition. Aquaculture 2020, 532, 736030. [Google Scholar] [CrossRef]
- Machado, M.; Azeredo, R.; Díaz-Rosales, P.; Afonso, A.; Peres, H.; Oliva-Teles, A.; Costas, B. Dietary tryptophan and methionine as modulators of European seabass (Dicentrarchus labrax) immune status and inflammatory response. Fish Shellfish Immunol. 2015, 42, 353–362. [Google Scholar] [CrossRef]
- Afonso, A.; Lousada, S.; Silva, J.; Ellis, A.E.; Silva, M.T. Neutrophil and macrophage responses to inflammation in the peritoneal cavity of rainbow trout Oncorhynchus mykiss. A light and electron microscopic cytochemical study. Dis. Aquat. Org. 1998, 34, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Pinto, L.; Martos-Sitcha, J.A.; Reis, B.; Azeredo, R.; Fernandez-Boo, S.; Pérez-Sánchez, J.; Calduch-Giner, J.A.; Engrola, S.; Conceição, L.E.; Dias, J.; et al. Dietary tryptophan supplementation induces a transient immune enhancement of gilthead seabream (Sparus aurata) juveniles fed fishmeal-free diets. Fish Shellfish Immunol. 2019, 93, 240–250. [Google Scholar] [CrossRef]
- Cuesta, A.; Meseguer, J.; Esteban, M. Total serum immunoglobulin M levels are affected by immunomodulators in seabream (Sparus aurata L.). Vet. Immunol. Immunopathol. 2004, 101, 203–210. [Google Scholar] [CrossRef]
- Ramos-Pinto, L.; Machado, M.; Calduch-Giner, J.; Pérez-Sánchez, J.; Dias, J.; Conceição, L.; Silva, T.; Costas, B. Dietary Histidine, Threonine, or Taurine Supplementation Affects Gilthead Seabream (Sparus aurata) Immune Status. Animals 2021, 11, 1193. [Google Scholar] [CrossRef]
- Torres, M.; Testa, C.P.; Gáspari, C.; Masutti, M.B.; Panitz, C.M.N.; Pedrosa, R.C.; de Almeida, E.A.; Di Mascio, P.; Filho, D.W. Oxidative stress in the mussel Mytella guyanensis from polluted mangroves on Santa Catarina Island, Brazil. Mar. Pollut. Bull. 2002, 44, 923–932. [Google Scholar] [CrossRef]
- Almeida, J.R.; Oliveira, C.; Gravato, C.; Guilhermino, L. Linking behavioural alterations with biomarkers responses in the European seabass Dicentrarchus labrax L. exposed to the organophosphate pesticide fenitrothion. Ecotoxicology 2010, 19, 1369–1381. [Google Scholar] [CrossRef]
- Bird, R.; Draper, H. Comparative studies on different methods of malonaldehyde determination. Methods Enzymol. 1984, 105, 299–305. [Google Scholar] [CrossRef]
- Peixoto, D.; Pinto, W.; Gonçalves, A.T.; Machado, M.; Reis, B.; Silva, J.; Navalho, J.; Dias, J.; Conceição, L.; Costas, B. Microalgal biomasses have potential as ingredients in microdiets for Senegalese sole (Solea senegalensis) post-larvae. J. Appl. Phycol. 2021, 33, 2241–2250. [Google Scholar] [CrossRef]
- Claiborne, A. Catalase activity. In Handbook of Methods for Oxygen Radical Research; Greenwald, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 283–284. [Google Scholar]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Baker, M.A.; Cerniglia, G.J.; Zaman, A. Microtiter plate assay for the measurement of glutathione and glutathione disulfide in large numbers of biological samples. Anal. Biochem. 1990, 190, 360–365. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-Transferases. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Frasco, M.; Guilhermino, L. Effects of dimethoate and beta-naphthoflavone on selected biomarkers of Poecilia reticulata. Fish Physiol. Biochem. 2002, 26, 149–156. [Google Scholar] [CrossRef]
- Flohé, L.; Ötting, F. Superoxide dismutase assays. Methods Enzymol. 1984, 105, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Lima, I.; Moreira, S.M.; Osten, J.R.-V.; Soares, A.M.; Guilhermino, L. Biochemical responses of the marine mussel Mytilus galloprovincialis to petrochemical environmental contamination along the North-western coast of Portugal. Chemosphere 2007, 66, 1230–1242. [Google Scholar] [CrossRef]
- Machado, M.; Azeredo, R.; Fontinha, F.; Fernández-Boo, S.; Conceicao, L.; Dias, J.; Costas, B. Dietary Methionine Improves the European Seabass (Dicentrarchus labrax) Immune Status, Inflammatory Response, and Disease Resistance. Front. Immunol. 2018, 9, 2672. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.; Du, B.; Xu, B. A critical review on production and industrial applications of beta-glucans. Food Hydrocoll. 2016, 52, 275–288. [Google Scholar] [CrossRef]
- Nordvi, M.F.; Løvmo, S.D.; Whatmore, P.; Sundh, H.; Sigholt, T.; Olsen, R.E. Low intestinal inflammation model (HP48) in Atlantic salmon (Salmo salar) and inflammatory mitigation by Bactocell. Aquaculture 2022, 563, 738920. [Google Scholar] [CrossRef]
- Burge, K.; Gunasekaran, A.; Eckert, J.; Chaaban, H. Curcumin and Intestinal Inflammatory Diseases: Molecular Mechanisms of Protection. Int. J. Mol. Sci. 2019, 20, 1912. [Google Scholar] [CrossRef] [Green Version]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A Review of Its Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef] [Green Version]
- Eichele, D.D.; Kharbanda, K.K. Dextran sodium sulfate colitis murine model: An indispensable tool for advancing our understanding of inflammatory bowel diseases pathogenesis. World J. Gastroenterol. 2017, 23, 6016–6029. [Google Scholar] [CrossRef]
- Rattigan, R.; O’Doherty, J.V.; Vigors, S.; Ryan, M.T.; Sebastiano, R.S.; Callanan, J.J.; Thornton, K.; Rajauria, G.; Margassery, L.M.; Dobson, A.D.W.; et al. The Effects of the Marine-Derived Polysaccharides Laminarin and Chitosan on Aspects of Colonic Health in Pigs Challenged with Dextran Sodium Sulphate. Mar. Drugs 2020, 18, 262. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lin, F.; Zhu, X.; Leone, V.A.; Dalal, S.; Tao, Y.; Messer, J.S.; Chang, E.B. Distinct roles of intracellular heat shock protein 70 in maintaining gastrointestinal homeostasis. Am. J. Physiol. Liver Physiol. 2018, 314, G164–G178. [Google Scholar] [CrossRef] [PubMed]
- Akdemir, F.; Orhan, C.; Tuzcu, M.; Sahin, N.; Juturu, V.; Sahin, K. The efficacy of dietary curcumin on growth performance, lipid peroxidation and hepatic transcription factors in rainbow trout Oncorhynchus Mykiss (Walbaum) reared under different stocking densities. Aquac. Res. 2017, 48, 4012–4021. [Google Scholar] [CrossRef]
- Xia, C.; Cai, Y.; Li, S.; Yang, J.; Xiao, G. Curcumin Increases HSP70 Expression in Primary Rat Cortical Neuronal Apoptosis Induced by gp120 V3 Loop Peptide. Neurochem. Res. 2015, 40, 1996–2005. [Google Scholar] [CrossRef]
- Cao, L.; Ding, W.; Du, J.; Jia, R.; Liu, Y.; Zhao, C.; Shen, Y.; Yin, G. Effects of curcumin on antioxidative activities and cytokine production in Jian carp (Cyprinus carpio var. Jian) with CCl4-induced liver damage. Fish Shellfish Immunol. 2015, 43, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Giri, S.S.; Sukumaran, V.; Park, S.C. Effects of bioactive substance from turmeric on growth, skin mucosal immunity and antioxidant factors in common carp, Cyprinus carpio. Fish Shellfish Immunol. 2019, 92, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Rajabiesterabadi, H.; Hoseini, S.M.; Fazelan, Z.; Hoseinifar, S.H.; Van Doan, H. Effects of dietary turmeric administration on stress, immune, antioxidant and inflammatory responses of common carp (Cyprinus carpio) during copper exposure. Aquac. Nutr. 2020, 26, 1143–1153. [Google Scholar] [CrossRef]
- Yonar, S.M.; Yonar, M.E.; Ural, M. Antioxidant effect of curcumin against exposure to malathion in Cyprinus carpio. Cell. Mol. Biol. 2017, 63, 68–72. [Google Scholar] [CrossRef]
- Manju, M.; Sherin, T.G.; Rajasekharan, K.N.; Oommen, O.V. Curcumin analogue inhibits lipid peroxidation in a freshwater teleost, Anabas testudineus (Bloch)—An in vitro and in vivo study. Fish Physiol. Biochem. 2009, 35, 413–420. [Google Scholar] [CrossRef]
- Manju, M.; Akbarsha, M.A.; Oommen, O.V. In vivo protective effect of dietary curcumin in fish Anabas testudineus (Bloch). Fish Physiol. Biochem. 2012, 38, 309–318. [Google Scholar] [CrossRef]
- Yonar, M.E. Chlorpyrifos-induced biochemical changes in Cyprinus carpio: Ameliorative effect of curcumin. Ecotoxicol. Environ. Saf. 2018, 151, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Xavier, M.; Dardengo, G.; Navarro-Guillén, C.; Lopes, A.; Colen, R.; Valente, L.; Conceição, L.; Engrola, S. Dietary Curcumin Promotes Gilthead Seabream Larvae Digestive Capacity and Modulates Oxidative Status. Animals 2021, 11, 1667. [Google Scholar] [CrossRef]
- Fazio, F. Fish hematology analysis as an important tool of aquaculture: A review. Aquaculture 2019, 500, 237–242. [Google Scholar] [CrossRef]
- Seibel, H.; Baßmann, B.; Rebl, A. Blood Will Tell: What Hematological Analyses Can Reveal About Fish Welfare. Front. Vet. Sci. 2021, 8, 616955. [Google Scholar] [CrossRef] [PubMed]
- Sugiharto, S.; Isroli, I.; Widiastuti, E.; Prabowo, N. Effect of turmeric extract on blood parameters, feed efficiency and abdominal fat content in broilers. J. Indones. Trop. Anim. Agric. 2011, 36, 21–26. [Google Scholar] [CrossRef] [Green Version]
- El-Feki, M.; Fahim, H.; Gad, M.; Abaci-Ghana, F. Changes in the haematological parameters of carp, Cyprinus carpjo L., induced by laminarin and Nigella sativa oil during the motile Aeromonas Septicemia Disease. Egypt. J. Aquat. Biol. Fish. 2000, 4, 43–93. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (% Feed) * | Control Diet | β-Glucans Diet | Curcumin Diet |
|---|---|---|---|
| Fishmeal super prime | 13.00 | 13.00 | 13.00 |
| Fishmeal 60 | 7.00 | 7.00 | 7.00 |
| CPSP 90 | 2.00 | 2.00 | 2.00 |
| Poultry meal 65 | 15.00 | 15.00 | 15.00 |
| Soy protein concentrate | 5.00 | 5.00 | 5.00 |
| Wheat gluten | 4.20 | 4.20 | 4.20 |
| Corn gluten meal | 8.00 | 8.00 | 8.00 |
| Soybean meal 48 | 10.00 | 10.00 | 10.00 |
| Rapeseed meal | 5.00 | 5.00 | 5.00 |
| Sunflower meal | 5.00 | 5.00 | 5.00 |
| Wheat meal | 11.13 | 10.13 | 10.13 |
| Vitamin and mineral premix | 1.00 | 1.00 | 1.00 |
| Vitamin C35 | 0.05 | 0.05 | 0.05 |
| Vitamin E50 | 0.02 | 0.02 | 0.02 |
| Betaine HCl | 0.20 | 0.20 | 0.20 |
| Antioxidant | 0.20 | 0.20 | 0.20 |
| Fish oil | 3.96 | 3.96 | 3.96 |
| Soybean oil | 5.28 | 5.28 | 5.28 |
| Rapeseed oil | 3.96 | 3.96 | 3.96 |
| Curcumin extract 95% | 0.20 | ||
| Algae beta-glucans extract | 1.00 | ||
| Proximate Analyses (% dry matter) | |||
| Moisture | 7.07 | ||
| Crude protein | 46.16 | ||
| Crude fat | 17.80 | ||
| Ash | 8.34 | ||
| Gross energy (MJ Kg−1) | 21.22 | ||
| Target Gene Name | Symbol | Accession nr. | Forward Primer (5′-3′) Reverse Primer (5′-3′) | PL 1 | MT 2 | E 3 |
|---|---|---|---|---|---|---|
| Mn superoxide dismutase | sod | JQ308833 | CCTGACCTGACCTACGACTATGG AGTGCCTCCTGATATTTCTCCTCTG | 134 | 60 | 2.05 |
| Glutathione peroxidase 1 | gpx | DQ524992 | GAAGGTGGATGTGAATGGAAAAGATG CTGACGGGACTCCAAATGATGG | 129 | 60 | 2.21 |
| Tumor necrosis factor alpha | tnfα | AJ413189.2 | TGAACAGAGGCGACAAACTG GCCACAAGCGTTATCTCCAT | 245 | 60 | 1.92 |
| Interleukin-10-like | il10 | XM_30418889.1 | AACATCCTGGGCTTCTATCTG GTGTCCTCCGTCTCATCTG | 65 | 57 | 2.07 |
| Immunoglobulin M heavy chain | igm | AM493677 | CAGCCTCGAGAAGTGGAAAC GAGGTTGACCAGGTTGGTGT | 136 | 59 | 2.19 |
| Macrophage colony stimulating factor receptor | csf1r | AM050293 | ACGTCTGGTCCTATGGCATC AGTCTGGTTGGGACATCTGG | 129 | 60 | 2.11 |
| CD8 alpha chain precursor | cd8α | AJ878605 | CTCGACTGGTCGGAGTTAA TCCATCAGCGGCTGCTCGT | 287 | 60 | 1.91 |
| Heat shock protein 70 | hsp70 | DQ524995.1 | ACGGCATCTTTGAGGTGAAG TGGCTGATGTCCTTCTTGTG | 124 | 55 | 2.05 |
| Occludin | ocln | KF861990.1 | TCATCTCCTACCCTCCAGTCA ATGGTCTGCTTGTGGTCCTC | 96 | 60 | 2.03 |
| Hepcidin 1 | hep | EF625901 | GCCATCGTGCTCACCTTTAT CCTGCTGCCATACCCCATCTT | 382 | 60 | 2.07 |
| Mucin-13-like | muc13 | XM030399162 | TTCAAACCCGTGTGGTCCAG GCACAAGCAGACATAGTTCGGATAT | 67 | 60 | 1.96 |
| Mucin-2-like | muc2 | XM_030425504.1 | GTGTGTGGCTGTGTTCCTTGCTTTGT GCGAACCAGTCTGGCTTGGACATCA | 67 | 60 | 1.98 |
| Parameters * | Feeding Trial | Inflammatory Trial | |||||
|---|---|---|---|---|---|---|---|
| CTRL | BG | CUR | CTRL | CTRL-D | BG-D | CUR-D | |
| HT (%) | 37.4 ± 6.7 | 36.4 ± 5.7 | 37.2 ± 6.9 | 37.0 b ± 6.4 | 39.7 ab ± 3.8 | 45.3 a ± 3.6 | 43.7 a ± 5.2 |
| RBC (×106 mL) | 1.4 ± 0.5 | 1.3 ± 0.4 | 1.2 ± 0.6 | 2.0 b ± 0.9 | 2.9 a ± 0.8 | 2.3 ab ± 0.7 | 2.7 ab ± 0.1 |
| HG (g dL−1) | 2.4 ± 1.0 | 2.6 ± 1.2 | 2.0 ± 0.7 | 2.0 ab ± 0.3 | 1.8 b ± 0.6 | 2.3 ab ± 0.8 | 2.9 a ± 1.1 |
| MCV (µm3) | 273.4 ± 72.8 | 310.1 ± 100.9 | 362.7 ± 209.6 | 209.7 a ± 70.9 | 134.7 b ± 27.1 | 190.1 a ± 48.3 | 162.1 ab ± 30.8 |
| MCH (pg cell−1) | 15.0 ± 3.2 | 16.1 ± 5.2 | 13.2 ± 3.9 | 11.5 ab ± 4.8 | 7.0 b ± 2.8 | 12.8 a ± 4.3 | 10.5 ab ± 3.2 |
| MCHC (g 100 mL−1) | 6.6 ± 2.7 | 7.6 ± 4.0 | 4.8 ± 0.5 | 5.4 ± 1.4 | 4.9 ± 1.9 | 6.3 ± 1.1 | 6.6 ± 2.0 |
| WBC (×104 mL) | 1.9 ± 0.9 | 2.0 ± 0.7 | 1.9 ± 0.7 | 1.7 ± 0.9 | 2.9 ± 1.7 | 3.0 ± 1.4 | 2.1 ± 1.3 |
| Thrombocytes (×104 mL) | 0.9 ± 0.4 | 1.2 ± 0.5 | 1.0 ± 0.4 | 0.7 b ± 0.3 | 1.6 a ± 1.0 | 1.6 a ± 0.8 | 1.2 ab ± 0.7 |
| Lymphocytes (×104 mL) | 0.6 ± 0.2 | 0.6 ± 0.2 | 0.6 ± 0.2 | 0.5 b ± 0.3 | 0.7 ab ± 0.3 | 1.0 a ± 0.5 | 0.6 ab ± 0.4 |
| Monocytes (×104 mL) | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.0 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.0 |
| Neutrophils (×104 mL) | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.2 | 0.1 ± 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira, C.; Peixoto, D.; Hinzmann, M.; Santos, P.; Ferreira, I.; Pereira, G.V.; Dias, J.; Costas, B. Dietary Strategies to Modulate the Health Condition and Immune Responses in Gilthead Seabream (Sparus aurata) Juveniles Following Intestinal Inflammation. Animals 2022, 12, 3019. https://doi.org/10.3390/ani12213019
Teixeira C, Peixoto D, Hinzmann M, Santos P, Ferreira I, Pereira GV, Dias J, Costas B. Dietary Strategies to Modulate the Health Condition and Immune Responses in Gilthead Seabream (Sparus aurata) Juveniles Following Intestinal Inflammation. Animals. 2022; 12(21):3019. https://doi.org/10.3390/ani12213019
Chicago/Turabian StyleTeixeira, Carla, Diogo Peixoto, Mariana Hinzmann, Paulo Santos, Inês Ferreira, Gabriella V. Pereira, Jorge Dias, and Benjamín Costas. 2022. "Dietary Strategies to Modulate the Health Condition and Immune Responses in Gilthead Seabream (Sparus aurata) Juveniles Following Intestinal Inflammation" Animals 12, no. 21: 3019. https://doi.org/10.3390/ani12213019