
Using Pedigree and Genomic Data toward Better Management of Inbreeding in Italian Dairy Sheep and Goat Breeds
 , , , ,
, , , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Breeds
2.2. Datasets and Quality Control
2.3. Pedigree Analysis
2.4. Genomic Inbreeding Calculation
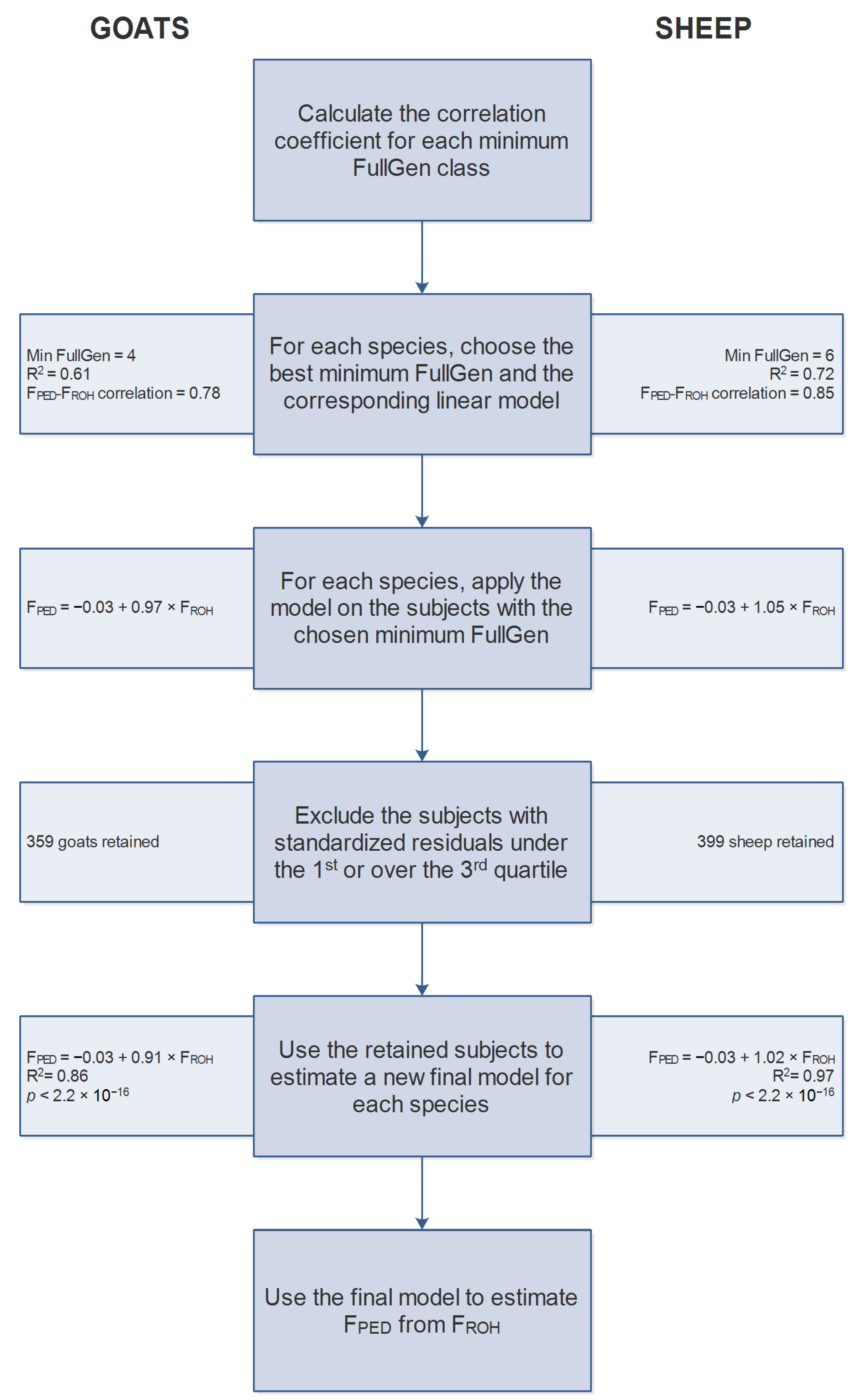
2.5. FPED-FROH Estimate
3. Results
3.1. Dataset Creation
3.2. Inbreeding Correlation and Linear Model
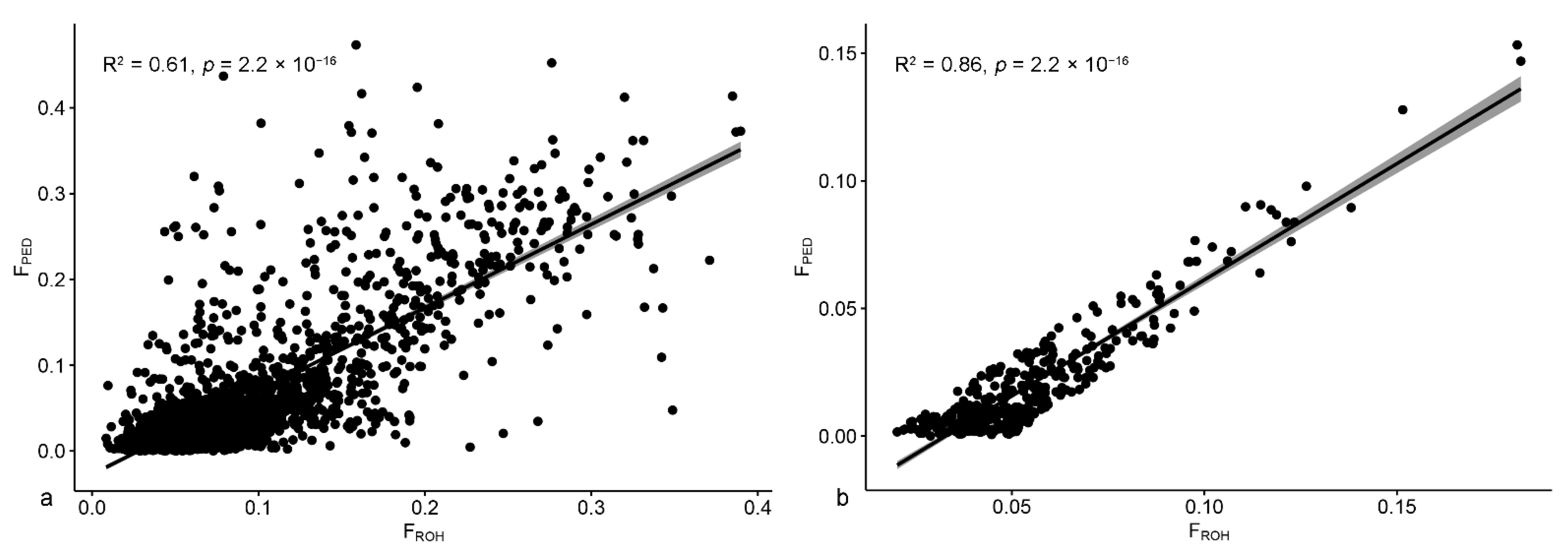
3.2.1. Goats
3.2.2. Sheep
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Panetto, J.; Machado, M.A.; da Silva, M.V.G.; Barbosa, R.S.; dos Santos, G.G.; Leite, R.D.M.; Peixoto, M.G.C. Parentage assignment using SNP markers, inbreeding and population size for the Brazilian Red Sindhi cattle. Livest. Sci. 2017, 204, 33–38. [Google Scholar] [CrossRef]
- Schiavo, G.; Bovo, S.; Bertolini, F.; Tinarelli, S.; Dall’Olio, S.; Costa, L.N.; Gallo, M.; Fontanesi, L. Comparative evaluation of genomic inbreeding parameters in seven commercial and autochthonous pig breeds. Animal 2020, 14, 910–920. [Google Scholar] [CrossRef] [PubMed]
- Polak, G.; Gurgul, A.; Jasielczuk, I.; Szmatoła, T.; Krupiński, J.; Bugno-Poniewierska, M. Suitability of Pedigree Information and Genomic Methods for Analyzing Inbreeding of Polish Cold-Blooded Horses Covered by Conservation Programs. Genes 2021, 12, 429. [Google Scholar] [CrossRef]
- Rodríguez-Ramilo, S.; Elsen, J.; Legarra, A. Inbreeding and effective population size in French dairy sheep: Comparison between genomic and pedigree estimates. J. Dairy Sci. 2019, 102, 4227–4237. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, B.; Zhang, T.; Yan, X.; Yu, Y.; Li, J.; Mei, B.; Wang, Z.; Zhang, Y.; Wang, R.; et al. Assessing Genetic Diversity and Estimating the Inbreeding Effect on Economic Traits of Inner Mongolia White Cashmere Goats Through Pedigree Analysis. Front. Vet. Sci. 2021, 8, 665872. [Google Scholar] [CrossRef] [PubMed]
- Hayes, B.J.; Goddard, M. Technical note: Prediction of breeding values using marker-derived relationship matrices. J. Anim. Sci. 2008, 86, 2089–2092. [Google Scholar] [CrossRef] [Green Version]
- Paiva, R.D.M.; de Sousa, J.E.R.; Ferreira, J.; Cunha, E.E.; de Paiva, M.P.S.L.M.; Gouveia, A.M.G.; Facó, O. Population structure and effect of inbreeding on milk yield of Saanen goats in Brazilian production systems. Small Rumin. Res. 2020, 192, 106194. [Google Scholar] [CrossRef]
- Legarra, A.; Baloche, G.; Barillet, F.; Astruc, J.; Soulas, C.; Aguerre, X.; Arrese, F.; Mintegi, L.; Lasarte, M.; Maeztu, F.; et al. Within- and across-breed genomic predictions and genomic relationships for Western Pyrenees dairy sheep breeds Latxa, Manech, and Basco-Béarnaise. J. Dairy Sci. 2014, 97, 3200–3212. [Google Scholar] [CrossRef] [Green Version]
- Villanueva, B.; Fernández, A.; Saura, M.; Caballero, A.; Fernández, J.; Morales-González, E.; Toro, M.A.; Pong-Wong, R. The value of genomic relationship matrices to estimate levels of inbreeding. Genet. Sel. Evol. 2021, 53, 42. [Google Scholar] [CrossRef]
- Andonov, S.; Lourenco, D.; Fragomeni, B.; Masuda, Y.; Pocrnic, I.; Tsuruta, S.; Misztal, I. Accuracy of breeding values in small genotyped populations using different sources of external information—A simulation study. J. Dairy Sci. 2017, 100, 395–401. [Google Scholar] [CrossRef]
- Cortes-Hernández, J.; García-Ruiz, A.; Vásquez-Peláez, C.G.; Ruiz-Lopez, F.D.J. Correlation of Genomic and Pedigree Inbreeding Coefficients in Small Cattle Populations. Animals 2021, 11, 3234. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Calus, M.P.; Guldbrandtsen, B.; Lund, M.S.; Sahana, G. Estimation of inbreeding using pedigree, 50k SNP chip genotypes and full sequence data in three cattle breeds. BMC Genet. 2015, 16, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero, A.; Villanueva, B.; Druet, T. On the estimation of inbreeding depression using different measures of inbreeding from molecular markers. Evol. Appl. 2021, 14, 416–428. [Google Scholar] [CrossRef]
- Ceballos, F.C.; Hazelhurst, S.; Ramsay, M. Assessing runs of Homozygosity: A comparison of SNP Array and whole genome sequence low coverage data. BMC Genom. 2018, 19, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weller, J.; Ezra, E.; Ron, M. Invited review: A perspective on the future of genomic selection in dairy cattle. J. Dairy Sci. 2017, 100, 8633–8644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciani, E.; Crepaldi, P.; Nicoloso, L.; Lasagna, E.; Sarti, F.M.; Moioli, B.; Napolitano, F.; Carta, A.; Usai, G.; D’Andrea, M.; et al. Genome-wide analysis of Italian sheep diversity reveals a strong geographic pattern and cryptic relationships between breeds. Anim. Genet. 2014, 45, 256–266. [Google Scholar] [CrossRef]
- Nicoloso, L.; The Italian Goat Consortium; Bomba, L.; Colli, L.; Negrini, R.; Milanesi, M.; Mazza, R.; Sechi, T.; Frattini, S.; Talenti, A.; et al. Genetic diversity of Italian goat breeds assessed with a medium-density SNP chip. Genet. Sel. Evol. 2015, 47, 62. [Google Scholar] [CrossRef]
- Mastrangelo, S.; Ciani, E.; Sardina, M.T.; Sottile, G.; Pilla, F.; Portolano, B. Runs of homozygosity reveal genome-wide autozygosity in Italian sheep breeds. Anim. Genet. 2018, 49, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Cortellari, M.; Barbato, M.; Talenti, A.; Bionda, A.; Carta, A.; Ciampolini, R.; Ciani, E.; Crisà, A.; Frattini, S.; Lasagna, E.; et al. The climatic and genetic heritage of Italian goat breeds with genomic SNP data. Sci. Rep. 2021, 11, 10986. [Google Scholar] [CrossRef]
- Carta, A.; Casu, S.; Salaris, S. Invited review: Current state of genetic improvement in dairy sheep. J. Dairy Sci. 2009, 92, 5814–5833. [Google Scholar] [CrossRef]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Wellmann, R. Optimum contribution selection for animal breeding and conservation: The R package optiSel. BMC Bioinform. 2019, 20, 25. [Google Scholar] [CrossRef] [PubMed]
- Wright, S. The genetical structure of populations. Ann. Eugen. 1949, 15, 323–354. [Google Scholar] [CrossRef] [PubMed]
- McQuillan, R.; Leutenegger, A.-L.; Abdel-Rahman, R.; Franklin, C.S.; Pericic, M.; Barac-Lauc, L.; Smolej-Narancic, N.; Janicijevic, B.; Polasek, O.; Tenesa, A.; et al. Runs of Homozygosity in European Populations. Am. J. Hum. Genet. 2008, 83, 359–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howrigan, D.P.; Simonson, M.A.; Keller, M.C. Detecting autozygosity through runs of homozygosity: A comparison of three autozygosity detection algorithms. BMC Genom. 2011, 12, 460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A Tool for Genome-wide Complex Trait Analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Meyermans, R.; Gorssen, W.; Buys, N.; Janssens, S. How to study runs of homozygosity using PLINK? A guide for analyzing medium density SNP data in livestock and pet species. BMC Genom. 2020, 21, 94. [Google Scholar] [CrossRef]
- VanRaden, P. Efficient Methods to Compute Genomic Predictions. J. Dairy Sci. 2008, 91, 4414–4423. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, I.; Fernandez, E.N.; Blasco, A.; Ravagnolo, O.; Legarra, A. Effects of ignoring inbreeding in model-based accuracy for BLUP and SSGBLUP. J. Anim. Breed. Genet. 2020, 137, 356–364. [Google Scholar] [CrossRef]
- Maltecca, C.; Tiezzi, F.; Cole, J.; Baes, C. Symposium review: Exploiting homozygosity in the era of genomics—Selection, inbreeding, and mating programs. J. Dairy Sci. 2020, 103, 5302–5313. [Google Scholar] [CrossRef]
- Teissier, M.; Larroque, H.; Robert-Granié, C. Weighted single-step genomic BLUP improves accuracy of genomic breeding values for protein content in French dairy goats: A quantitative trait influenced by a major gene. Genet. Sel. Evol. 2018, 50, 31. [Google Scholar] [CrossRef] [PubMed]
- Cesarani, A.; Sechi, T.; Gaspa, G.; Usai, M.G.; Sorbolini, S.; Macciotta, N.P.P.; Carta, A. Investigation of genetic diversity and selection signatures between Sarda and Sardinian Ancestral black, two related sheep breeds with evident morphological differences. Small Rumin. Res. 2019, 177, 68–75. [Google Scholar] [CrossRef]
- Zhu, S.; Guo, T.; Yuan, C.; Liu, J.; Li, J.; Han, M.; Zhao, H.; Wu, Y.; Sun, W.; Wang, X.; et al. Evaluation of Bayesian alphabet and GBLUP based on different marker density for genomic prediction in Alpine Merino sheep. G3 Genes Genomes Genet. 2021, 11, jkab206. [Google Scholar] [CrossRef] [PubMed]
- Colleau, J.-J. An indirect approach to the extensive calculation of relationship coefficients. Genet. Sel. Evol. 2002, 34, 409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar, I.; Misztal, I. Technical Note: Recursive Algorithm for Inbreeding Coefficients Assuming Nonzero Inbreeding of Unknown Parents. J. Dairy Sci. 2008, 91, 1669–1672. [Google Scholar] [CrossRef]
- Amin, N.; Van Duijn, C.M.; Aulchenko, Y.S. A Genomic Background Based Method for Association Analysis in Related Individuals. PLoS ONE 2007, 2, e1274. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Benyamin, B.; McEvoy, B.P.; Gordon, S.; Henders, A.; Nyholt, D.; Madden, P.A.; Heath, A.C.; Martin, N.; Montgomery, G.; et al. Common SNPs explain a large proportion of the heritability for human height. Nat. Genet. 2010, 42, 565–569. [Google Scholar] [CrossRef] [Green Version]
- Bjelland, D.; Weigel, K.; Vukasinovic, N.; Nkrumah, J. Evaluation of inbreeding depression in Holstein cattle using whole-genome SNP markers and alternative measures of genomic inbreeding. J. Dairy Sci. 2013, 96, 4697–4706. [Google Scholar] [CrossRef]
- Alemu, S.W.; Kadri, N.K.; Harland, C.; Faux, P.; Charlier, C.; Caballero, A.; Druet, T. An evaluation of inbreeding measures using a whole-genome sequenced cattle pedigree. Heredity 2021, 126, 410–423. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, J.; Cesarani, A.; Garcia, A.; Sumreddee, P.; Larios, N.; Mancin, E.; García, J.; Núñez, R.; Ramírez, R. Genetic Background and Inbreeding Depression in Romosinuano Cattle Breed in Mexico. Animals 2021, 11, 321. [Google Scholar] [CrossRef] [PubMed]
- Luan, T.; Yu, X.; Dolezal, M.; Bagnato, A.; Meuwissen, T.H. Genomic prediction based on runs of homozygosity. Genet. Sel. Evol. 2014, 46, 64. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/Breed | N. Subjects | Mean FPED | Mean FROH | Mean FGRM | FPED-FROH Correlation Coefficient (p-Value) | FPED-FGRM Correlation Coefficient (p-Value) | FROH-FGRM Correlation Coefficient (p-Value) |
|---|---|---|---|---|---|---|---|
| Goats | 3028 | 0.017 ± 0.039 | 0.059 ± 0.032 | 0.012 ± 0.046 | 0.278 (<0.0001) | 0.272 (<0.0001) | 0.604 (<0.0001) |
| Camosciata delle Alpi | 2093 | 0.016 ± 0.038 | 0.057 ± 0.033 | 0.013 ± 0.043 | 0.260 (<0.0001) | 0.228 (<0.0001) | 0.671 (<0.0001) |
| Saanen | 935 | 0.020 ± 0.042 | 0.062 ± 0.030 | 0.010 ± 0.051 | 0.312 (<0.0001) | 0.351 (<0.0001) | 0.492 (<0.0001) |
| Sheep | 2057 | 0.062 ± 0.076 | 0.097 ± 0.063 | 0.006 ± 0.064 | 0.817 (<0.0001) | 0.365 (<0.0001) | 0.477 (<0.0001) |
| Sarda | 1053 | 0.093 ± 0.095 | 0.135 ± 0.065 | 0.023 ± 0.073 | 0.804 (<0.0001) | 0.346 (<0.0001) | 0.489 (<0.0001) |
| Delle Langhe | 104 | 0.060 ± 0.037 | 0.099 ± 0.038 | −0.025 ± 0.127 | 0.436 (<0.0001) | 0.225 (0.022) | 0.354 (<0.0001) |
| Comisana | 529 | 0.018 ± 0.010 | 0.044 ± 0.016 | −0.008 ± 0.021 | 0.379 (<0.0001) | −0.130 (0.003) | 0.229 (<0.0001) |
| Massese | 371 | 0.039 ± 0.010 | 0.067 ± 0.017 | −0.013 ± 0.025 | 0.318 (<0.0001) | −0.070 (0.181) | 0.224 (<0.0001) |
| Minimum FullGen | N. Animals | N. Goats | N. Sheep | Correlation Coefficient | LRM Intercept | LRM Slope | LRM R2 |
|---|---|---|---|---|---|---|---|
| 0 | 5085 | 3028 | 2057 | 0.712 | −0.028 | 0.860 | 0.507 |
| 1 | 4549 | 2493 | 2056 | 0.725 | −0.028 | 0.888 | 0.526 |
| 2 | 3911 | 1877 | 2034 | 0.735 | −0.027 | 0.910 | 0.540 |
| 3 | 3311 | 1358 | 1953 | 0.753 | −0.026 | 0.937 | 0.567 |
| 4 | 2522 | 717 | 1805 | 0.782 | −0.027 | 0.971 | 0.611 |
| 5 | 1602 | 167 | 1435 | 0.825 | −0.028 | 1.010 | 0.681 |
| 6 | 927 | 18 | 909 | 0.849 | −0.030 | 1.056 | 0.720 |
| 7 | 378 | 2 | 376 | 0.847 | −0.029 | 1.085 | 0.718 |
| 8 | 107 | 0 | 107 | 0.773 | 0.008 | 0.997 | 0.597 |
| Breed | Estimated FPED Class | N. Subjects | FROH Mean | FROH SD | FROH 95% CI | FROH Range |
|---|---|---|---|---|---|---|
| Goats | 0.00–0.05 | 613 | 0.054 | 0.017 | 0.052–0.055 | 0.008–0.088 |
| Camosciata delle Alpi | 0.00–0.05 | 453 | 0.052 | 0.017 | 0.050–0.053 | 0.008–0.088 |
| Saanen | 0.00–0.05 | 160 | 0.058 | 0.015 | 0.056–0.061 | 0.024–0.088 |
| Goats | 0.05–0.10 | 81 | 0.103 | 0.013 | 0.101–0.106 | 0.088–0.138 |
| Camosciata delle Alpi | 0.05–0.10 | 49 | 0.102 | 0.011 | 0.099–0.105 | 0.089–0.132 |
| Saanen | 0.05–0.10 | 32 | 0.106 | 0.015 | 0.100–0.111 | 0.088–0.138 |
| Goats | 0.10–0.15 | 17 | 0.162 | 0.014 | 0.156–0.169 | 0.143–0.182 |
| Camosciata delle Alpi | 0.10–0.15 | 9 | 0.157 | 0.014 | 0.148–0.166 | 0.143–0.179 |
| Saanen | 0.10–0.15 | 8 | 0.168 | 0.012 | 0.159–0.176 | 0.153–0.182 |
| Breed | Estimated FPED Class | N. Subjects | FROH Mean | FROH SD | FROH 95% CI | FROH Range |
|---|---|---|---|---|---|---|
| Sheep | 0.00–0.05 | 419 | 0.055 | 0.014 | 0.054–0.057 | 0.022–0.078 |
| Comisana | 0.00–0.05 | 163 | 0.045 | 0.013 | 0.043–0.047 | 0.022–0.078 |
| Massese | 0.00–0.05 | 198 | 0.061 | 0.010 | 0.059–0.062 | 0.029–0.078 |
| Sarda | 0.00–0.05 | 33 | 0.068 | 0.009 | 0.065–0.071 | 0.041–0.078 |
| Delle Langhe | 0.00–0.05 | 25 | 0.067 | 0.012 | 0.062–0.072 | 0.027–0.078 |
| Sheep | 0.05–0.10 | 231 | 0.097 | 0.014 | 0.095–0.098 | 0.078–0.127 |
| Comisana | 0.05–0.10 | 2 | 0.092 | 0.014 | 0.072–0.111 | 0.082–0.101 |
| Massese | 0.05–0.10 | 59 | 0.090 | 0.011 | 0.087–0.093 | 0.078–0.120 |
| Sarda | 0.05–0.10 | 122 | 0.099 | 0.015 | 0.096–0.101 | 0.078–0.127 |
| Delle Langhe | 0.05–0.10 | 48 | 0.100 | 0.013 | 0.096–0.103 | 0.079–0.124 |
| Sheep | 0.10–0.15 | 89 | 0.150 | 0.015 | 0.146–0.153 | 0.128–0.176 |
| Comisana | 0.10–0.15 | 1 | 0.162 | |||
| Sarda | 0.10–0.15 | 79 | 0.149 | 0.014 | 0.146–0.152 | 0.128–0.176 |
| Delle Langhe | 0.10–0.15 | 9 | 0.152 | 0.018 | 0.140–0.164 | 0.128–0.175 |
| Sheep | 0.15–0.20 | 71 | 0.199 | 0.013 | 0.196–0.203 | 0.177–0.225 |
| Sarda | 0.15–0.20 | 68 | 0.199 | 0.013 | 0.196–0.203 | 0.177–0.225 |
| Delle Langhe | 0.15–0.20 | 3 | 0.203 | 0.022 | 0.178–0.227 | 0.179–0.222 |
| Sheep (Sarda) | 0.20–0.25 | 56 | 0.250 | 0.015 | 0.246–0.254 | 0.225–0.274 |
| Sheep (Sarda) | 0.25–0.30 | 27 | 0.293 | 0.014 | 0.288–0.298 | 0.275–0.321 |
| Sheep (Sarda) | 0.30–0.35 | 13 | 0.336 | 0.013 | 0.329–0.343 | 0.324–0.371 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortellari, M.; Negro, A.; Bionda, A.; Grande, S.; Cesarani, A.; Carta, A.; Macciotta, N.; Biffani, S.; Crepaldi, P. Using Pedigree and Genomic Data toward Better Management of Inbreeding in Italian Dairy Sheep and Goat Breeds. Animals 2022, 12, 2828. https://doi.org/10.3390/ani12202828
Cortellari M, Negro A, Bionda A, Grande S, Cesarani A, Carta A, Macciotta N, Biffani S, Crepaldi P. Using Pedigree and Genomic Data toward Better Management of Inbreeding in Italian Dairy Sheep and Goat Breeds. Animals. 2022; 12(20):2828. https://doi.org/10.3390/ani12202828
Chicago/Turabian StyleCortellari, Matteo, Alessio Negro, Arianna Bionda, Silverio Grande, Alberto Cesarani, Antonello Carta, Nicola Macciotta, Stefano Biffani, and Paola Crepaldi. 2022. "Using Pedigree and Genomic Data toward Better Management of Inbreeding in Italian Dairy Sheep and Goat Breeds" Animals 12, no. 20: 2828. https://doi.org/10.3390/ani12202828