
Formulating Diets for Improved Health Status of Pigs: Current Knowledge and Perspectives

Abstract
:Simple Summary
Abstract
1. Nutrition and Health: The Merge
2. Immune System Stimulation, Nutrient Efficiency and Requirements
3. Immune Challenges in Pork Production
3.1. Enteric Pathogen Challenge
3.2. Respiratory Pathogen Challenge
3.3. Degradation of Sanitary Condition
3.4. Bacterial Lipopolysaccharide
4. Nutritional Strategies for Improved Health Status
4.1. Low Protein, Amino Acid-Supplemented Diets
4.2. Dietary Fiber Level and Source
4.3. Diet Complexity
4.4. Functional Amino Acids
4.5. Organic Acids
4.6. Plant Secondary Metabolites
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- van der Meer, Y.; Lammers, A.; Jansman, A.J.M.; Rijnen, M.M.J.A.; Hendriks, W.H.; Gerrits, W.J.J. Performance of Pigs Kept under Different Sanitary Conditions Affected by Protein Intake and Amino Acid Supplementation. J. Anim. Sci. 2016, 94, 4704–4719. [Google Scholar] [CrossRef]
- van der Meer, Y.; Jansman, A.J.M.; Gerrits, W.J.J. Low Sanitary Conditions Increase Energy Expenditure for Maintenance and Decrease Incremental Protein Efficiency in Growing Pigs. Animal 2020, 14, 1811–1820. [Google Scholar] [CrossRef]
- Buchanan, J.B.; Johnson, R.W. Regulation of Food Intake by Inflammatory Cytokines in the Brain. Neuroendocrinology 2007, 86, 183–190. [Google Scholar] [CrossRef]
- Johnson, R.W. Immune and Endocrine Regulation of Food Intake in Sick Animals. Domest. Anim. Endocrinol. 1998, 15, 309–319. [Google Scholar] [CrossRef]
- Karrow, N.A. Activation of the Hypothalamic-Pituitary-Adrenal Axis and Autonomic Nervous System during Inflammation and Altered Programming of the Neuroendocrine-Immune Axis during Fetal and Neonatal Development: Lessons Learned from the Model Inflammagen, Lipopolysaccharide. Brain Behav. Immun. 2006, 20, 144–158. [Google Scholar] [CrossRef]
- Meier, U.; Gressner, A.M. Endocrine Regulation of Energy Metabolism: Review of Pathobiochemical and Clinical Chemical Aspects of Leptin, Ghrelin, Adiponectin, and Resistin. Clin. Chem. 2004, 50, 1511–1525. [Google Scholar] [CrossRef]
- Le Floc’h, N.; Melchior, D.; Obled, C. Modifications of Protein and Amino Acid Metabolism during Inflammation and Immune System Activation. Livest. Prod. Sci. 2004, 87, 37–45. [Google Scholar] [CrossRef]
- Pastorelli, H.; van Milgen, J.; Lovatto, P.; Montagne, L. Meta-Analysis of Feed Intake and Growth Responses of Growing Pigs after a Sanitary Challenge. Animal 2012, 6, 952–961. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, L.A.; Ferreira, F.N.A.; Costa, M.O.; Wellington, M.O.; Columbus, D.A. Factors Affecting Performance Response of Pigs Exposed to Different Challenge Models: A Multivariate Approach. J. Anim. Sci. 2021, 99, skab035. [Google Scholar] [CrossRef]
- Rodrigues, L.A.; Wellington, M.O.; González-Vega, J.C.; Htoo, J.K.; Van Kessel, A.G.; Columbus, D.A. Functional Amino Acid Supplementation, Regardless of Dietary Protein Content, Improves Growth Performance and Immune Status of Weaned Pigs Challenged with Salmonella Typhimurium. J. Anim. Sci. 2021, 99, skaa365. [Google Scholar] [CrossRef]
- Rodrigues, L.A.; Wellington, M.O.; González-Vega, J.C.; Htoo, J.K.; Van Kessel, A.G.; Columbus, D.A. A Longer Adaptation Period to a Functional Amino Acid-Supplemented Diet Improves Growth Performance and Immune Status of Salmonella Typhimurium-Challenged Pigs. J. Anim. Sci. 2021, 99, skab146. [Google Scholar] [CrossRef]
- Schweer, W.; Schwartz, K.; Patience, J.F.; Karriker, L.; Sparks, C.; Weaver, M.; Fitzsimmons, M.; Burkey, T.E.; Gabler, N.K. Porcine Reproductive and Respiratory Syndrome Virus Reduces Feed Efficiency, Digestibility, and Lean Tissue Accretion in Grow-Finish Pigs. Trans. Anim. Sci. 2017, 1, 480–488. [Google Scholar] [CrossRef] [Green Version]
- Dee, S.; Guzman, J.E.; Hanson, D.; Garbes, N.; Morrison, R.; Amodie, D.; Galina Pantoja, L. A Randomized Controlled Trial to Evaluate Performance of Pigs Raised in Antibiotic-Free or Conventional Production Systems Following Challenge with Porcine Reproductive and Respiratory Syndrome Virus. PLoS ONE 2018, 13, e0208430. [Google Scholar] [CrossRef] [Green Version]
- Khafipour, E.; Munyaka, P.M.; Nyachoti, C.M.; Krause, D.O.; Rodriguez-Lecompte, J.C. Effect of Crowding Stress and Escherichia coli K88+ Challenge in Nursery Pigs Supplemented with Anti-Escherichia coli K88+ Probiotics. J. Anim. Sci. 2014, 92, 2017–2029. [Google Scholar] [CrossRef]
- Jayaraman, B.; Htoo, J.; Nyachoti, C.M. Effects of Dietary Threonine:Lysine Ratioes and Sanitary Conditions on Performance, Plasma Urea Nitrogen, Plasma-Free Threonine and Lysine of Weaned Pigs. Anim. Nutr. 2015, 1, 283–288. [Google Scholar] [CrossRef]
- Laskoski, F. Effects of Pigs per Feeder Hole and Group Size on Feed Intake Onset, Growth Performance, and Ear and Tail Lesions in Nursery Pigs with Consistent Space Allowance. J. Swine Health Prod. 2019, 27, 12–18. [Google Scholar]
- Rauw, W.M.; de Mercado de la Peña, E.; Gomez-Raya, L.; García Cortés, L.A.; Ciruelos, J.J.; Gómez Izquierdo, E. Impact of Environmental Temperature on Production Traits in Pigs. Sci. Rep. 2020, 10, 2106. [Google Scholar] [CrossRef] [Green Version]
- Wellington, M.O.; Htoo, J.K.; Van Kessel, A.G.; Columbus, D.A. Impact of Dietary Fiber and Immune System Stimulation on Threonine Requirement for Protein Deposition in Growing Pigs. J. Anim. Sci. 2018, 96, 5222–5232. [Google Scholar] [CrossRef]
- Dong, G.Z.; Pluske, J.R. The Low Feed Intake in Newly-Weaned Pigs: Problems and Possible Solutions. Asian-Austral. J. Anim. Sci. 2007, 20, 440–452. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Duarte, M.E.; Sevarolli Loftus, A.; Kim, S.W. Intestinal Health of Pigs Upon Weaning: Challenges and Nutritional Intervention. Front. Vet. Sci. 2021, 8, 628258. [Google Scholar] [CrossRef]
- Blecha, F.; Pollman, D.S.; Nichols, D.A. Weaning Pigs at an Early Age Decreases Cellular Immunity. J. Anim. Sci. 1983, 56, 396–400. [Google Scholar] [CrossRef]
- Rooke, J.A.; Bland, I.M. The Acquisition of Passive Immunity in the New-Born Piglet. Livest. Prod. Sci. 2002, 78, 13–23. [Google Scholar] [CrossRef]
- Yuan, Y.; Jansen, J.; Charles, D.; Zanella, A.J. The Influence of Weaning Age on Post-Mixing Agonistic Interactions in Growing Pigs. Appl. Anim. Behav. Sci. 2004, 88, 39–46. [Google Scholar] [CrossRef]
- Spreeuwenberg, M.A.; Verdonk, J.M.; Gaskins, H.R.; Verstegen, M.W. Small Intestine Epithelial Barrier Function Is Compromised in Pigs with Low Feed Intake at Weaning. J. Nutr. 2001, 131, 1520–1527. [Google Scholar] [CrossRef] [Green Version]
- Pluske, J.R.; Kim, J.C.; Black, J.L.; Pluske, J.R.; Kim, J.C.; Black, J.L. Manipulating the Immune System for Pigs to Optimise Performance. Anim. Prod. Sci. 2018, 58, 666–680. [Google Scholar] [CrossRef]
- Marquardt, R.R.; Li, S. Antimicrobial Resistance in Livestock: Advances and Alternatives to Antibiotics. Anim. Front. 2018, 8, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Parra, M.D.; Fuentes, P.; Tecles, F.; Martínez-Subiela, S.; Martínez, J.S.; Muñoz, A.; Cerón, J.J. Porcine Acute Phase Protein Concentrations in Different Diseases in Field Conditions. J. Vet. Med. B 2006, 53, 488–493. [Google Scholar] [CrossRef]
- Reeds, P.J.; Jahoor, F. The Amino Acid Requirements of Disease. Clin. Nutr. 2001, 20, 15–22. [Google Scholar] [CrossRef]
- Obled, C. Amino Acid Requirements in Inflammatory States. Can. J. Anim. Sci. 2003, 83, 365–373. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative Stress: An Essential Factor in the Pathogenesis of Gastrointestinal Mucosal Diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [Green Version]
- Rakhshandeh, A.; de Lange, C.F.M.; Htoo, J.K.; Gheisari, A.; Rakhshandeh, A.R. Immune System Stimulation Increases the Plasma Cysteine Flux and Whole-Body Glutathione Synthesis Rate in Starter Pigs. J. Anim. Sci. 2019, 97, 3871–3881. [Google Scholar] [CrossRef]
- Le Floc’h, N.; Wessels, A.; Corrent, E.; Wu, G.; Bosi, P. The Relevance of Functional Amino Acids to Support the Health of Growing Pigs. Anim. Feed Sci. Technol. 2018, 245, 104–116. [Google Scholar] [CrossRef]
- Luppi, A. Swine Enteric Colibacillosis: Diagnosis, Therapy and Antimicrobial Resistance. Porc. Health Manag. 2017, 3, 16. [Google Scholar] [CrossRef]
- Luzzi, I.; Galetta, P.; Massari, M.; Rizzo, C.; Dionisi, A.M.; Filetici, E.; Cawthorne, A.; Tozzi, A.; Argentieri, M.; Bilei, S.; et al. An Easter Outbreak of Salmonella Typhimurium DT 104A Associated with Traditional Pork Salami in Italy. Eurosurveillance 2007, 12, 11–12. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, K.G.; Sørensen, G.; Torpdahl, M.; Kjeldsen, M.K.; Jensen, T.; Gubbels, S.; Bjerager, G.O.; Wingstrand, A.; Porsbo, L.J.; Ethelberg, S. A Long-Lasting Outbreak of Salmonella Typhimurium U323 Associated with Several Pork Products, Denmark, 2010. Epidemiol. Infect. 2013, 141, 260–268. [Google Scholar] [CrossRef]
- Clements, A.; Young, J.C.; Constantinou, N.; Frankel, G. Infection Strategies of Enteric Pathogenic Escherichia coli. Gut Microbes 2012, 3, 71–87. [Google Scholar] [CrossRef] [Green Version]
- Fàbrega, A.; Vila, J. Salmonella enterica Serovar Typhimurium Skills To Succeed in the Host: Virulence and Regulation. Clin. Microb. Rev. 2013, 26, 308–341. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Jayaratne, R.; Barrett, K.E. The Role of Ion Transporters in the Pathophysiology of Infectious Diarrhea. Cell Mol. Gastroenterol. Hepatol. 2018, 6, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Vereecke, L.; Beyaert, R.; van Loo, G. Enterocyte Death and Intestinal Barrier Maintenance in Homeostasis and Disease. Trends Mol. Med. 2011, 17, 584–593. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Intestinal Redox Biology and Oxidative Stress. Semin. Cell Dev. Biol. 2012, 23, 729–737. [Google Scholar] [CrossRef] [Green Version]
- Parker, A.; Vaux, L.; Patterson, A.M.; Modasia, A.; Muraro, D.; Fletcher, A.G.; Byrne, H.M.; Maini, P.K.; Watson, A.J.M.; Pin, C. Elevated Apoptosis Impairs Epithelial Cell Turnover and Shortens Villi in TNF-Driven Intestinal Inflammation. Cell Death Dis. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, R.; Boivin, M.; Ma, T. Mechanism of Cytokine Modulation of Epithelial Tight Junction Barrier. Front. Biosci 2009, 14, 2765–2778. [Google Scholar] [CrossRef] [Green Version]
- Pan, L.; Zhao, P.F.; Ma, X.K.; Shang, Q.H.; Xu, Y.T.; Long, S.F.; Wu, Y.; Yuan, F.M.; Piao, X.S. Probiotic Supplementation Protects Weaned Pigs against Enterotoxigenic Escherichia coli K88 Challenge and Improves Performance Similar to Antibiotics. J. Anim. Sci. 2017, 95, 2627–2639. [Google Scholar] [CrossRef]
- Choi, J.; Wang, L.; Liu, S.; Lu, P.; Zhao, X.; Liu, H.; Lahaye, L.; Santin, E.; Liu, S.; Nyachoti, M.; et al. Effects of a Microencapsulated Formula of Organic Acids and Essential Oils on Nutrient Absorption, Immunity, Gut Barrier Function, and Abundance of Enterotoxigenic Escherichia coli F4 in Weaned Piglets Challenged with E. coli F4. J. Anim. Sci. 2020, 98, skaa259. [Google Scholar] [CrossRef]
- Koo, B.; Bustamante-García, D.; Kim, J.W.; Nyachoti, C.M. Health-Promoting Effects of Lactobacillus-Fermented Barley in Weaned Pigs Challenged with Escherichia coli K88+. Animal 2020, 14, 39–49. [Google Scholar] [CrossRef]
- Heo, J.-M. Reducing the Protein Content in Diets for Weaner Pigs to Control Post-Weaning Diarrhoea: Physiological and Metabolic Responses of the Gastrointestinal Tract. Ph.D. Thesis, Murdoch University, Perth, Australia, 2010. [Google Scholar]
- Ren, M.; Liu, X.T.; Wang, X.; Zhang, G.J.; Qiao, S.Y.; Zeng, X.F. Increased Levels of Standardized Ileal Digestible Threonine Attenuate Intestinal Damage and Immune Responses in Escherichia coli K88+ Challenged Weaned Piglets. Anim. Feed Sci. Technol. 2014, 195, 67–75. [Google Scholar] [CrossRef]
- Capozzalo, M.M.; Resink, J.W.; Htoo, J.K.; Kim, J.C.; de Lange, F.M.; Mullan, B.P.; Hansen, C.F.; Pluske, J.R. Determination of the Optimum Standardised Ileal Digestible Sulphur Amino Acids to Lysine Ratio in Weaned Pigs Challenged with Enterotoxigenic Escherichia coli. Anim. Feed Sci. Technol. 2017, 227, 118–130. [Google Scholar] [CrossRef] [Green Version]
- Wellington, M.O.; Agyekum, A.K.; Hamonic, K.; Htoo, J.K.; Van Kessel, A.G.; Columbus, D.A. Effect of Supplemental Threonine above Requirement on Growth Performance of Salmonella Typhimurium Challenged Pigs Fed High-Fiber Diets. J. Anim. Sci. 2019, 97, 3636–3647. [Google Scholar] [CrossRef]
- Koo, B.; Choi, J.; Yang, C.; Nyachoti, C.M. Diet Complexity and L-Threonine Supplementation: Effects on Growth Performance, Immune Response, Intestinal Barrier Function, and Microbial Metabolites in Nursery Pigs. J. Anim. Sci. 2020, 98, skaa125. [Google Scholar] [CrossRef] [PubMed]
- Enøe, C.; Mousing, J.; Schirmer, A.L.; Willeberg, P. Infectious and Rearing-System Related Risk Factors for Chronic Pleuritis in Slaughter Pigs. Prev. Vet. Med. 2002, 54, 337–349. [Google Scholar] [CrossRef]
- Fraile, L.; Alegre, A.; López-Jiménez, R.; Nofrarías, M.; Segalés, J. Risk Factors Associated with Pleuritis and Cranio-Ventral Pulmonary Consolidation in Slaughter-Aged Pigs. Vet. J. 2010, 184, 326–333. [Google Scholar] [CrossRef]
- Meyns, T.; Van Steelant, J.; Rolly, E.; Dewulf, J.; Haesebrouck, F.; Maes, D. A Cross-Sectional Study of Risk Factors Associated with Pulmonary Lesions in Pigs at Slaughter. Vet. J. 2011, 187, 388–392. [Google Scholar] [CrossRef]
- Blanchard, B.; Vena, M.M.; Cavalier, A.; Lannic, J.L.; Gouranton, J.; Kobisch, M. Electron Microscopic Observation of the Respiratory Tract of SPF Piglets Inoculated with Mycoplasma hyopneumoniae. Vet. Microbiol. 1992, 30, 329–341. [Google Scholar] [CrossRef]
- Ferraz, M.E.S.; Almeida, H.M.S.; Storino, G.Y.; Sonálio, K.; Souza, M.R.; Moura, C.A.A.; Costa, W.M.T.; Lunardi, L.; Linhares, D.C.L.; de Oliveira, L.G. Lung Consolidation Caused by Mycoplasma hyopneumoniae Has a Negative Effect on Productive Performance and Economic Revenue in Finishing Pigs. Prev. Vet. Med. 2020, 182, 105091. [Google Scholar] [CrossRef]
- Gauger, P.C.; Lager, K.M.; Vincent, A.L.; Opriessnig, T.; Kehrli, M.E.; Cheung, A.K. Postweaning Multisystemic Wasting Syndrome Produced in Gnotobiotic Pigs Following Exposure to Various Amounts of Porcine Circovirus Type 2a or Type 2b. Vet. Microbiol. 2011, 153, 229–239. [Google Scholar] [CrossRef]
- Helm, E.T.; Curry, S.M.; De Mille, C.M.; Schweer, W.P.; Burrough, E.R.; Zuber, E.A.; Lonergan, S.M.; Gabler, N.K. Impact of Porcine Reproductive and Respiratory Syndrome Virus on Muscle Metabolism of Growing Pigs. J. Anim. Sci. 2019, 97, 3213–3227. [Google Scholar] [CrossRef]
- Cho, H.M.; Kim, E.; Wickramasuriya, S.S.; Shin, T.K.; Heo, J.M. Growth and Gut Performance of Young Pigs in Response to Different Dietary Cellulose Concentration and Rearing Condition. Anim. Biosci. 2021, 34, 1653–1662. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.M.; Macelline, S.P.; Wickramasuriya, S.S.; Shin, T.K.; Kim, E.; Son, H.C.; Heo, J.M. Moderate Dietary Boron Supplementation Improved Growth Performance, Crude Protein Digestibility and Diarrhea Index in Weaner Pigs Regardless of the Sanitary Condition. Anim. Biosci. 2022, 35, 434–443. [Google Scholar] [CrossRef]
- Le Floc’h, N.; LeBellego, L.; Matte, J.J.; Melchior, D.; Sève, B. The Effect of Sanitary Status Degradation and Dietary Tryptophan Content on Growth Rate and Tryptophan Metabolism in Weaning Pigs. J. Anim. Sci. 2009, 87, 1686–1694. [Google Scholar] [CrossRef] [Green Version]
- Montagne, L.; Le Floc’h, N.; Arturo-Schaan, M.; Foret, R.; Urdaci, M.C.; Le Gall, M. Comparative Effects of Level of Dietary Fiber and Sanitary Conditions on the Growth and Health of Weanling Pigs. J. Anim. Sci. 2012, 90, 2556–2569. [Google Scholar] [CrossRef] [PubMed]
- te Pas, M.F.W.; Jansman, A.J.M.; Kruijt, L.; van der Meer, Y.; Vervoort, J.J.M.; Schokker, D. Sanitary Conditions Affect the Colonic Microbiome and the Colonic and Systemic Metabolome of Female Pigs. Front. Vet. Sci. 2020, 7, 585730. [Google Scholar] [CrossRef] [PubMed]
- Waititu, S.M.; Yin, F.; Patterson, R.; Yitbarek, A.; Rodriguez-Lecompte, J.C.; Nyachoti, C.M. Dietary Supplementation with a Nucleotide-Rich Yeast Extract Modulates Gut Immune Response and Microflora in Weaned Pigs in Response to a Sanitary Challenge. Animal 2017, 11, 2156–2164. [Google Scholar] [CrossRef]
- Zhao, J.; Harper, A.F.; Estienne, M.J.; Webb, K.E., Jr.; McElroy, A.P.; Denbow, D.M. Growth Performance and Intestinal Morphology Responses in Early Weaned Pigs to Supplementation of Antibiotic-Free Diets with an Organic Copper Complex and Spray-Dried Plasma Protein in Sanitary and Nonsanitary Environments. J. Anim. Sci. 2007, 85, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Ridder, K.; Levesque, C.L.; Htoo, J.K.; de Lange, C.F.M. Immune System Stimulation Reduces the Efficiency of Tryptophan Utilization for Body Protein Deposition in Growing Pigs. J. Anim. Sci. 2012, 90, 3485–3491. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.W.; Mellencamp, M.A.; Carroll, J.A.; Boyd, R.D.; Allee, G.L. Acute Feed Intake and Acute-Phase Protein Responses Following a Lipopolysaccharide Challenge in Pigs from Two Dam Lines. Vet. Immunol. Immunopath. 2005, 107, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.Y.; Sun, L.H.; Lin, Y.C.; Ma, X.Y.; Zheng, C.T.; Zhou, G.L.; Chen, F.; Zou, S.T. Effects of Dietary Glycyl-Glutamine on Growth Performance, Small Intestinal Integrity, and Immune Responses of Weaning Piglets Challenged with Lipopolysaccharide. J. Anim. Sci. 2009, 87, 4050–4056. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.W.; von Borell, E. Lipopolysaccharide-Induced Sickness Behavior in Pigs Is Inhibited by Pretreatment with Indomethacin. J. Anim. Sci. 1994, 72, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Litvak, N.; Rakhshandeh, A.; Htoo, J.K.; de Lange, C.F.M. Immune System Stimulation Increases the Optimal Dietary Methionine to Methionine plus Cysteine Ratio in Growing Pigs. J. Anim. Sci. 2013, 91, 4188–4196. [Google Scholar] [CrossRef] [PubMed]
- Roth, J.; Conn, C.A.; Kluger, M.J.; Zeisberger, E. Kinetics of Systemic and Intrahypothalamic IL-6 and Tumor Necrosis Factor during Endotoxin Fever in Guinea Pigs. Am. J. Physiol.-Reg. I 1993, 265, R653–R658. [Google Scholar] [CrossRef]
- Wu, C.; Xu, Q.; Wang, R.; Qin, L.; Peng, X.; Hu, L.; Liu, Y.; Fang, Z.; Lin, Y.; Xu, S.; et al. Effects of Dietary β-Glucan Supplementation on Growth Performance and Immunological and Metabolic Parameters of Weaned Pigs Administered with Escherichia coli Lipopolysaccharide. Food Funct. 2018, 9, 3338–3343. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Lahaye, L.; He, Z.; Zhang, J.; Yang, C.; Piao, X. Micro-Encapsulated Essential Oils and Organic Acids Combination Improves Intestinal Barrier Function, Inflammatory Responses and Microbiota of Weaned Piglets Challenged with Enterotoxigenic Escherichia coli F4 (K88+). Anim. Nutr. 2020, 6, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Xie, L.; Liu, B.; Zhou, L.; Hu, Y.; Ajuwon, K.M.; Fang, R. Dietary Supplementation of EGF Ameliorates the Negatively Effects of LPS on Early-Weaning Piglets: From Views of Growth Performance, Nutrient Digestibility, Microelement Absorption and Possible Mechanisms. Animals 2021, 11, 1598. [Google Scholar] [CrossRef] [PubMed]
- Brockmeier, S.L.; Halbur, P.G.; Thacker, E.L. Porcine Respiratory Disease Complex. In Polymicrobial Diseases; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2002; pp. 231–258. ISBN 978-1-68367-232-6. [Google Scholar]
- Thacker, E.L. Immunology of the Porcine Respiratory Disease Complex. Vet. Clin. N. Am.-Food A 2001, 17, 551–565. [Google Scholar] [CrossRef]
- Hansen, M.S.; Pors, S.E.; Jensen, H.E.; Bille-Hansen, V.; Bisgaard, M.; Flachs, E.M.; Nielsen, O.L. An Investigation of the Pathology and Pathogens Associated with Porcine Respiratory Disease Complex in Denmark. J. Comp. Pathol. 2010, 143, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Kyriazakis, I.; Emmans, G.C. The Growth of Mammals Following a Period of Nutritional Limitation. J. Theor. Biol. 1992, 156, 485–498. [Google Scholar] [CrossRef]
- Smith, B.N.; Oelschlager, M.L.; Abdul Rasheed, M.S.; Dilger, R.N. Dietary Soy Isoflavones Reduce Pathogen-Related Mortality in Growing Pigs under Porcine Reproductive and Respiratory Syndrome Viral Challenge. J. Anim. Sci. 2020, 98, skaa024. [Google Scholar] [CrossRef]
- Schweer, W.P.; Schwartz, K.; Burrough, E.R.; Yoon, K.J.; Sparks, J.C.; Gabler, N.K. The Effect of Porcine Reproductive and Respiratory Syndrome Virus and Porcine Epidemic Diarrhea Virus Challenge on Growing Pigs: Growth Performance and Digestibility1. J. Anim. Sci. 2016, 94, 514–522. [Google Scholar] [CrossRef] [Green Version]
- Kahindi, R.; Regassa, A.; Htoo, J.; Nyachoti, M. Optimal Sulfur Amino Acid to Lysine Ratio for Post Weaning Piglets Reared under Clean or Unclean Sanitary Conditions. Anim. Nutr. 2017, 3, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, B.; Htoo, J.K.; Nyachoti, C.M. Effects of Different Dietary Tryptophan: Lysine Ratios and Sanitary Conditions on Growth Performance, Plasma Urea Nitrogen, Serum Haptoglobin and Ileal Histomorphology of Weaned Pigs. Anim. Sci. J. 2017, 88, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.M.; González-Ortiz, G.; Melo-Durán, D.; Heo, J.M.; Cordero, G.; Bedford, M.R.; Kim, J.C. Stimbiotic Supplementation Improved Performance and Reduced Inflammatory Response via Stimulating Fiber Fermenting Microbiome in Weaner Pigs Housed in a Poor Sanitary Environment and Fed an Antibiotic-Free Low Zinc Oxide Diet. PLoS ONE 2020, 15, e0240264. [Google Scholar] [CrossRef] [PubMed]
- Bannerman, D.D.; Goldblum, S.E. Mechanisms of Bacterial Lipopolysaccharide-Induced Endothelial Apoptosis. Am. J. PHysiol.-Lung C 2003, 284, L899–L914. [Google Scholar] [CrossRef] [PubMed]
- Kang, P.; Zhang, L.; Hou, Y.; Ding, B.; Yi, D.; Wang, L.; Zhu, H.; Liu, Y.; Yin, Y.; Wu, G. Effects of L-Proline on the Growth Performance, and Blood Parameters in Weaned Lipopolysaccharide (LPS)-Challenged Pigs. Asian-Australas. J. Anim. Sci. 2014, 27, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Opal, S.M.; Scannon, P.J.; Vincent, J.-L.; White, M.; Carroll, S.F.; Palardy, J.E.; Parejo, N.A.; Pribble, J.P.; Lemke, J.H. Relationship between Plasma Levels of Lipopolysaccharide (LPS) and LPS-Binding Protein in Patients with Severe Sepsis and Septic Shock. J. Infect. Dis. 1999, 180, 1584–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfieri, A.; Watson, J.J.; Kammerer, R.A.; Tasab, M.; Progias, P.; Reeves, K.; Brown, N.J.; Brookes, Z.L. Angiopoietin-1 Variant Reduces LPS-Induced Microvascular Dysfunction in a Murine Model of Sepsis. Crit. Care 2012, 16, R182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeMay, L.G.; Vander, A.J.; Kluger, M.J. Role of Interleukin 6 in Fever in Rats. Am. J. Physiol.-Reg. I 1990, 258, R798–R803. [Google Scholar] [CrossRef] [PubMed]
- Bikker, P.; Dirkzwager, A.; Fledderus, J.; Trevisi, P.; le Huërou-Luron, I.; Lallès, J.P.; Awati, A. The Effect of Dietary Protein and Fermentable Carbohydrates Levels on Growth Performance and Intestinal Characteristics in Newly Weaned Piglets. J. Anim. Sci. 2006, 84, 3337–3345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Wielen, N.v.d.; Hee, B.v.d.; Wang, J.; Hendriks, W.; Gilbert, M. Impact of Fermentable Protein, by Feeding High Protein Diets, on Microbial Composition, Microbial Catabolic Activity, Gut Health and beyond in Pigs. Microorganisms 2020, 8, 1735. [Google Scholar] [CrossRef] [PubMed]
- Prohászka, L.; Baron, F. The Predisposing Role of High Dietary Protein Supplies in Enteropathogenic E. coli Infections of Weaned Pigs. Zent. Vet. Riehe B 1980, 27, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Nyachoti, C.M.; Omogbenigun, F.O.; Rademacher, M.; Blank, G. Performance Responses and Indicators of Gastrointestinal Health in Early-Weaned Pigs Fed Low-Protein Amino Acid-Supplemented Diets. J. Anim. Sci. 2006, 84, 125–134. [Google Scholar] [CrossRef]
- Pieper, R.; Kröger, S.; Richter, J.F.; Wang, J.; Martin, L.; Bindelle, J.; Htoo, J.K.; von Smolinski, D.; Vahjen, W.; Zentek, J.; et al. Fermentable Fiber Ameliorates Fermentable Protein-Induced Changes in Microbial Ecology, but Not the Mucosal Response, in the Colon of Piglets. J. Nutr. 2012, 142, 661–667. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.K.; Muir, J.G.; Gibson, P.R. Review Article: Insights into Colonic Protein Fermentation, Its Modulation and Potential Health Implications. Aliment. Pharm. Ther. 2016, 43, 181–196. [Google Scholar] [CrossRef]
- Wellock, I.J.; Fortomaris, P.D.; Houdijk, J.G.M.; Kyriazakis, I. Effects of Dietary Protein Supply, Weaning Age and Experimental Enterotoxigenic Escherichia coli Infection on Newly Weaned Pigs: Health. Animal 2008, 2, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Rist, V.T.S.; Weiss, E.; Eklund, M.; Mosenthin, R. Impact of Dietary Protein on Microbiota Composition and Activity in the Gastrointestinal Tract of Piglets in Relation to Gut Health: A Review. Animal 2013, 7, 1067–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, J.M.; Kim, J.C.; Hansen, C.F.; Mullan, B.P.; Hampson, D.J.; Pluske, J.R. Feeding a Diet with Decreased Protein Content Reduces Indices of Protein Fermentation and the Incidence of Postweaning Diarrhea in Weaned Pigs Challenged with an Enterotoxigenic Strain of Escherichia coli. J. Anim. Sci. 2009, 87, 2833–2843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, J.M.; Kim, J.C.; Yoo, J.; Pluske, J.R. A Between-Experiment Analysis of Relationships Linking Dietary Protein Intake and Post-Weaning Diarrhea in Weanling Pigs under Conditions of Experimental Infection with an Enterotoxigenic Strain of Escherichia coli. Anim. Sci. J. 2015, 86, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Pollock, J.; Hutchings, M.R.; Hutchings, K.E.K.; Gally, D.L.; Houdijk, J.G.M. Changes in the Ileal, but Not Fecal, Microbiome in Response to Increased Dietary Protein Level and Enterotoxigenic Escherichia coli Exposure in Pigs. Appl. Environ. Microb. 2019, 85, e01252-19. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, M.S.; Ijssennagger, N.; Kies, A.K.; van Mil, S.W.C. Protein Fermentation in the Gut; Implications for Intestinal Dysfunction in Humans, Pigs, and Poultry. Am. J. Physiol.-Gastr. L 2018, 315, G159–G170. [Google Scholar] [CrossRef] [PubMed]
- Lallès, J.-P.; Montoya, C.A. Dietary Alternatives to In-Feed Antibiotics, Gut Barrier Function and Inflammation in Piglets Post-Weaning: Where Are We Now? Anim. Feed Sci. Technol. 2021, 274, 114836. [Google Scholar] [CrossRef]
- Yu, D.; Zhu, W.; Hang, S. Effects of Low-Protein Diet on the Intestinal Morphology, Digestive Enzyme Activity, Blood Urea Nitrogen, and Gut Microbiota and Metabolites in Weaned Pigs. Arch. Anim. Nutr. 2019, 73, 287–305. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Song, P.; Fan, P.; He, T.; Jacobs, D.; Levesque, C.L.; Johnston, L.J.; Ji, L.; Ma, N.; Chen, Y.; et al. Moderate Dietary Protein Restriction Optimized Gut Microbiota and Mucosal Barrier in Growing Pig Model. Front. Cell. Infect. Microbiol. 2018, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Spring, S.; Premathilake, H.; Bradway, C.; Shili, C.; DeSilva, U.; Carter, S.; Pezeshki, A. Effect of Very Low-Protein Diets Supplemented with Branched-Chain Amino Acids on Energy Balance, Plasma Metabolomics and Fecal Microbiome of Pigs. Sci. Rep. 2020, 10, 15859. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Qiao, S.; Ren, M.; Zeng, X.; Ma, X.; Wu, Z.; Thacker, P.; Wu, G. Supplementation with Branched-Chain Amino Acids to a Low-Protein Diet Regulates Intestinal Expression of Amino Acid and Peptide Transporters in Weanling Pigs. Amino Acids 2013, 45, 1191–1205. [Google Scholar] [CrossRef] [PubMed]
- Jarrett, S.; Ashworth, C.J. The Role of Dietary Fibre in Pig Production, with a Particular Emphasis on Reproduction. J. Anim. Sci. Biotechnol. 2018, 9, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diether, N.E.; Willing, B.P. Microbial Fermentation of Dietary Protein: An Important Factor in Diet–Microbe–Host Interaction. Microorganisms 2019, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Williams, B.A.; Mikkelsen, D.; Flanagan, B.M.; Gidley, M.J. “Dietary Fibre”: Moving beyond the “Soluble/Insoluble” Classification for Monogastric Nutrition, with an Emphasis on Humans and Pigs. J. Anim. Sci. Biotechnol. 2019, 10, 45. [Google Scholar] [CrossRef] [Green Version]
- Htoo, J.K.; Araiza, B.A.; Sauer, W.C.; Rademacher, M.; Zhang, Y.; Cervantes, M.; Zijlstra, R.T. Effect of Dietary Protein Content on Ileal Amino Acid Digestibility, Growth Performance, and Formation of Microbial Metabolites in Ileal and Cecal Digesta of Early-Weaned Pigs1,2. J. Anim. Sci. 2007, 85, 3303–3312. [Google Scholar] [CrossRef]
- Jeaurond, E.A.; Rademacher, M.; Pluske, J.R.; Zhu, C.H.; de Lange, C.F.M. Impact of Feeding Fermentable Proteins and Carbohydrates on Growth Performance, Gut Health and Gastrointestinal Function of Newly Weaned Pigs. Can. J. Anim. Sci. 2008, 88, 271–281. [Google Scholar] [CrossRef]
- Molist, F.; van Oostrum, M.; Pérez, J.F.; Mateos, G.G.; Nyachoti, C.M.; van der Aar, P.J. Relevance of Functional Properties of Dietary Fibre in Diets for Weanling Pigs. Anim. Feed Sci. Technol. 2014, 189, 1–10. [Google Scholar] [CrossRef]
- Jha, R.; Berrocoso, J.D. Review: Dietary Fiber Utilization and Its Effects on Physiological Functions and Gut Health of Swine. Animal 2015, 9, 1441–1452. [Google Scholar] [CrossRef] [Green Version]
- Che, L.; Chen, H.; Yu, B.; He, J.; Zheng, P.; Mao, X.; Yu, J.; Huang, Z.; Chen, D. Long-Term Intake of Pea Fiber Affects Colonic Barrier Function, Bacterial and Transcriptional Profile in Pig Model. Nutr. Cancer 2014, 66, 388–399. [Google Scholar] [CrossRef]
- Li, Q.; Peng, X.; Burrough, E.R.; Sahin, O.; Gould, S.A.; Gabler, N.K.; Loving, C.L.; Dorman, K.S.; Patience, J.F. Dietary Soluble and Insoluble Fiber With or Without Enzymes Altered the Intestinal Microbiota in Weaned Pigs Challenged With Enterotoxigenic E. coli F18. Front. Microbiol. 2020, 11, 1110. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Burrough, E.R.; Gabler, N.K.; Loving, C.L.; Sahin, O.; Gould, S.A.; Patience, J.F. A Soluble and Highly Fermentable Dietary Fiber with Carbohydrases Improved Gut Barrier Integrity Markers and Growth Performance in F18 ETEC Challenged Pigs. J. Anim. Sci. 2019, 97, 2139–2153. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, L.L.; Jakobsen, M.; Jensen, B.B. Effects of Dietary Oligosaccharides on Microbial Diversity and Fructo-Oligosaccharide Degrading Bacteria in Faeces of Piglets Post-Weaning. Anim. Feed Sci. Technol. 2003, 109, 133–150. [Google Scholar] [CrossRef]
- Konstantinov, S.R.; Awati, A.; Smidt, H.; Williams, B.A.; Akkermans, A.D.L.; de Vos, W.M. Specific Response of a Novel and Abundant Lactobacillus amylovorus-Like Phylotype to Dietary Prebiotics in the Guts of Weaning Piglets. Appl. Environ. Microb. 2004, 70, 3821–3830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Piao, X.S.; Kim, S.W.; Wang, L.; Shen, Y.B.; Lee, H.S.; Li, S.Y. Effects of Chito-Oligosaccharide Supplementation on the Growth Performance, Nutrient Digestibility, Intestinal Morphology, and Fecal Shedding of Escherichia coli and Lactobacillus in Weaning Pigs. J. Anim. Sci. 2008, 86, 2609–2618. [Google Scholar] [CrossRef] [Green Version]
- Molist, F.; Gómez de Segura, A.; Pérez, J.F.; Bhandari, S.K.; Krause, D.O.; Nyachoti, C.M. Effect of Wheat Bran on the Health and Performance of Weaned Pigs Challenged with Escherichia coli K88+. Livest. Sci. 2010, 133, 214–217. [Google Scholar] [CrossRef]
- Bach Knudsen, K.E.; Hedemann, M.S.; Lærke, H.N. The Role of Carbohydrates in Intestinal Health of Pigs. Anim. Feed Sci. Technol. 2012, 173, 41–53. [Google Scholar] [CrossRef]
- Taciak, M.; Barszcz, M.; Święch, E.; Tuśnio, A.; Bachanek, I. Interactive Effects of Protein and Carbohydrates on Production of Microbial Metabolites in the Large Intestine of Growing Pigs. Arch. Anim. Nutr. 2017, 71, 192–209. [Google Scholar] [CrossRef]
- Mariscal-Landín, G.; Sève, B.; Colléaux, Y.; Lebreton, Y. Endogenous Amino Nitrogen Collected from Pigs with End-to-End Ileorectal Anastomosis Is Affected by the Method of Estimation and Altered by Dietary Fiber. J. Nutr. 1995, 125, 136–146. [Google Scholar] [CrossRef]
- Schmidt-Wittig, U.; Enss, M.-L.; Coenen, M.; Gärtner, K.; Hedrich, H.J. Response of Rat Colonic Mucosa to a High Fiber Diet. Ann. Nutr. Metab. 1996, 40, 343–350. [Google Scholar] [CrossRef]
- Hedemann, M.S.; Eskildsen, M.; Lærke, H.N.; Pedersen, C.; Lindberg, J.E.; Laurinen, P.; Knudsen, K.E.B. Intestinal Morphology and Enzymatic Activity in Newly Weaned Pigs Fed Contrasting Fiber Concentrations and Fiber Properties. J. Anim. Sci. 2006, 84, 1375–1386. [Google Scholar] [CrossRef] [PubMed]
- Wellington, M.O.; Hamonic, K.; Krone, J.E.C.; Htoo, J.K.; Van Kessel, A.G.; Columbus, D.A. Effect of Dietary Fiber and Threonine Content on Intestinal Barrier Function in Pigs Challenged with Either Systemic E. Coli Lipopolysaccharide or Enteric Salmonella Typhimurium. J. Anim. Sci. Biotechnol. 2020, 11, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluske, J.R.; Siba, P.M.; Pethick, D.W.; Durmic, Z.; Mullan, B.P.; Hampson, D.J. The Incidence of Swine Dysentery in Pigs Can Be Reduced by Feeding Diets That Limit the Amount of Fermentable Substrate Entering the Large Intestine. J. Nutr. 1996, 126, 2920–2933. [Google Scholar] [CrossRef]
- Dritz, S.S.; Shi, J.; Kielian, T.L.; Goodband, R.D.; Nelssen, J.L.; Tokach, M.D.; Chengappa, M.M.; Smith, J.E.; Blecha, F. Influence of Dietary β-Glucan on Growth Performance, Nonspecific Immunity, and Resistance to Streptococcus suis Infection in Weanling Pigs. J. Anim. Sci. 1995, 73, 3341–3350. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Swine, 11th ed.; The National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Li, D.F.; Nelssen, J.L.; Reddy, P.G.; Blecha, F.; Hancock, J.D.; Allee, G.L.; Goodband, R.D.; Klemm, R.D. Transient Hypersensitivity to Soybean Meal in the Early-Weaned Pig. J. Anim. Sci. 1990, 68, 1790–1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoner, G.R.; Allee, G.L.; Nelssen, J.L.; Johnston, M.E.; Goodband, R.D. Effect of Select Menhaden Fish Meal in Starter Diets for Pigs. J. Anim. Sci. 1990, 68, 2729–2735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peace, R.M.; Campbell, J.; Polo, J.; Crenshaw, J.; Russell, L.; Moeser, A. Spray-Dried Porcine Plasma Influences Intestinal Barrier Function, Inflammation, and Diarrhea in Weaned Pigs. J. Nutr. 2011, 141, 1312–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahan, D.C.; Fastinger, N.D.; Peters, J.C. Effects of Diet Complexity and Dietary Lactose Levels during Three Starter Phases on Postweaning Pig Performance. J. Anim. Sci. 2004, 82, 2790–2797. [Google Scholar] [CrossRef]
- Dritz, S.S.; Owen, K.Q.; Goodband, R.D.; Nelssen, J.L.; Tokach, M.D.; Chengappa, M.M.; Blecha, F. Influence of Lipopolysaccharide-Induced Immune Challenge and Diet Complexity on Growth Performance and Acute-Phase Protein Production in Segregated Early-Weaned Pigs. J. Anim. Sci. 1996, 74, 1620–1628. [Google Scholar] [CrossRef] [Green Version]
- Wolter, B.F.; Ellis, M.; Corrigan, B.P.; DeDecker, J.M.; Curtis, S.E.; Parr, E.N.; Webel, D.M. Impact of Early Postweaning Growth Rate as Affected by Diet Complexity and Space Allocation on Subsequent Growth Performance of Pigsin a Wean-to-Finish Production System. J. Anim. Sci. 2003, 81, 353–359. [Google Scholar] [CrossRef]
- Koo, B.; Kim, J.W.; de Lange, C.F.M.; Hossain, M.M.; Nyachoti, C.M. Effects of Diet Complexity and Multicarbohydrase Supplementation on Growth Performance, Nutrient Digestibility, Blood Profile, Intestinal Morphology, and Fecal Score in Newly Weaned Pigs. J. Anim. Sci. 2017, 95, 4060–4071. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, J.; Wang, G.; Cai, S.; Zeng, X.; Qiao, S. Advances in Low-Protein Diets for Swine. J. Anim. Sci. Biotechnol. 2018, 9, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, R.G.; Biden, R.S. The Effect of Protein Nutrition between 5–5 and 20 Kg Live Weight on the Subsequent Performance and Carcass Quality of Pigs. Anim. Sci. 1978, 27, 223–228. [Google Scholar] [CrossRef]
- Skinner, L.D.; Levesque, C.L.; Wey, D.; Rudar, M.; Zhu, J.; Hooda, S.; de Lange, C.F.M. Impact of Nursery Feeding Program on Subsequent Growth Performance, Carcass Quality, Meat Quality, and Physical and Chemical Body Composition of Growing-Finishing Pigs. J. Anim. Sci. 2014, 92, 1044–1054. [Google Scholar] [CrossRef]
- Reeds, P.J.; Fjeld, C.R.; Jahoor, F. Do the Differences between the Amino Acid Compositions of Acute-Phase and Muscle Proteins Have a Bearing on Nitrogen Loss in Traumatic States? J. Nutr. 1994, 124, 906–910. [Google Scholar] [CrossRef]
- Quiniou, N.; Dourmad, J.-Y.; Noblet, J. Effect of Energy Intake on the Performance of Different Types of Pig from 45 to 100 Kg Body Weight. 1. Protein and Lipid Deposition. Anim. Sci. 1996, 63, 277–288. [Google Scholar] [CrossRef]
- Le Naou, T.; Le Floc’h, N.; Louveau, I.; Gilbert, H.; Gondret, F. Metabolic Changes and Tissue Responses to Selection on Residual Feed Intake in Growing Pigs. J. Anim. Sci. 2012, 90, 4771–4780. [Google Scholar] [CrossRef] [Green Version]
- Klasing, K.C. Nutrition and the Immune System. Br. Poult. Sci. 2007, 48, 525–537. [Google Scholar] [CrossRef]
- Jan-Willem Van Klinken, B.; Einerhand, A.W.C.; Büller, H.A.; Dekker, J. Strategic Biochemical Analysis of Mucins. Anal. Biochem. 1998, 265, 103–116. [Google Scholar] [CrossRef]
- Bauchart-Thevret, C.; Stoll, B.; Chacko, S.; Burrin, D.G. Sulfur Amino Acid Deficiency Upregulates Intestinal Methionine Cycle Activity and Suppresses Epithelial Growth in Neonatal Pigs. Am. J. Physiol.-Endoc. M 2009, 296, E1239–E1250. [Google Scholar] [CrossRef] [Green Version]
- Cebra, J.J. Influences of Microbiota on Intestinal Immune System Development. Am. J. Clin Nutr. 1999, 69, 1046s–1051s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, G.K.; Bertolo, R.F.; Adjiri-Awere, A.; Pencharz, P.B.; Ball, R.O. Adequate Oral Threonine Is Critical for Mucin Production and Gut Function in Neonatal Piglets. Am. J. Physiol.-Gastr. L 2007, 292, G1293–G1301. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Functional Amino Acids in Nutrition and Health. Amino Acids 2013, 45, 407–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Zong, E.Y.; Huang, P.F.; Yang, H.S.; Yan, S.L.; Li, J.Z.; Li, Y.L.; Ding, X.Q.; He, S.P.; Xiong, X.; et al. The Effects of Dietary Sulfur Amino Acids on Serum Biochemical Variables, Mucosal Amino Acid Profiles, and Intestinal Inflammation in Weaning Piglets. Livest. Sci. 2019, 220, 32–36. [Google Scholar] [CrossRef]
- Yan, S.; Long, L.; Zong, E.; Huang, P.; Li, J.; Li, Y.; Ding, X.; Xiong, X.; Yin, Y.; Yang, H. Dietary Sulfur Amino Acids Affect Jejunal Cell Proliferation and Functions by Affecting Antioxidant Capacity, Wnt/β-Catenin, and the Mechanistic Target of Rapamycin Signaling Pathways in Weaning Piglets. J. Anim. Sci. 2018, 96, 5124–5133. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xu, K.; Liu, H.; Liu, G.; Bai, M.; Peng, C.; Li, T.; Yin, Y. Impact of the Gut Microbiota on Intestinal Immunity Mediated by Tryptophan Metabolism. Front. Cell. Infect. Microbiol. 2018, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.; Dai, Z.; Liu, N.; Ji, Y.; Chen, J.; Zhang, Y.; Yang, Y.; Li, J.; Wu, Z.; Wu, G. Dietary L-Tryptophan Modulates the Structural and Functional Composition of the Intestinal Microbiome in Weaned Piglets. Front. Microbiol. 2018, 9, 1736. [Google Scholar] [CrossRef]
- Klasing, K.C. Nutritional Modulation of Resistance to Infectious Diseases. Poult. Sci. 1998, 77, 1119–1125. [Google Scholar] [CrossRef]
- Defa, L.; Changting, X.; Shiyan, Q.; Jinhui, Z.; Johnson, E.W.; Thacker, P.A. Effects of Dietary Threonine on Performance, Plasma Parameters and Immune Function of Growing Pigs. Anim. Feed Sci. Technol. 1999, 78, 179–188. [Google Scholar] [CrossRef]
- Wang, X.; Qiao, S.Y.; Liu, M.; Ma, Y.X. Effects of Graded Levels of True Ileal Digestible Threonine on Performance, Serum Parameters and Immune Function of 10–25kg Pigs. Anim. Feed Sci. Technol. 2006, 129, 264–278. [Google Scholar] [CrossRef]
- Rodrigues, L.A.; Panisson, J.C.; Kpogo, L.A.; González-Vega, J.C.; Htoo, J.K.; Van Kessel, A.G.; Columbus, D.A. Functional Amino Acid Supplementation Postweaning Mitigates the Response of Normal Birth Weight More than for Low Birth Weight Pigs to a Subsequent Salmonella Challenge. Animal 2022, 16, 100566. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, L.A.; Panisson, J.C.; Van Kessel, A.G.; Columbus, D.A. Functional Amino Acid Supplementation Attenuates the Negative Effects of Plant-Based Nursery Diets on the Response of Pigs to a Subsequent Salmonella Typhimurium Challenge. J. Anim. Sci. 2022, skac267. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.H.; Seok, W.J.; Kim, I.H. Organic Acids Mixture as a Dietary Additive for Pigs—A Review. Animals 2020, 10, 952. [Google Scholar] [CrossRef] [PubMed]
- Gómez-García, M.; Sol, C.; de Nova, P.J.G.; Puyalto, M.; Mesas, L.; Puente, H.; Mencía-Ares, Ó.; Miranda, R.; Argüello, H.; Rubio, P.; et al. Antimicrobial Activity of a Selection of Organic Acids, Their Salts and Essential Oils against Swine Enteropathogenic Bacteria. Porc. Health Manag. 2019, 5, 32. [Google Scholar] [CrossRef] [Green Version]
- Zentek, J.; Buchheit-Renko, S.; Ferrara, F.; Vahjen, W.; Kessel, A.G.V.; Pieper, R. Nutritional and Physiological Role of Medium-Chain Triglycerides and Medium-Chain Fatty Acids in Piglets. Anim. Health Res. Rev. 2011, 12, 83–93. [Google Scholar] [CrossRef]
- Gebru, E.; Lee, J.S.; Son, J.C.; Yang, S.Y.; Shin, S.A.; Kim, B.; Kim, M.K.; Park, S.C. Effect of Probiotic-, Bacteriophage-, or Organic Acid-Supplemented Feeds or Fermented Soybean Meal on the Growth Performance, Acute-Phase Response, and Bacterial Shedding of Grower Pigs Challenged with Salmonella enterica Serotype Typhimurium. J. Anim. Sci. 2010, 88, 3880–3886. [Google Scholar] [CrossRef] [PubMed]
- Calveyra, J.C.; Nogueira, M.G.; Kich, J.D.; Biesus, L.L.; Vizzotto, R.; Berno, L.; Coldebella, A.; Lopes, L.; Morés, N.; Lima, G.J.M.M.; et al. Effect of Organic Acids and Mannanoligosaccharide on Excretion of Salmonella Typhimurium in Experimentally Infected Growing Pigs. Res. Vet. Sci. 2012, 93, 46–47. [Google Scholar] [CrossRef] [Green Version]
- Martín-Peláez, S.; Costabile, A.; Hoyles, L.; Rastall, R.A.; Gibson, G.R.; La Ragione, R.M.; Woodward, M.J.; Mateu, E.; Martín-Orúe, S.M. Evaluation of the Inclusion of a Mixture of Organic Acids or Lactulose into the Feed of Pigs Experimentally Challenged with Salmonella Typhimurium. Vet. Microbiol. 2010, 142, 337–345. [Google Scholar] [CrossRef]
- Walsh, M.C.; Rostagno, M.H.; Gardiner, G.E.; Sutton, A.L.; Richert, B.T.; Radcliffe, J.S. Controlling Salmonella Infection in Weanling Pigs through Water Delivery of Direct-Fed Microbials or Organic Acids. Part I: Effects on Growth Performance, Microbial Populations, and Immune Status. J. Anim. Sci. 2012, 90, 261–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabà, L.; Litjens, R.; Allaart, J.; van den Hil, P.R. Feed Additive Blends Fed to Nursery Pigs Challenged with Salmonella. J. Anim. Sci. 2020, 98, skz382. [Google Scholar] [CrossRef] [Green Version]
- Owusu-Asiedu, A.; Nyachoti, C.M.; Marquardt, R.R. Response of Early-Weaned Pigs to an Enterotoxigenic Escherichia coli (K88) Challenge When Fed Diets Containing Spray-Dried Porcine Plasma or Pea Protein Isolate plus Egg Yolk Antibody, Zinc Oxide, Fumaric Acid, or Antibiotic. J. Anim. Sci. 2003, 81, 1790–1798. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.J.; Park, J.W.; Baek, D.H.; Kim, J.K.; Kim, I.H. Feeding the Blend of Organic Acids and Medium Chain Fatty Acids Reduces the Diarrhea in Piglets Orally Challenged with Enterotoxigenic Escherichia coli K88. Anim. Feed Sci. Technol. 2017, 224, 46–51. [Google Scholar] [CrossRef]
- Ren, C.; Zhou, Q.; Guan, W.; Lin, X.; Wang, Y.; Song, H.; Zhang, Y. Immune Response of Piglets Receiving Mixture of Formic and Propionic Acid Alone or with Either Capric Acid or Bacillus licheniformis after Escherichia coli Challenge. BioMed Res. Int. 2019, 2019, 6416187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez, M.J.; Berrios, R.; Stelzhammer, S.; Hohmann, M.; Verri, W., Jr.; Bracarense, A.P.F.R.L. Ingestion of Organic Acids and Cinnamaldehyde Improves Tissue Homeostasis of Piglets Exposed to Enterotoxic Escherichia coli (ETEC). J. Anim. Sci. 2020, 98, skaa012. [Google Scholar] [CrossRef]
- Han, Y.; Zhan, T.; Zhao, Q.; Tang, C.; Zhang, K.; Han, Y.; Zhang, J. Effects of Mixed Organic Acids and Medium Chain Fatty Acids as Antibiotic Alternatives on the Performance, Serum Immunity, and Intestinal Health of Weaned Piglets Orally Challenged with Escherichia coli K88. Anim. Feed Sci. Technol. 2020, 269, 114617. [Google Scholar] [CrossRef]
- Stensland, I.; Kim, J.C.; Bowring, B.; Collins, A.M.; Mansfield, J.P.; Pluske, J.R. A Comparison of Diets Supplemented with a Feed Additive Containing Organic Acids, Cinnamaldehyde and a Permeabilizing Complex, or Zinc Oxide, on Post-Weaning Diarrhoea, Selected Bacterial Populations, Blood Measures and Performance in Weaned Pigs Experimentally Infected with Enterotoxigenic E. coli. Animals 2015, 5, 1147–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamwal, K.; Bhattacharya, S.; Puri, S. Plant Growth Regulator Mediated Consequences of Secondary Metabolites in Medicinal Plants. J. Appl. Res. Med. Aromat. Plants 2018, 9, 26–38. [Google Scholar] [CrossRef]
- Nath, H.; Samtiya, M.; Dhewa, T. Beneficial Attributes and Adverse Effects of Major Plant-Based Foods Anti-Nutrients on Health: A Review. Hum. Nutr. Metab. 2022, 28, 200147. [Google Scholar] [CrossRef]
- Redondo, L.; Chacana, P.; Dominguez, J.; Fernandez Miyakawa, M. Perspectives in the Use of Tannins as Alternative to Antimicrobial Growth Promoter Factors in Poultry. Front. Microbiol. 2014, 5, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaak, C.K.; Petkau, J.C.; Karmin, O.; Ominski, K.; Rodriguez-Lecompte, J.C.; Siow, Y.L. Seasonal Variations in Phenolic Compounds and Antioxidant Capacity of Cornus Stolonifera Plant Material: Applications in Agriculture. Can. J. Plant Sci. 2013, 93, 725–734. [Google Scholar] [CrossRef] [Green Version]
- Rossi, B.; Toschi, A.; Piva, A.; Grilli, E. Single Components of Botanicals and Nature-Identical Compounds as a Non-Antibiotic Strategy to Ameliorate Health Status and Improve Performance in Poultry and Pigs. Nutr. Res. Rev. 2020, 33, 218–234. [Google Scholar] [CrossRef]
- Omonijo, F.A.; Ni, L.; Gong, J.; Wang, Q.; Lahaye, L.; Yang, C. Essential Oils as Alternatives to Antibiotics in Swine Production. Anim. Nutr. 2018, 4, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhou, Y.F.; Zou, Y.; Hu, X.M.; Zheng, L.F.; Wei, H.K.; Giannenas, I.; Jin, L.Z.; Peng, J.; Jiang, S.W. Effects of Dietary Oregano Essential Oil Supplementation on the Stress Response, Antioxidative Capacity, and HSPs MRNA Expression of Transported Pigs. Livest. Sci. 2015, 180, 143–149. [Google Scholar] [CrossRef]
- Su, G.; Zhou, X.; Wang, Y.; Chen, D.; Chen, G.; Li, Y.; He, J. Effects of Plant Essential Oil Supplementation on Growth Performance, Immune Function and Antioxidant Activities in Weaned Pigs. Lipids Health Dis. 2018, 17, 139. [Google Scholar] [CrossRef] [Green Version]
- Li, S.Y.; Ru, Y.J.; Liu, M.; Xu, B.; Péron, A.; Shi, X.G. The Effect of Essential Oils on Performance, Immunity and Gut Microbial Population in Weaner Pigs. Livest. Sci. 2012, 145, 119–123. [Google Scholar] [CrossRef]
- Lan, R.X.; Li, T.S.; Kim, I.H. Effects of Essential Oils Supplementation in Different Nutrient Densities on Growth Performance, Nutrient Digestibility, Blood Characteristics and Fecal Microbial Shedding in Weaning Pigs. Anim. Feed Sci. Technol. 2016, 214, 77–85. [Google Scholar] [CrossRef]
- Tian, Q.; Piao, X. Essential Oil Blend Could Decrease Diarrhea Prevalence by Improving Antioxidative Capability for Weaned Pigs. Animals 2019, 9, 847. [Google Scholar] [CrossRef] [Green Version]
- Ruzauskas, M.; Bartkiene, E.; Stankevicius, A.; Bernatoniene, J.; Zadeike, D.; Lele, V.; Starkute, V.; Zavistanaviciute, P.; Grigas, J.; Zokaityte, E.; et al. The Influence of Essential Oils on Gut Microbial Profiles in Pigs. Animals 2020, 10, 1734. [Google Scholar] [CrossRef]
- Cairo, P.L.G.; Gois, F.D.; Sbardella, M.; Silveira, H.; de Oliveira, R.M.; Allaman, I.B.; Cantarelli, V.S.; Costa, L.B. Effects of Dietary Supplementation of Red Pepper (Schinus Terebinthifolius Raddi) Essential Oil on Performance, Small Intestinal Morphology and Microbial Counts of Weanling Pigs. J. Sci. Food Agric. 2018, 98, 541–548. [Google Scholar] [CrossRef]
- Oh, H.J.; Kim, I.H.; Song, M.H.; Kwak, W.G.; Yun, W.; Lee, J.H.; Lee, C.H.; Oh, S.Y.; Liu, S.; An, J.S.; et al. Effects of Microencapsulated Complex of Organic Acids and Essential Oils on Growth Performance, Nutrient Retention, Blood Profiles, Fecal Microflora, and Lean Meat Percentage in Weaning to Finishing Pigs. Can. J. Anim. Sci. 2019, 99, 41–49. [Google Scholar] [CrossRef]
- Coddens, A.; Loos, M.; Vanrompay, D.; Remon, J.P.; Cox, E. Cranberry Extract Inhibits in Vitro Adhesion of F4 and F18+ Escherichia coli to Pig Intestinal Epithelium and Reduces in Vivo Excretion of Pigs Orally Challenged with F18+ Verotoxigenic E. coli. Vet. Microbiol. 2017, 202, 64–71. [Google Scholar] [CrossRef]
- Brenes, A.; Viveros, A.; Chamorro, S.; Arija, I. Use of Polyphenol-Rich Grape by-Products in Monogastric Nutrition. A Review. Anim. Feed Sci. Technol. 2016, 211, 1–17. [Google Scholar] [CrossRef]
- Kafantaris, I.; Stagos, D.; Kotsampasi, B.; Hatzis, A.; Kypriotakis, A.; Gerasopoulos, K.; Makri, S.; Goutzourelas, N.; Mitsagga, C.; Giavasis, I. Grape Pomace Improves Performance, Antioxidant Status, Fecal Microbiota and Meat Quality of Piglets. Animal 2018, 12, 246–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gessner, D.K.; Fiesel, A.; Most, E.; Dinges, J.; Wen, G.; Ringseis, R.; Eder, K. Supplementation of a Grape Seed and Grape Marc Meal Extract Decreases Activities of the Oxidative Stress-Responsive Transcription Factors NF-ΚB and Nrf2 in the Duodenal Mucosa of Pigs. Acta Vet. Scand. 2013, 55, 18. [Google Scholar] [CrossRef] [PubMed]


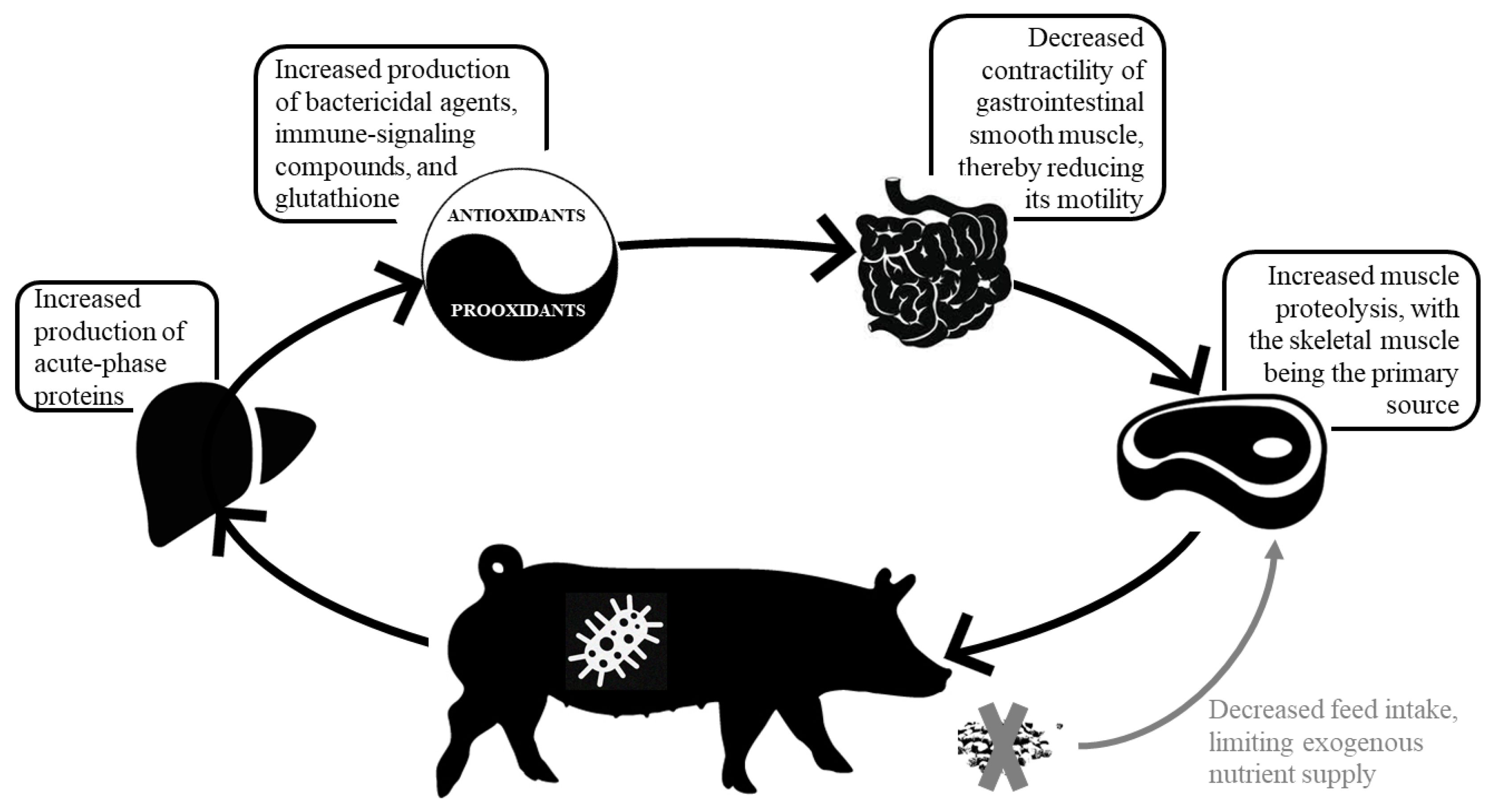{kind=link}
| Item. | Age (Weight) | Agent | Effect | Reference |
|---|---|---|---|---|
| Enteric pathogen challenge | 28 d (8.5 kg) | ETEC | Increased gut permeability, decreased small intestinal villus height | [44] |
| 21 d (6.9 kg) | ETEC | Increased fecal ETEC score, decreased amino acid digestibility, increased pH in the caecum and proximal colon | [46] | |
| 21 d (6.4 kg) | ETEC | Shorter duodenal villus height and deeper jejunal crypt depth | [50] | |
| - | ETEC | Production of toxins (heat-labile and heat-stable) in the small intestinal epithelial cell layer | [33] | |
| (4.9 kg) | ETEC | Intestinal damage and reduced nutrient digestibility | [43] | |
| 21 d (7.3 kg) | ETEC | Impaired mucosal immune function, intestinal morphology and integrity | [47] | |
| - | ST | Injection of Salmonella invasion protein into the intestinal epithelial cells, leading to cytoskeletal rearrangement | [38] | |
| (13.9 kg) | ST | Activation of acute-phase response, imbalance to antioxidant defense systems | [10] | |
| (22.6 kg) | ST | Decreased serum albumin concentration and increased haptoglobin concentration | [49] | |
| Respiratory pathogen challenge | - | AP MYC | Chronic pleurits | [51] |
| - | AP | Cranio-ventral pulmonary consolidation lesions | [52] | |
| - | AP MYC | Pleuritis and pneumonia | [53] | |
| - | MYC | Exfoliation of epithelial cells, increased peribronchiolar and perivascular monocellular cell accumulation | [54] | |
| 75 d | MYC | Lung consolidation | [55] | |
| - | PCV2 | Decrease in lymphocytes followed by an increase in neutrophils | [56] | |
| (13.1 kg) | PRRSV | Decreased expression of markers of skeletal muscle synthesis and increased liver glycogenolysis | [57] | |
| (33.6 kg) | PRRSV | Reduced digestibility of dry matter, energy and nitrogen, whole body accretion of lean, protein and fat | [12] | |
| Degradation of sanitary condition | 21 d (6.2 kg) | LSC | Lower apparent ileal and total tract digestibility of dry matter and lower apparent total tract digestibility of protein | [58] |
| 21 d | LSC | Decreased villus height and crypt depth in ileum | [59] | |
| 28 d (7.8 kg) | LSC | Greater plasma haptoglobin concentration and decreased Trp concentration | [60] | |
| 28 d | LSC | Increased diarrhea occurrence, higher counts of Lactobacillus and Enterobacteria and lower anaerobic sulfite bacteria, increased fecal volatile fatty acid concentration | [61] | |
| 7 d | LSC | Higher abundance of lactate fermenting microbes, altered amino acid metabolism, immune response, and microbiome-specific metabolites in blood | [62] | |
| 24 d | LSC | Increased serum haptoglobin, IgG antibody titers against keyhole limpet hemocyanin, pleuritis scores at slaughter | [1] | |
| 70 d | LSC | Reduced incremental protein efficiency | [2] | |
| 21 d | LSC | Impaired ileal immune response | [63] | |
| 18 d (6.0 kg) | LSC | Shorter villous length and lower crypt depth | [64] | |
| Bacterial lipopolysaccharide | (20.0 kg) | LPS | Increased plasma concentrations of acute-phase proteins and white blood cell counts, and decreased plasma albumin | [65] |
| (21.3 kg) | LPS | Increased serum amyloid concentration | [66] | |
| 14 d (4.5 kg) | LPS | Increased crypt depth of the duodenum and decreased ratio of villus height to crypt depth of the ileum | [67] | |
| - | LPS | Anorexia and fever | [68] | |
| (20.0 kg) | LPS | Increased eye temperature, and disturbed N balance | [69] | |
| - | LPS | Increase in interleukin-6 and tumor necrosis factor activity in plasma | [70] | |
| (20.5 kg) | LPS | Altered serum concentration of albumin, haptoglobin, fibrinogen, whole blood white blood cell, and platelet count | [18] | |
| 24 d (6.6 kg) | LPS | Increased rectal temperature, and serum concentrations of haptoglobin, tumor necrosis factor-α and interleukine-1 beta | [71] | |
| - | LPS | Increased plasma concentrations of cortisol, prostaglandin E2, interleukin-6, tumour necrosis factor-α and interleukin-1β | [72] | |
| 25 d | LPS | Decreased apparent digestibility of crude fat and microelement absorption | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, L.A.; Koo, B.; Nyachoti, M.; Columbus, D.A. Formulating Diets for Improved Health Status of Pigs: Current Knowledge and Perspectives. Animals 2022, 12, 2877. https://doi.org/10.3390/ani12202877
Rodrigues LA, Koo B, Nyachoti M, Columbus DA. Formulating Diets for Improved Health Status of Pigs: Current Knowledge and Perspectives. Animals. 2022; 12(20):2877. https://doi.org/10.3390/ani12202877
Chicago/Turabian StyleRodrigues, Lucas A., Bonjin Koo, Martin Nyachoti, and Daniel A. Columbus. 2022. "Formulating Diets for Improved Health Status of Pigs: Current Knowledge and Perspectives" Animals 12, no. 20: 2877. https://doi.org/10.3390/ani12202877