
Evaluating Self-Directed Behaviours and Their Association with Emotional Arousal across Two Cognitive Tasks in Bonobos (Pan paniscus)
 , , ,
, , ,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Housing
2.2. Testing Procedure
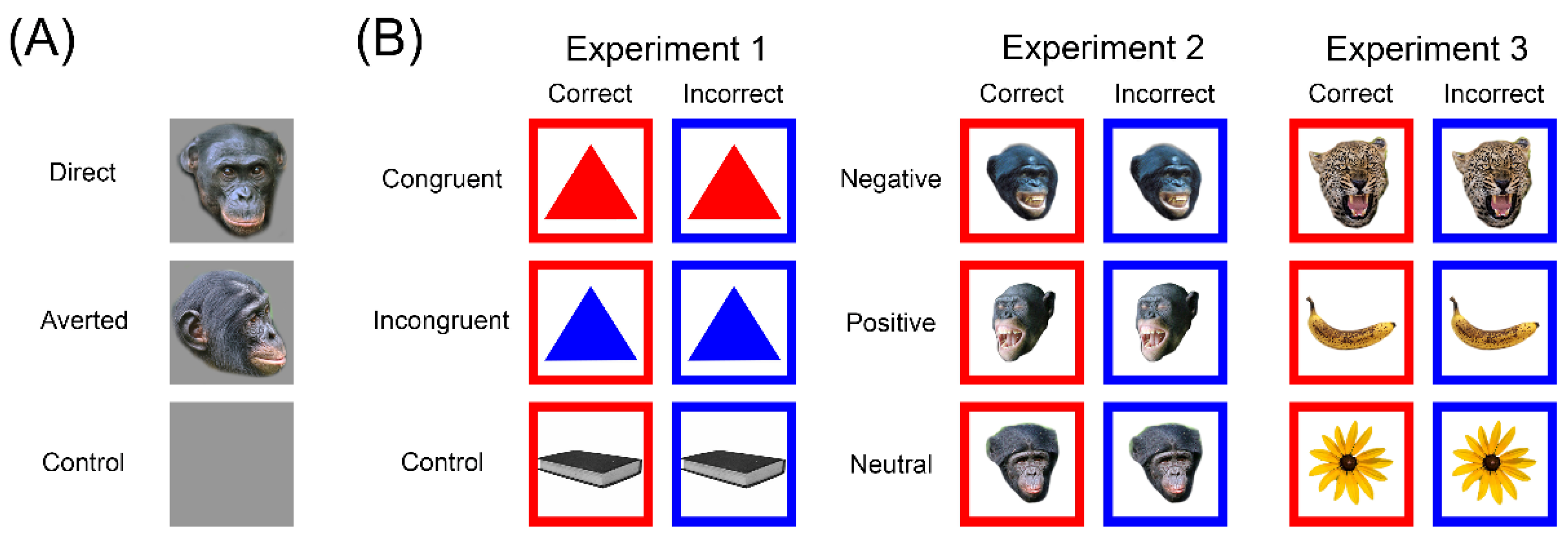
2.3. Touchscreen Tasks
2.4. Video Coding
2.5. Tube Task
2.6. Data Analyses
2.6.1. Handedness and Side Index
2.6.2. Linear Mixed Models
2.6.3. Behavioural Reactivity
3. Results
3.1. Production of SDB Types Per Study
3.2. Accuracy in the Two Studies
3.3. Linear Mixed Models
3.3.1. Study 1
3.3.2. Study 2
3.4. Handedness and Behavioural Reactivity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tinbergen, N. “Derived” activities; their causation, biological significance, origin, and emancipation during evolution. Q. Rev. Biol. 1952, 27, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Pavani, S.; Maestripieri, D.; Schino, G.; Turillazzi, P.G.; Scucchi, S. Factors Influencing Scratching Behaviour in Long-Tailed Macaques (Macaca fascicularis). Folia Primatol. 1991, 57, 34–38. [Google Scholar] [CrossRef]
- Maestripieri, D.; Schino, G.; Aureli, F.; Troisi, A. A modest proposal: Displacement activities as an indicator of emotions in primates. Anim. Behav. 1992, 44, 967–979. [Google Scholar] [CrossRef]
- Schino, G.; Troisi, A.; Perretta, G.; Monaco, V. Measuring anxiety in nonhuman primates: Effect of lorazepam on macaque scratching. Pharmacol. Biochem. Behav. 1991, 38, 889–891. [Google Scholar] [CrossRef]
- Schino, G.; National, I.; Perretta, G.; National, I.; National, I.; Troisi, A. Primate displacement activities as an ethopharmacological model of anxiety. Anxiety 1996, 2, 186–191. [Google Scholar] [CrossRef]
- Baker, K.C.; Aureli, F. Behavioural indicators of anxiety: An empirical test in chimpanzees. Behaviour 1997, 134, 1031–1050. [Google Scholar] [CrossRef]
- Castles, D.L.; Whiten, A. Post-conflict Behaviour of Wild Olive Baboons. II. Stress and Self-directed Behaviour. Ethology 1998, 104, 148–160. [Google Scholar] [CrossRef]
- Troisi, A. Displacement activities as a behavioral measure of stress in nonhuman primates and human subjects. Stress 2002, 5, 47–54. [Google Scholar] [CrossRef]
- Diezinger, F.; Anderson, J.R. Starting from scratch: A first look at a “displacement activity” in group-living rhesus monkeys. Am. J. Primatol. 1986, 11, 117–124. [Google Scholar] [CrossRef]
- Aureli, F.; Yates, K. Distress prevention by grooming others in crested black macaques. Biol. Lett. 2010, 6, 27–29. [Google Scholar] [CrossRef]
- Norscia, I.; Palagi, E. When play is a family business: Adult play, hierarchy, and possible stress reduction in common marmosets. Primates 2011, 52, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Aureli, F.; Van Schaik, C.P.; Van Hooff, J.A.R.A.M. Functional aspects of reconciliation among captive long-tailed macaques (Macaca fascicularis). Am. J. Primatol. 1989, 19, 39–51. [Google Scholar] [CrossRef]
- Fraser, O.N.; Stahl, D.; Aureli, F. Stress reduction through consolation in chimpanzees. Proc. Natl. Acad. Sci. USA 2008, 105, 8557–8562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leavens, D.A.; Aureli, F.; Hopkins, W.D.; Hyatt, C.W. Effects of cognitive challenge on self-directed behaviors by chimpanzees (Pan troglodytes). Am. J. Primatol. 2001, 55, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elder, C.M.; Menzel, C.R. Dissociation of cortisol and behavioral indicators of stress in an orangutan (Pongo pygmaeus) during a computerized task. Primates 2001, 42, 345–357. [Google Scholar] [CrossRef]
- Itakura, S. Emotional behavior during the learning of a contingency task in a chimpanzee. Percept. Mot. Ski. 1993, 76, 563–566. [Google Scholar] [CrossRef]
- Yamanashi, Y.; Matsuzawa, T. Emotional consequences when chimpanzees (Pan troglodytes) face challenges: Individual differences in self-directed behaviours during cognitive tasks. Anim. Welf. 2010, 19, 25–30. [Google Scholar]
- Leeds, A.; Lukas, K.E. Experimentally evaluating the function of self-directed behaviour in two adult mandrills (Mandrillus sphinx). Anim. Welf. 2018, 27, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Pomerantz, O.; Terkel, J. Effects of positive reinforcement training techniques on the psychological welfare of zoo-housed chimpanzees (Pan troglodytes). Am. J. Primatol. 2009, 71, 687–695. [Google Scholar] [CrossRef]
- Aureli, F.; de Waal, F.B.M. Inhibition of social behavior in chimpanzees under high-density conditions. Am. J. Primatol. 1997, 41, 213–228. [Google Scholar] [CrossRef]
- Leavens, D.A.; Aureli, F.; Hopkins, W.D. Behavioral evidence for the cutaneous expression of emotion in a chimpanzee (Pan troglodytes). Behaviour 2004, 141, 979–997. [Google Scholar] [CrossRef]
- Marchant, L.F.; McGrew, W.C. Laterality of limb function in wild chimpanzees of Gombe National Park: Comprehensive study of spontaneous activities. J. Hum. Evol. 1996, 30, 427–443. [Google Scholar] [CrossRef]
- Jordan, C. Das Verhalten Zoolebender Zwergschimpansen; Goethe University: Frankfurt am Main, Germany, 1977. [Google Scholar]
- Hopkins, W.D.; Russell, J.L.; Freeman, H.D.; Reynolds, E.A.M.; Griffis, C.; Leavens, D.A. Lateralized Scratching in Chimpanzees (Pan troglodytes): Evidence of a Functional Assymetry During Arousal. Emotion 2006, 6, 553–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, F.E.; Smith, L.J. Effect of a cognitive challenge device containing food and non-food rewards on chimpanzee well-being. Am. J. Primatol. 2013, 75, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Wagner, K.E.; Hopper, L.M.; Ross, S.R. Asymmetries in the production of self-directed behavior by chimpanzees and gorillas during a computerized cognitive test. Anim. Cogn. 2016, 19, 343–350. [Google Scholar] [CrossRef]
- Rogers, L.J.; Andrew, R.J. Comparative Vertebrate Lateralization; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Güntürkün, O.; Ströckens, F.; Ocklenburg, S. Brain lateralization: A comparative perspective. Physiol. Rev. 2020, 100, 1019–1063. [Google Scholar] [CrossRef]
- Rogers, L.J. Lateralization in Vertebrates: Its Early Evolution, General Pattern, and Development. Adv. Study Behav. 2002, 31, 107–161. [Google Scholar] [CrossRef]
- Rogers, L.J. Relevance of brain and behavioural lateralization to animal welfare. Appl. Anim. Behav. Sci. 2010, 127, 8. [Google Scholar] [CrossRef]
- Dimond, S.; Harries, R. Face touching in monkeys, apes and man: Evolutionary origins and cerebral asymmetry. Neuropsychologia 1984, 22, 227–233. [Google Scholar] [CrossRef]
- Hopkins, W.D.; de Waal, F.B.M. Behavioral laterality in captive bonobos (Pan paniscus): Replication and extension. Int. J. Primatol. 1995, 16, 261–276. [Google Scholar] [CrossRef]
- Neufuss, J.; Humle, T.; Cremaschi, A.; Kivell, T.L. Nut-cracking behaviour in wild-born, rehabilitated bonobos (Pan paniscus): A comprehensive study of hand-preference, hand grips and efficiency. Am. J. Primatol. 2017, 79, e22589. [Google Scholar] [CrossRef] [PubMed]
- Brand, C.M.; Marchant, L.F.; Boose, K.J.; White, F.J.; Rood, T.M.; Meinelt, A. Laterality of Grooming and Tool Use in a Group of Captive Bonobos (Pan paniscus). Folia Primatol. 2017, 88, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Chapelain, A.S.; Hogervorst, E.; Mbonzo, P.; Hopkins, W.D. Hand Preferences for Bimanual Coordination in 77 Bonobos (Pan paniscus): Replication and Extension. Int. J. Primatol. 2011, 32, 491–510. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Phillips, K.A.; Bania, A.; Calcutt, S.E.; Gardner, M.; Russell, J.; Schaeffer, J.; Lonsdorf, E.V.; Ross, S.R.; Schapiro, S.J. Hand preferences for coordinated bimanual actions in 777 great apes: Implications for the evolution of handedness in Hominins. J. Hum. Evol. 2011, 60, 605–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, W.D.; Schaeffer, J.; Russell, J.L.; Bogart, S.L.; Meguerditchian, A.; Coulon, O. A comparative assessment of handedness and its potential neuroanatomical correlates in chimpanzees (Pan troglodytes) and bonobos (Pan paniscus). Behaviour 2015, 152, 461–492. [Google Scholar] [CrossRef] [Green Version]
- Meguerditchian, A.; Phillips, K.A.; Chapelain, A.; Mahovetz, L.M.; Milne, S.; Stoinski, T.; Bania, A.; Lonsdorf, E.; Schaeffer, J.; Russell, J.; et al. Handedness for unimanual grasping in 564 great apes: The effect on grip morphology and a comparison with hand use for a bimanual coordinated task. Front. Psychol. 2015, 6, 1794. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, W.D.; Lyn, H.; Cantalupo, C. Volumetric and lateralized differences in selected brain regions of chimpanzees (Pan troglodytes) and bonobos (Pan paniscus). Am. J. Primatol. 2009, 71, 988–997. [Google Scholar] [CrossRef] [Green Version]
- Rosati, A.G.; Hare, B. Chimpanzees and Bonobos Exhibit Emotional Responses to Decision Outcomes. PLoS ONE 2013, 8, e63058. [Google Scholar] [CrossRef] [Green Version]
- Stimpson, C.D.; Barger, N.; Taglialatela, J.P.; Gendron-Fitzpatrick, A.; Hof, P.R.; Hopkins, W.D.; Sherwood, C.C. Differential serotonergic innervation of the amygdala in bonobos and chimpanzees. Soc. Cogn. Affect. Neurosci. 2016, 11, 413–422. [Google Scholar] [CrossRef] [Green Version]
- Issa, H.A.; Staes, N.; Diggs-Galligan, S.; Stimpson, C.D.; Gendron-Fitzpatrick, A.; Taglialatela, J.P.; Hof, P.R.; Hopkins, W.D.; Sherwood, C.C. Comparison of bonobo and chimpanzee brain microstructure reveals differences in socio-emotional circuits. Brain Struct. Funct. 2019, 224, 239–251. [Google Scholar] [CrossRef]
- Staes, N.; Smaers, J.B.; Kunkle, A.E.; Hopkins, W.D.; Bradley, B.J.; Sherwood, C.C. Evolutionary divergence of neuroanatomical organization and related genes in chimpanzees and bonobos. Cortex 2019, 118, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Mathôt, S.; Schreij, D.; Theeuwes, J. OpenSesame: An open-source, graphical experiment builder for the social sciences. Behav. Res. Methods 2012, 44, 314–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bethell, E.J.; Holmes, A.; MacLarnon, A.; Semple, S. Emotion Evaluation and Response Slowing in a Non-Human Primate: New Directions for Cognitive Bias Measures of Animal Emotion? Behav. Sci. 2016, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Allritz, M.; Call, J.; Borkenau, P. How chimpanzees (Pan troglodytes) perform in a modified emotional Stroop task. Anim. Cogn. 2016, 19, 435–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laméris, D.W.; Verspeek, J.; Eens, M.; Stevens, J.M.G. Social and nonsocial stimuli alter the performance of bonobos during a pictorial emotional Stroop task. Am. J. Primatol. 2022, 84, e23356. [Google Scholar] [CrossRef]
- Fagot, J.; Vauclair, J. Manual laterality in nonhuman primates: A distinction between handedness and manual specialization. Psychol. Bull. 1991, 109, 76–89. [Google Scholar] [CrossRef]
- Hopkins, W.D. Hand Preferences for a Coordinated Bimanual Task in 110 Chimpanzees (Pan troglodytes): Cross-Sectional Analysis. J. Comp. Psychol. 1995, 109, 291–297. [Google Scholar] [CrossRef]
- Staes, N.; Sherwood, C.C.; Freeman, H.; Brosnan, S.F.; Schapiro, S.J.; Hopkins, W.D.; Bradley, B.J. Serotonin receptor 1a variation is associated with anxiety and agonistic behavior in chimpanzees. Mol. Biol. Evol. 2019, 36, 1418–1429. [Google Scholar] [CrossRef]
- Kret, M.E.; Jaasma, L.; Bionda, T.; Wijnen, J.G. Bonobos (Pan paniscus) show an attentional bias toward conspecifics’ emotions. Proc. Natl. Acad. Sci. USA 2016, 113, 201522060. [Google Scholar] [CrossRef] [Green Version]
- Troisi, A.; Schino, G.; D’Antoni, M.; Pandolfi, N.; Aureli, F.; D’Amato, F.R. Scratching as a behavioral index of anxiety in macaque mothers. Behav. Neural Biol. 1991, 56, 307–313. [Google Scholar] [CrossRef]
- Braccini, S.N.; Caine, N.G. Hand Preference Predicts Reactions to Novel Foods and Predators in Marmosets (Callithrix geoffroyi). J. Comp. Psychol. 2009, 123, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.J.; Rogers, L.J. Cognitive bias, hand preference and welfare of common marmosets. Behav. Brain Res. 2015, 287, 100–108. [Google Scholar] [CrossRef]
- Lilak, A.L.; Phillips, K.A. Consistency of hand preference across low-level and high-level tasks in capuchin monkeys (Cebus apella). Am. J. Primatol. 2008, 70, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Westergaard, G.C.; Champoux, M.; Suomi, S.J. Plasma cortisol is associated with handedness in infant rhesus monkeys. Dev. Psychobiol. 2001, 38, 116–122. [Google Scholar] [CrossRef]
- Vaughan, E.; Le, A.; Casey, M.; Workman, K.P.; Lacreuse, A. Baseline cortisol levels and social behavior differ as a function of handedness in marmosets (Callithrix jacchus). Am. J. Primatol. 2019, 81, e23057. [Google Scholar] [CrossRef]
- Neal, S.J.; Caine, N.G. Scratching under positive and negative arousal in common marmosets (Callithrix jacchus). Am. J. Primatol. 2016, 78, 216–226. [Google Scholar] [CrossRef]
- ASAB. Guidelines for the treatment of animals in behavioural research and teaching. Anim. Behav. 2020, 159, I–XI. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Subject | Sex | Age | Study 1 | Study 2 |
|---|---|---|---|---|
| Busira | Female | 16 | Yes | Yes |
| Habari | Male | 14 | Yes | Yes |
| Kianga | Female | 17 | Yes | No |
| Kikongo | Male | 7 | Yes | No |
| Mokonzi | Male | 7 | Yes | Yes |
| Nayembi | Female | 15 | Yes | No |
| Vifijo | Male | 27 | Yes | No |
| Zamba | Male | 22 | Yes | Yes |
| Nose Wipe | Gentle Scratch | Rough Scratch | Self-Touch | |
|---|---|---|---|---|
| Study 1 (RST) | 69.2% | 19.8% | 9.1% | 1.9% |
| Study 2 (MEST) | 80.6% | 11.9% | 5.6% | 1.9% |
| Total (% of total) | 75.5% | 18.4% | 7.2% | 1.9% |
| Subject | HI-Tube | HI-Screen | HI-Gentle Scratch | HI-Rough Scratch | HI-Nose Wipe | SI-Gentle Scratch | SI-Rough Scratch |
|---|---|---|---|---|---|---|---|
| Busira | −0.01 | 0.92 | 0.79 | 0.77 | 0.98 | 0.28 | −0.15 |
| Habari | 0.61 | −0.47 | −0.33 | −0.69 | −0.35 | 0.25 | 0.43 |
| Kianga | −0.76 | 0.00 | −0.21 | −0.47 | 0.16 | −0.32 | 0.20 |
| Kikongo | 0.15 | 0.96 | −0.27 | NA | −0.54 | 0.45 | NA |
| Mokonzi | 0.25 | 0.52 | 0.03 | −0.50 | −0.03 | 0.10 | 0.5 |
| Nayembi | 1.00 | 0.72 | 0.33 | 0.40 | 1.00 | 0.33 | −0.20 |
| Vifijo | 0.72 | 0.97 | 1.00 | 0.33 | 0.78 | 0.00 | −0.33 |
| Zamba | 0.68 | −0.67 | 0.00 | 0.41 | −0.17 | −0.50 | −0.76 |
| Study 1 | Study 2 | ||||||
|---|---|---|---|---|---|---|---|
| Nose wipe | Chisq | df | p | Nose wipe | Chisq | df | p |
| Accuracy | 5.989 | 1 | 0.014 | Accuracy | 21.216 | 1 | <0.001 |
| Gentle Scratch | Chisq | df | p | Gentle Scratch | Chisq | df | p |
| Accuracy a | 0.277 | 1 | 0.599 | Accuracy a | 8.328 | 1 | 0.004 |
| Hemispace a | 0.558 | 1 | 0.455 | Hemispace a | 0.001 | 1 | 0.969 |
| Accuracy * Hemispace | 0.102 | 1 | 0.750 | Accuracy * Hemispace | 0.031 | 1 | 0.860 |
| Rough scratch | Chisq | df | p | Rough scratch | Chisq | df | p |
| Accuracy a | 0.816 | 1 | 0.366 | Accuracy | 15.604 | 1 | <0.001 |
| Hemispace a | 3.866 | 1 | 0.049 | Hemispace | 0.086 | 1 | 0.769 |
| Accuracy * Hemispace | 0.146 | 1 | 0.702 | Accuracy * Hemispace | 6.469 | 1 | 0.011 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laméris, D.W.; Verspeek, J.; Salas, M.; Staes, N.; Torfs, J.R.R.; Eens, M.; Stevens, J.M.G. Evaluating Self-Directed Behaviours and Their Association with Emotional Arousal across Two Cognitive Tasks in Bonobos (Pan paniscus). Animals 2022, 12, 3002. https://doi.org/10.3390/ani12213002
Laméris DW, Verspeek J, Salas M, Staes N, Torfs JRR, Eens M, Stevens JMG. Evaluating Self-Directed Behaviours and Their Association with Emotional Arousal across Two Cognitive Tasks in Bonobos (Pan paniscus). Animals. 2022; 12(21):3002. https://doi.org/10.3390/ani12213002
Chicago/Turabian StyleLaméris, Daan W., Jonas Verspeek, Marina Salas, Nicky Staes, Jonas R. R. Torfs, Marcel Eens, and Jeroen M. G. Stevens. 2022. "Evaluating Self-Directed Behaviours and Their Association with Emotional Arousal across Two Cognitive Tasks in Bonobos (Pan paniscus)" Animals 12, no. 21: 3002. https://doi.org/10.3390/ani12213002