
Extracellular Vesicles in Veterinary Medicine
 , , , and
, , , and 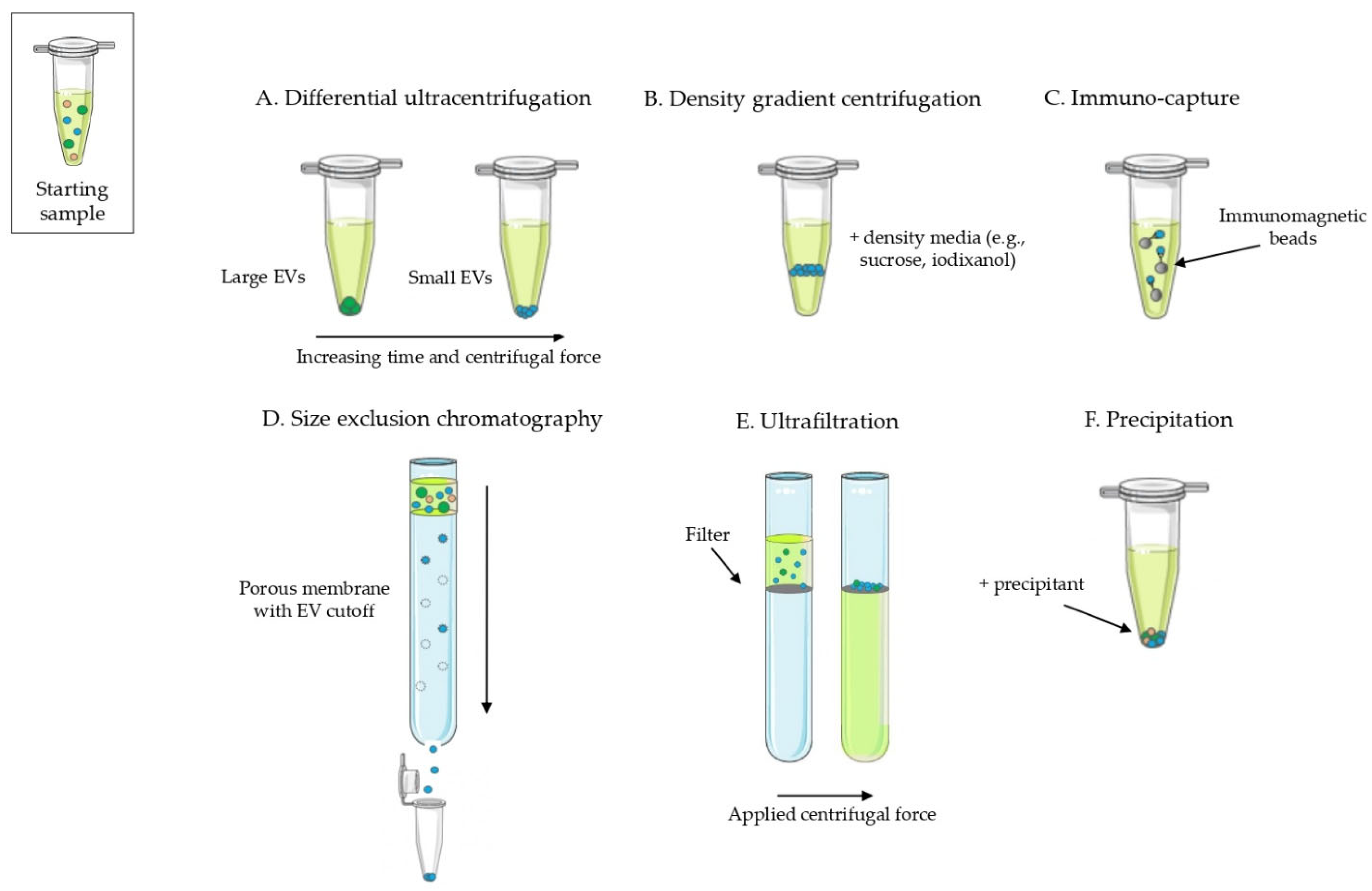{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Critical Points of EV Research in Veterinary Medicine
- Lack of standardization on EV isolation methods, often use of a single method: in the 80% of the reviewed papers a single technique is applied to isolate EVs, with only 20% of papers combining at least two methods. UC is the most commonly used, both as a single or combined technique, followed by commercial EV isolation kits (Figure 5). In addition, limitations of methods and controls are often not properly discussed or included.
- Lack of a constant, proper EV characterization: EVs are characterized in about 85% of the studies, and confirmation of the EV presence with at least two different methods is reported in 64% (Figure 6a). Several methods are used among which standard electron microscopy (EM) (32%), Western blot analysis (WB) (27%), and nanoparticle tracking analysis (18%) are the most commonly used (Figure 6b). However, in 15% of published papers (most of them published 2019–2021), no EV characterization is performed at all.
- Lack of a list of specific EV-related or -unrelated markers that can be used for EV characterization in different animal species;
- Lack of tissue- or cell-specific markers to isolate EV subpopulations;
- Lack of reference genes to be applied when EV-associated nucleic acids are investigated;
- Lack of species-specific pre-analytical indications. Often, veterinary scientists try to apply protocols and techniques derived from human (or sometimes lab animals) studies for the analysis of EVs in other animal species. However, there are several factors that can affect EV isolation and analysis, especially when body fluids are analyzed. The starting material (e.g., blood, saliva, urine, milk) can vary in composition depending on the animal species and this can imply differences in methods and protocols to analyze EVs.
- One internationally accepted (MISEV 2018) EV isolation/purification method;
- Minimum of two internationally accepted (MISEV 2018) methods for EV characterization.
3. EVs in Physiology and Pathophysiology
3.1. EVs in Immunity
3.2. EVs in Reproductive Physiology
3.3. Evs in Pathophysiology of Infectious Diseases
3.4. EVs in Environmental Adaptation and Nutrition
4. EVs as Biomarkers
4.1. EVs as Biomarkers in Reproduction
4.2. EVs as Biomarkers in Oncology
4.3. EVs as Biomarkers for Other Diseases
5. EVs in Therapy
6. EVs as Vaccines
7. EVs from Animal Milk for Human Applications
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minciacchi, V.R.; Freeman, M.R.; Vizio, D. Extracellular Vesicles in Cancer: Exosomes, Microvesicles and The Emerging Role of Large Oncosomes. Semin. Cell Dev. Biol. 2015, 40, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanwlani, R.; Fonseka, P.; Chitti, S.V.; Mathivan, S. Milk-Derived Extracellular Vesicles In Inter-Organism, Cross-Species Communication and Drug Delivery. Proteomes 2020, 8, 11. [Google Scholar] [CrossRef]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular Vesicles: Unique Intercellular Delivery Vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef] [Green Version]
- Teng, F.; Fussenegger, M. Shedding Light on Extracellular Vesicle Biogenesis and Bioengineering. Adv. Sci. 2021, 8, 2003505. [Google Scholar] [CrossRef]
- Van Niel, G.; Angelo, G.D.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Publ. Gr. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Coumans, F.A.W.; Brisson, A.R.; Buzas, E.I.; Dignat-george, F.; Drees, E.E.E.; El-andaloussi, S.; Emanueli, C.; Gasecka, A.; Hendrix, A.; Hill, A.F.; et al. Methodological Guidelines to Study Extracellular Vesicles. Circ. Res. 2017, 120, 1632–1648. [Google Scholar] [CrossRef]
- Couch, Y.; Buzàs, E.I.; Di Vizio, D.; Gho, Y.S.; Harrison, P.; Hill, A.F.; Lötvall, J.; Raposo, G.; Stahl, P.D.; Théry, C.; et al. A Brief History of Nearly EV-Erything—The Rise and Rise of Extracellular Vesicles. J. Extracell. Vesicles 2021, 10, e12144. [Google Scholar] [CrossRef]
- Pan, B.T.; Johnstone, R.M. Fate of the Transferrin Receptor during Maturation of Sheep Reticulocytes in Vitro: Selective Externalization of the Receptor. Cell 1983, 33, 967–978. [Google Scholar] [CrossRef]
- Harding, C.; Heuser, J.; Stahl, P. Receptor-Mediated Endocytosis of Transferrin and Recycling of the Transferrin Receptor in Rat Reticulocytes. J. Cell Biol. 1983, 97, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Haas-neill, S.; Forsythe, P. A Budding Relationship: Bacterial Extracellular Vesicles in the Microbiota–Gut–Brain Axis. Int. J. Mol. Sci. 2020, 21, 8899. [Google Scholar] [CrossRef] [PubMed]
- Tarashi, S.; Zamani, M.S.; Omrani, M.D.; Fateh, A.; Moshiri, A.; Saedisomeolia, A.; Siadat, S.D.; Kubow, S. Commensal and Pathogenic Bacterial-Derived Extracellular Vesicles in Host-Bacterial and Interbacterial Dialogues: Two Sides of the Same Coin. J. Immunol. Res. 2022, 2022, 8092170. [Google Scholar] [CrossRef] [PubMed]
- Askenase, P.W. Ancient Evolutionary Origin and Properties of Universally Produced Natural Exosomes Contribute to Their Therapeutic Superiority Compared to Artificial Nanoparticles. Int. J. Mol. Sci. 2021, 22, 1429. [Google Scholar] [CrossRef]
- Tomley, F.M.; Shirley, M.W. Livestock Infectious Diseases and Zoonoses. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2637–2642. [Google Scholar] [CrossRef] [Green Version]
- Kakkar, M.; Chauhan, A.S.; Bahl, T.; Rahi, M. Opportunities for One Health Policies to Reduce Poverty. Rev. Sci. Tech. 2019, 38, 135–144. [Google Scholar] [CrossRef]
- Amuasi, J.H.; Lucas, T.; Horton, R.; Winkler, A.S. Reconnecting for Our Future: The Lancet One Health Commission. Lancet 2020, 395, 1469–1471. [Google Scholar] [CrossRef]
- Bongiovanni, L.; Andriessen, A.; Wauben, M.H.M.; Hoen, E.N.M.N.T.; de Bruin, A. Extracellular Vesicles: Novel Opportunities to Understand and Detect Neoplastic Diseases. Vet. Pathol. 2021, 58, 453–471. [Google Scholar] [CrossRef]
- Diomaiuto, E.; Principe, V.; De Luca, A.; Laperuta, F.; Alterisio, C.; Di Loria, A. Exosomes in Dogs and Cats: An Innovative Approach to Neoplastic and Non-Neoplastic Diseases. Pharmaceuticals 2021, 14, 766. [Google Scholar] [CrossRef]
- Ong, S.L.; Blenkiron, C.; Haines, S.; Acevedo-Fani, A.; Leite, J.A.S.; Zempleni, J.; Anderson, R.C.; McCann, M.J. Ruminant Milk-Derived Extracellular Vesicles: A Nutritional and Therapeutic Opportunity? Nutrients 2021, 13, 2505. [Google Scholar] [CrossRef]
- Johnstone, R.M.; Adam, M.; Pan, B.T. The Fate of the Transferrin Receptor during Maturation of Sheep Reticulocytes in Vitro. Can. J. Biochem. Cell Biol. 1984, 62, 1246–1254. [Google Scholar] [CrossRef] [PubMed]
- Bazzan, E.; Tinè, M.; Casara, A.; Biondini, D.; Semenzato, U.; Cocconcelli, E.; Balestro, E.; Damin, M.; Radu, C.M.; Turato, G.; et al. Critical Review of the Evolution of Extracellular Vesicles’ Knowledge: From 1946 to Today. Int. J. Mol. Sci. 2021, 22, 6417. [Google Scholar] [CrossRef] [PubMed]
- Sammarco, A.; Finesso, G.; Cavicchioli, L.; Ferro, S.; Caicci, F.; Zanetti, R.; Sacchetto, R.; Zappulli, V. Preliminary Investigation of Extracellular Vesicles in Mammary Cancer of Dogs and Cats: Identification and Characterization. Vet. Comp. Oncol. 2018, 16, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Aguilera-Rojas, M.; Badewien-Rentzsch, B.; Plendl, J.; Kohn, B.; Einspanier, R. Exploration of Serum- and Cell Culture-Derived Exosomes from Dogs. BMC Vet. Res. 2018, 14, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klymiuk, M.C.; Balz, N.; Elashry, M.I.; Heimann, M.; Wenisch, S.; Arnhold, S. Exosomes Isolation and Identification from Equine Mesenchymal Stem Cells. BMC Vet. Res. 2019, 15, 42. [Google Scholar] [CrossRef] [Green Version]
- Decaro, N.; Buonavoglia, C. Canine Coronavirus: Not Only an Enteric Pathogen. Vet. Clin. North Am. Small Anim. Pract. 2011, 41, 1121–1132. [Google Scholar] [CrossRef]
- Li, D.; Luo, H.; Ruan, H.; Chen, Z.; Chen, S.; Wang, B.; Xie, Y. Isolation and Identification of Exosomes from Feline Plasma, Urine and Adipose- Derived Mesenchymal Stem Cells. BMC Vet. Res. 2021, 17, 272. [Google Scholar]
- Almiñana, C.; Vegas, A.R.; Tekin, M.; Hassan, M.; Uzbekov, R.; Fröhlich, T.; Bollwein, H.; Bauersachs, S. Isolation and Characterization of Equine Uterine Extracellular Vesicles: A Comparative Methodological Study. Int. J. Mol. Sci. 2021, 22, 979. [Google Scholar] [CrossRef]
- Méchin, M.C.; Nachat, R.; Coudane, F.; Adoue, V.; Arnaud, J.; Serre, G.; Simon, M. La Désimination Ou Citrullination Une Modification Post-Traductionnelle Aux Multiples Facettes. Medecine/Sciences 2011, 27, 49–54. [Google Scholar] [CrossRef]
- Michael, C. What the Shark Immune System Can and Cannot Provide for the Expanding Design Landscape of Immunotherapy. Expert Opin. Drug Discov. 2014, 9, 725–739. [Google Scholar]
- Criscitiello, M.F.; Kraev, I.; Lange, S. Deiminated Proteins in Extracellular Vesicles and Plasma of Nurse Shark (Ginglymostoma cirratum)—Novel Insights into Shark Immunity. Fish Shellfish Immunol. 2019, 92, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Magnadóttir, B.; Kraev, I.; Guðmundsdóttir, S.; Dodds, A.W.; Lange, S. Extracellular Vesicles from Cod (Gadus morhua L.) Mucus Contain Innate Immune Factors and Deiminated Protein Cargo. Dev. Comp. Immunol. 2019, 99, 103397. [Google Scholar] [CrossRef] [PubMed]
- Merchant, M.E.; Mills, K.; Leger, N.; Jerkins, E.; Vliet, K.A.; McDaniel, N. Comparisons of Innate Immune Activity of All Known Living Crocodylian Species. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2006, 143, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Barksdale, S.M.; Hrifko, E.J.; van Hoek, M.L. Cathelicidin Antimicrobial Peptide from Alligator Mississippiensis Has Antibacterial Activity against Multi-Drug Resistant Acinetobacter Baumanii and Klebsiella Pneumoniae. Dev. Comp. Immunol. 2017, 70, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Criscitiello, M.F.; Kraev, I.; Petersen, L.H.; Lange, S. Deimination Protein Profiles in Alligator Mississippiensis Reveal Plasma and Extracellular Vesicle-Specific Signatures Relating to Immunity, Metabolic Function, and Gene Regulation. Front. Immunol. 2020, 11, 651. [Google Scholar] [CrossRef]
- Tsagkogeorga, G.; McGowen, M.R.; Davies, K.T.J.; Jarman, S.; Polanowski, A.; Bertelsen, M.F.; Rossiter, S.J. A Phylogenomic Analysis of the Role and Timing of Molecular Adaptation in the Aquatic Transition of Cetartiodactyl Mammals. R. Soc. Open Sci. 2015, 2, 150156. [Google Scholar] [CrossRef] [Green Version]
- Tian, R.; Wang, Z.; Niu, X.; Zhou, K.; Xu, S.; Yang, G. Evolutionary Genetics of Hypoxia Tolerance in Cetaceans during Diving. Genome Biol. Evol. 2016, 8, 827–839. [Google Scholar] [CrossRef] [Green Version]
- Seluanov, A.; Gladyshev, V.N.; Vijg, J.; Gorbunova, V. Mechanisms of Cancer Resistance in Long-Lived Mammals. Nat. Rev. Cancer 2018, 18, 433–441. [Google Scholar] [CrossRef]
- Siming, M.; Vadim, N.G. Molecular Signature of Longevity: Insights from Cross-Species Comparative Studies. Semin. Cell Dev. Bio. 2017, 70, 190–203. [Google Scholar] [CrossRef]
- Wunschmann, A.; Siebert, U.; Frese, K.; Weiss, R.; Lockyer, C.; Heide-Jorgensen, G.M.; Baumgartner, W. Evidence of Infectious Disease in Harbour Porpoises (Phocoena Phocoena) Hunted in the Waters of Greenland and by-Caught in the German North Sea and Baltic Sea. Vet. Rec. 2011, 148, 715–720. [Google Scholar] [CrossRef]
- Siebert, U.; Prenger-Berninghoff, E.; Weiss, R. Regional Differences in Bacterial Flora in Harbour Porpoises from the North Atlantic: Environmental Effects? J. Appl. Microbiol. 2009, 106, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Magnadóttir, B.; Uysal-Onganer, P.; Kraev, I.; Svansson, V.; Skírnisson, K.; Lange, S. Deiminated Proteins and Extracellular Vesicles as Novel Biomarkers in Pinnipeds: Grey Seal (Halichoerus gryptus) and Harbour Seal (Phoca vitulina). Biochimie 2020, 171–172, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Magnadóttir, B.; Uysal-Onganer, P.; Kraev, I.; Svansson, V.; Hayes, P.; Lange, S. Deiminated Proteins and Extracellular Vesicles—Novel Serum Biomarkers in Whales and Orca. Comp. Biochem. Physiol. Part D Genomics Proteomics 2020, 34, 100676. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.A.; Kraev, I.; Lange, S. Protein Deimination and Extracellular Vesicle Profiles in Antarctic Seabirds. Biology 2020, 9, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidarimath, M.; Khalaj, K.; Kridli, R.T.; Kan, F.W.K.; Koti, M. Extracellular Vesicle Mediated Intercellular Communication at the Porcine Maternal-Fetal Interface: A New Paradigm for Conceptus- Endometrial Cross-Talk. Nat. Publ. Gr. 2017, 7, 40476. [Google Scholar] [CrossRef] [Green Version]
- Matsuno, Y.; Onuma, A.; Fujioka, Y.A.; Yasuhara, K.; Fujii, W.; Naito, K.; Sugiura, K. Effects of Exosome-like Vesicles on Cumulus Expansion in Pigs in Vitro. J. Reprod. Dev. 2017, 63, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Matsuno, Y.; Kanke, T.; Maruyama, N.; Fujii, W.; Naito, K.; Sugiura, K. Characterization of MRNA Profiles of the Exosome-like Vesicles in Porcine Follicular Fluid. PLoS ONE 2019, 14, e0217760. [Google Scholar] [CrossRef] [Green Version]
- Grzesiak, M.; Popiolek, K.; Knapczyk-Stwora, K. Extracellular Vesicles in Follicular Fluid of Sexually Mature Gilts’ Ovarian Antral Follicles—Identification and Proteomic Analysis. J. Physiol. Pharmacol. 2020, 71, 137–143. [Google Scholar] [CrossRef]
- Gatien, J.; Mermillod, P.; Tsikis, G.; Bernardi, O.; Idrissi, S.J.; Uzbekov, R.; Le Bourhis, D.; Salvetti, P.; Almiñana, C.; Saint-Dizier, M. Metabolomic Profile of Oviductal Extracellular Vesicles across the Estrous Cycle in Cattle. Int. J. Mol. Sci. 2019, 20, 6339. [Google Scholar] [CrossRef] [Green Version]
- Franchi, A.; Moreno-Irusta, A.; Domínguez, E.M.; Adre, A.J.; Giojalas, L.C. Extracellular Vesicles from Oviductal Isthmus and Ampulla Stimulate the Induced Acrosome Reaction and Signaling Events Associated with Capacitation in Bovine Spermatozoa. J. Cell. Biochem. 2020, 121, 2877–2888. [Google Scholar] [CrossRef]
- Lopera-Vasquez, R.; Hamdi, M.; Maillo, V.; Gutierrez-Adan, A.; Bermejo-Alvarez, P.; Angel Ramirez, M.; Yanez-Mo, M.; Rizos, D. Effect of Bovine Oviductal Extracellular Vesicles on Embryo Development and Quality in Vitro. Reproduction 2017, 153, 461–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavani, K.C.; Hendrix, A.; Van Den Broeck, W.; Couck, L.; Szymanska, K.; Lin, X.; De Koster, J.; Van Soom, A.; Leemans, B. Isolation and Characterization of Functionally Active Extracellular Vesicles from Culture Medium Conditioned by Bovine Embryos in Vitro. Int. J. Mol. Sci. 2019, 20, 38. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.M.; Viil, J.; Lättekivi, F.; Ord, J.; Reshi, Q.U.A.; Jääger, K.; Velthut-Meikas, A.; Andronowska, A.; Jaakma, Ü.; Salumets, A.; et al. Bovine Follicular Fluid and Extracellular Vesicles Derived from Follicular Fluid Alter the Bovine Oviductal Epithelial Cells Transcriptome. Int. J. Mol. Sci. 2020, 21, 5365. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Meng, K.; Wang, H.; Wang, Y.; Zhao, Y.; Kang, J.; Zhang, Y.; Quan, F. Identification of Small Extracellular Vesicle Subtypes in Follicular Fluid: Insights into the Function and MiRNA Profiles. J. Cell. Physiol. 2021, 236, 5633–5645. [Google Scholar] [CrossRef] [PubMed]
- Gebremedhn, S.; Gad, A.; Aglan, H.S.; Laurincik, J.; Prochazka, R.; Salilew-Wondim, D.; Hoelker, M.; Schellander, K.; Tesfaye, D. Extracellular Vesicles Shuttle Protective Messages against Heat Stress in Bovine granulosa Cells. Sci. Rep. 2020, 10, 15824. [Google Scholar] [CrossRef]
- Riou, C.; Brionne, A.; Cordeiro, L.; Harichaux, G.; Gargaros, A.; Labas, V.; Gautron, J.; Gérard, N. Avian Uterine Fluid Proteome: Exosomes and Biological Processes Potentially Involved in Sperm Survival. Mol. Reprod. Dev. 2020, 87, 454–470. [Google Scholar] [CrossRef]
- Cordeiro, L.; Lin, H.H.; Carvalho, A.V.; Uzbekov, R.; Blesbois, E.; Grasseau, I. First Insights on Seminal Extracellular Vesicles in Chickens of Contrasted Fertility. Reproduction 2021, 161, 489–498. [Google Scholar] [CrossRef]
- Bai, R.; Latifi, Z.; Kusama, K.; Nakamura, K.; Shimada, M.; Imakawa, K. Induction of Immune-Related Gene Expression by Seminal Exosomes in the Porcine Endometrium. Biochem. Biophys. Res. Commun. 2018, 495, 1094–1101. [Google Scholar] [CrossRef]
- Luvoni, G.C. Current Progress on Assisted Reproduction in Dogs and Cats: In Vitro Embryo Production. Reprod. Nutr. Dev. 2000, 40, 505–512. [Google Scholar] [CrossRef] [Green Version]
- de Almeida Monteiro Melo Ferraz, M.; Fujihara, M.; Nagashima, J.B.; Noonan, M.J.; Inoue-Murayama, M.; Songsasen, N. Follicular Extracellular Vesicles Enhance Meiotic Resumption of Domestic Cat Vitrified Oocytes. Sci. Rep. 2020, 10, 8619. [Google Scholar] [CrossRef]
- De Almeida Monteiro Melo Ferraz, M.; Nagashima, J.B.; Noonan, M.J.; Crosier, A.E.; Songsasen, N. Oviductal Extracellular Vesicles Improve Post-Thaw Sperm Function in Red Wolves and Cheetahs. Int. J. Mol. Sci. 2020, 21, 3733. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.H.; Badierah, R.; Redwan, E.M.; El-Fakharany, E.M. A Comprehensive Insight into the Role of Exosomes in Viral Infection: Dual Faces Bearing Different Functions. Pharmaceutics 2021, 13, 1045. [Google Scholar] [CrossRef] [PubMed]
- Schorey, J.S.; Cheng, Y.; Singh, P.P.; Smith, V.L. Exosomes and Other Extracellular Vesicles in Host–Pathogen Interactions. EMBO Rep. 2015, 16, 24–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcnamara, R.P.; Dittmer, D.P. ScienceDirect Extracellular Vesicles in Virus Infection and Pathogenesis. Curr. Opin. Virol. 2020, 44, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Nahand, J.S.; Mahjoubin-Tehran, M.; Moghoofei, M.; Pourhanifeh, M.H.; Mirzaei, H.R.; Asemi, Z.; Khatami, A.; Bokharaei-Salim, F.; Mirzaei, H.; Hamblin, M.R. Exosomal MiRNAs: Novel Players in Viral Infection. Epigenomics 2020, 12, 353–370. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liao, L.; Zhao, Q.; Zhang, X.; Li, H.; Lin, W.; Chen, F.; Xie, Q. MicroRNA Expression Profile in Extracellular Vesicles Derived from ALV-J Infected Chicken Semen. Virus Res. 2020, 286, 198083. [Google Scholar] [CrossRef]
- Changluan, Z.; Tan, L.; Sun, Y.; Qiu, X.; Lia, Y.; Song, C.; Liu, W.; Nair, V.; Ding, C. Exosomes Carry MicroRNAs into Neighboring Cells to Promote Diffusive Infection of Newcastle Disease Virus. Viruses 2019, 11, 527. [Google Scholar]
- Mao, L.; Liang, P.; Li, W.; Zhang, S.; Liu, M.; Yang, L.; Li, J.; Li, H.; Hao, F.; Sun, M.; et al. Exosomes Promote Caprine Parainfluenza Virus Type 3 Infection by Inhibiting Autophagy. J. Gen. Virol. 2020, 101, 717–734. [Google Scholar] [CrossRef]
- Sadeghipour, S.; Mathias, R.A. Herpesviruses Hijack Host Exosomes for Viral Pathogenesis. Semin. Cell Dev. Biol. 2017, 67, 91–100. [Google Scholar] [CrossRef]
- Patters, B.J.; Kumar, S. The Role of Exosomal Transport of Viral Agents in Persistent HIV Pathogenesis. Retrovirology 2018, 15, 79. [Google Scholar] [CrossRef] [Green Version]
- Carossino, M.; Dini, P.; Kalbfleisch, T.S.; Loynachan, A.T.; Canisso, I.F.; Shuck, K.M.; Timoney, P.J.; Cook, R.F.; Balasuriya, U.B.R. Downregulation of MicroRNA Eca-Mir-128 in Seminal Exosomes and Enhanced Expression of CXCL16 in the Stallion Reproductive Tract Are Associated with Long-Term Persistence of Equine Arteritis Virus. J. Virol. 2018, 92, e00015-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burny, A.; Cleuter, Y.; Kettmann, R.; Mammerickx, M.; Marbaix, G.; Portetelle, D.; Van Den Broeke, A.; Willems, L.; Thomas, R. Bovine Leukaemia: Facts and Hypotheses Derived from the Study of an Infectious Cancer. Vet. Microbiol. 1988, 17, 197–218. [Google Scholar] [CrossRef]
- Meas, S.; Usui, T.; Ohashi, K.; Sugimoto, C.; Onuma, M. Vertical Transmission of Bovine Leukemia Virus and Bovine Immunodeficiency Virus in Dairy Cattle Herds. Vet. Microbiol. 2002, 84, 275–282. [Google Scholar] [CrossRef]
- Yamada, T.; Shigemura, H.; Ishiguro, N.; Inoshima, Y. Cell Infectivity in Relation to Bovine Leukemia Virus Gp51 and P24 in Bovine Milk Exosomes. PLoS ONE 2013, 8, e77359. [Google Scholar] [CrossRef] [PubMed]
- Montaner-Tarbes, S.; Pujo, M.; Jabbar, T.; Hawes, P.; Chapman, D.; Del Portillo, H.; Fraile, L.; Sánchez-Cordón, P.J.; Dixon, L.; Montoya, M. Serum-Derived Extracellular Vesicles from African Swine Fever Virus-Infected Pigs Selectively Recruit Viral and Porcine Proteins. Viruses 2019, 11, 882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montaner-Tarbes, S.; Novell, E.; Tarancón, V.; Borrás, F.E.; Montoya, M.; Fraile, L.; del Portillo, H.A. Targeted-Pig Trial on Safety and Immunogenicity of Serum-Derived Extracellular Vesicles Enriched Fractions Obtained from Porcine Respiratory and Reproductive Virus Infections. Sci. Rep. 2018, 8, 17487. [Google Scholar] [CrossRef] [Green Version]
- Llanos, A.J.; Riquelme, R.A.; Sanhueza, E.M.; Hanson, M.A.; Blanco, C.E.; Parer, J.T.; Herrera, E.A.; Pulgar, V.M.; Reyes, R.V.; Cabello, G.; et al. The Fetal Llama versus the Fetal Sheep: Different Strategies to Withstand Hypoxia. High Alt. Med. Biol. 2004, 4, 193–202. [Google Scholar] [CrossRef]
- Dzal, Y.A.; Seow, A.; Borecky, L.G.; Chung, D.; Gill, S.K.G.; Milsom, W.K.; Pamenter, M.E. Glutamatergic Receptors Modulate Normoxic but Not Hypoxic Ventilation and Metabolism in Naked Mole Rats. Front. Physiol. 2019, 10, 106. [Google Scholar] [CrossRef] [Green Version]
- Dzal, Y.A.; Jenkin, S.E.M.; Lague, S.L.; Reichert, M.N.; York, J.M.; Pamenter, M.E. Oxygen in Demand: How Oxygen Has Shaped Vertebrate Physiology. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2015, 186, 4–26. [Google Scholar] [CrossRef]
- Munro, D.; Baldy, C.; Pamenter, M.E.; Treberg, J.R. The Exceptional Longevity of the Naked Mole-Rat May Be Explained by Mitochondrial Antioxidant Defenses. Aging Cell 2019, 18, e12916. [Google Scholar] [CrossRef] [Green Version]
- Woodley, R.; Buffenstein, R. Thermogenic Changes with Chronic Cold Exposure in the Naked Mole-Rat (Heterocephalus Glaber). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 133, 827–834. [Google Scholar] [CrossRef]
- Tian, X.; Azpurua, J.; Hine, C.; Vaidya, A.; Myakishev-Rempel, M.; Ablaeva, J.; Mao, Z.; Nevo, E.; Gorbunova, V.; Seluanov, A. High Molecular Weight Hyaluronan Mediates the Cancer Resistance of the Naked Mole-Rat. Nature 2013, 499, 346–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pamenter, M.E.; Uysal-Onganer, P.; Huynh, K.W.; Kraev, I.; Lange, S. Post-Translational Deimination of Immunological and Metabolic Protein Markers in Plasma and Extracellular Vesicles of Naked Mole-Rat (Heterocephalus glaber). Int. J. Mol. Sci. 2019, 20, 5378. [Google Scholar] [CrossRef]
- Wu, H.; Guang, X.; Al-Fageeh, M.B.; Cao, J.; Pan, S.; Zhou, H.; Zhang, L.; Abutarboush, M.H.; Xing, Y.; Xie, Z.; et al. Camelid Genomes Reveal Evolution and Adaptation to Desert Environments. Nat. Commun. 2014, 5, 5188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurgens, K. Strategies of Adaptation of Oxygen Transport Systems in Mammals to Life at High Altitude. Naturwissenschaften 1989, 76, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Criscitiello, M.F.; Kraev, I.; Lange, S. Deiminated Proteins in Extracellular Vesicles and Serum of Llama (Lama Glama)—Novel Insights into Camelid Immunity. Mol. Immunol. 2020, 117, 37–53. [Google Scholar] [CrossRef]
- Muyldermans, S. Nanobodies: Natural Single-Domain Antibodies. Annu. Rev. Biochem. 2013, 82, 775–797. [Google Scholar] [CrossRef] [Green Version]
- Henry, K.A.; van Faassen, H.; Harcus, D.; Marcil, A.; Hill, J.J.; Muyldermans, S.; MacKenzie, C.R. Llama Peripheral B-Cell Populations Producing Conventional and Heavy Chain-Only IgG Subtypes Are Phenotypically Indistinguishable but Immunogenetically Distinct. Immunogenetics 2019, 71, 307–320. [Google Scholar] [CrossRef]
- Carregari, V.C.; Rosa-Fernandes, L.; Baldasso, P.; Bydlowski, S.P.; Marangoni, S.; Larsen, M.R.; Palmisano, G. Snake Venom Extracellular Vesicles (SVEVs) Reveal Wide Molecular and Functional Proteome Diversity. Sci. Rep. 2018, 8, 12067. [Google Scholar] [CrossRef] [Green Version]
- Sanwlani, R.; Fonseka, P.; Mathivanan, S. Are Dietary Extracellular Vesicles Bioavailable and Functional in Consuming Organisms? Subcell Biochem. 2021, 97, 509–521. [Google Scholar]
- Kirchner, B.; Buschmann, D.; Paul, V.; Pfaffl, M.W. Postprandial Transfer of Colostral Extracellular Vesicles and Their Protein and MiRNA Cargo in Neonatal Calves. PLoS ONE 2020, 15, e0229606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Feng, S.; Wang, X.; Qazi, I.H.; Long, K.; Luo, Y.; Li, G.; Ning, C.; Wang, Y.; Hu, S.; et al. Exploration of Exosomal MicroRNA Expression Profiles in Pigeon ‘Milk’ during the Lactation Period. BMC Genomics 2018, 19, 828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, S.; Xiong, B.; Nan, X.; Wang, K.; Jiang, L.; Junhu, Y. Different Diets Change the Expression of Bovine Serum Extracellular Vesicle-MiRNAs. Animals 2019, 9, 1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulet, G.; Massias, J.; Taly, V. Liquid Biopsy: General Concepts. Acta Cytol. 2019, 63, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Yekula, A.; Muralidharan, K.; Kang, K.M.; Wang, L.; Balaj, L.; Carter, B.S. From Laboratory to Clinic: Translation of Extracellular Vesicle Based Cancer Biomarkers. Methods 2020, 177, 58–66. [Google Scholar] [CrossRef]
- Narita, M.; Nishida, H.; Asahina, R.; Nakata, K.; Yano, H.; Ueda, T.; Inden, M.; Akiyoshi, H.; Maeda, S.; Kamishina, H. Identification of Reference Genes of Extracellular Vesicles Isolated from Plasma Samples Oh Healthy Dogs by Ultracentrifugation, Precipitation and Membrane Affinity Chromatography Methods. Am. J. Vet. Res. 2019, 80, 449–454. [Google Scholar] [CrossRef]
- Hua, R.; Wang, Y.; Lian, W.; Li, W.; Xi, Y.; Xue, S.; Kang, T.; Lei, M. Small RNA-Seq Analysis of Extracellular Vesicles from Porcine Uterine Flushing Fluids during Peri-Implantation. Gene 2021, 766, 145117. [Google Scholar] [CrossRef]
- Zhou, C.; Cai, G.; Meng, F.; Xu, Z.; He, Y.; Hu, Q.; Zheng, E.; Huang, S.; Xu, Z.; Gu, T.; et al. Deep-Sequencing Identification of MicroRNA Biomarkers in Serum Exosomes for Early Pig Pregnancy. Front. Genet. 2020, 11, 536. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, H.; Zhang, R.; Tian, H.; Cheng, L.; Cheng, W.; Zhao, Y. Screening and Identification of MicroRNAs from Plasma-Derived Extracellular Vesicles (EVs) of Dazu Black Goat (Capra hircus) in Early Pregnant Stages. Gene 2021, 790, 145706. [Google Scholar] [CrossRef]
- Mellisho, E.A.; Velásquez, A.E.; Nuñez, M.J.; Cabezas, J.G.; Cueto, J.A.; Fader, C.; Castro, F.O.; Rodríguez-Álvarez, L. Identification and Characteristics of Extracellular Vesicles from Bovine Blastocysts Produced in Vitro. PLoS ONE 2017, 12, e0178306. [Google Scholar] [CrossRef] [Green Version]
- Melo-Baez, B.; Wong, Y.S.; Aguilera, C.J.; Cabezas, J.; Mançanares, A.C.F.; Riadi, G.; Castro, F.O.; Rodriguez-Alvarez, L. Micrornas from Extracellular Vesicles Secreted by Bovine Embryos as Early Biomarkers of Developmental Competence. Int. J. Mol. Sci. 2020, 21, 8888. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Silva, A.C.; Zhang, W.; Rutigliano, H.; Zhou, A. Raman Spectroscopy Characterization Extracellular Vesicles from Bovine Placenta and Peripheral Blood Mononuclear Cells. PLoS ONE 2020, 15, e0235214. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, K.; Stiebing, C.; Matthaus, C.; Schmitt, M.; Popp, J. Advantages and Limitations of Raman Spectroscopy for Molecular Diagnostics: An Update. Expert Rev. Mol. Diagn. 2015, 15, 773–787. [Google Scholar] [CrossRef]
- Alzhrani, G.N.; Alanazi, S.T.; Alsharif, S.Y.; Albalawi, A.M.; Alsharif, A.A.; Abdel-Maksoud, M.S.; Elsherbiny, N. Exosomes: Isolation, Characterization, and Biomedical Applications. Cell Biol. Int. 2021, 45, 1807–1831. [Google Scholar] [CrossRef] [PubMed]
- Howard, J.; Wyse, C.; Argyle, D.; Quinn, C.; Kelly, P.; McCann, A. Exosomes as Biomarkers of Human and Feline Mammary Tumours; A Comparative Medicine Approach to Unravelling the Aggressiveness of TNBC. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188431. [Google Scholar] [CrossRef] [PubMed]
- Sabbagh, Q.; Andre-Gregoire, G.; Guevel, L.; Gavard, J. Vesiclemia: Counting on Extracellular Vesicles for Glioblastoma Patients. Oncogene 2020, 39, 6043–6052. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, Q.; Zhang, J.; Li, C.; Miao, Y.R.; Lei, Q.; Li, Q.; Guo, A.Y. EVmiRNA: A Database of MiRNA Profiling in Extracellular Vesicles. Nucleic Acids Res. 2019, 47, D89–D93. [Google Scholar] [CrossRef] [Green Version]
- De Miguel Pérez, D.; Rodriguez Martínez, A.; Ortigosa Palomo, A.; Delgado Ureña, M.; Garcia Puche, J.L.; Robles Remacho, A.; Exposito Hernandez, J.; Lorente Acosta, J.A.; Ortega Sánchez, F.G.; Serrano, M.J. Extracellular Vesicle-MiRNAs as Liquid Biopsy Biomarkers for Disease Identification and Prognosis in Metastatic Colorectal Cancer Patients. Sci. Rep. 2020, 10, 3974. [Google Scholar] [CrossRef] [Green Version]
- Ozawa, P.M.M.; Jucoski, T.S.; Vieira, E.; Carvalho, T.M.; Malheiros, D.; Ribeiro, E.M.D.S.F. Liquid Biopsy for Breast Cancer Using Extracellular Vesicles and Cell-Free MicroRNAs as Biomarkers. Transl. Res. 2020, 223, 40–60. [Google Scholar] [CrossRef]
- Fish, E.J.; Irizarry, K.J.; Deinnocentes, P.; Ellis, C.J.; Prasad, N.; Moss, A.G.; Bird, R.C. Malignant Canine Mammary Epithelial Cells Shed Exosomes Containing Differentially Expressed MicroRNA That Regulate Oncogenic Networks. BMC Cancer 2018, 18, 832. [Google Scholar] [CrossRef]
- Narita, M.; Nishida, H.; Asahina, R.; Nakata, K.; Yano, H.; Dickinson, P.J.; Tanaka, T.; Akiyoshi, H.; Maeda, S.; Kamishina, H. Expression of Micrornas in Plasma and in Extracellular Vesicles Derived from Plasma for Dogs with Glioma and Dogs with Other Brain Diseases. Am. J. Vet. Res. 2020, 81, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.Z.; Ruan, J.S.; Jiang, Z.S.; Wang, L.; Wang, S.M. Extracellular Vesicles: A New Perspective in Tumor Therapy. Biomed. Res. Int. 2018, 2018, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Liu, C.; Bi, Z.Y.; Zhou, Q.; Zhang, H.; Li, L.L.; Zhang, J.; Zhu, W.; Song, Y.Y.Y.; Zhang, F.; et al. Comprehensive Landscape of Extracellular Vesicle-Derived RNAs in Cancer Initiation, Progression, Metastasis and Cancer Immunology. Mol. Cancer 2020, 19, 102. [Google Scholar] [CrossRef] [PubMed]
- Garnica, T.K.; Lesbon, J.C.C.; Ávila, A.C.F.C.M.; Rochetti, A.L.; Matiz, O.R.S.; Ribeiro, R.C.S.; Zoppa, A.; Nishiya, A.T.; Costa, M.T.; de Nardi, A.B.; et al. Liquid Biopsy Based on Small Extracellular Vesicles Predicts Chemotherapy Response of Canine Multicentric Lymphomas. Sci. Rep. 2020, 10, 20371. [Google Scholar] [CrossRef]
- Masaoutis, C.; Mihailidou, C.; Tsourouflis, G.; Theocharis, S. Exosomes in Lung Cancer Diagnosis and Treatment. From the Translating Research into Future Clinical Practice. Biochimie 2018, 151, 27–36. [Google Scholar] [CrossRef]
- Campos, A.; Sharma, S.; Obermair, A.; Salomon, C. Extracellular Vesicle-Associated MiRNAs and Chemoresistance: A Systematic Review. Cancers 2021, 13, 4608. [Google Scholar] [CrossRef]
- Asada, H.; Tomiyasu, H.; Uchikai, T.; Ishihara, G.; Goto-Koshino, Y.; Ohno, K.; Tsujimoto, H. Comprehensive Analysis of MiRNA and Protein Profiles within Exosomes Derived from Canine Lymphoid Tumour Cell Lines. PLoS ONE 2019, 14, e0208567. [Google Scholar] [CrossRef] [Green Version]
- Beaumier, A.; Robinson, S.R.; Robinson, N.; Lopez, K.E.; Meola, D.M.; Barber, L.G.; Bulmer, B.J.; Calvalido, J.; Rush, J.E.; Yeri, A.; et al. Extracellular Vesicular MicroRNAs as Potential Biomarker for Early Detection of Doxorubicin-Induced Cardiotoxicity. J. Vet. Intern. Med. 2020, 34, 1260–1271. [Google Scholar] [CrossRef] [Green Version]
- Ichii, O.; Ohta, H.; Horino, T.; Nakamura, T.; Hosotani, M.; Mizoguchi, T.; Morishita, K.; Nakamura, K.; Hoshino, Y.; Takagi, S.; et al. Urinary Exosome-Derived MicroRNAs Reflecting the Changes of Renal Function and Histopathology in Dogs. Sci. Rep. 2017, 7, 40340. [Google Scholar] [CrossRef] [Green Version]
- Pomatto, M.A.C.; Gai, C.; Bussolati, B.; Camussi, G. Extracellular Vesicles in Renal Pathophysiology. Front. Mol. Biosci. 2017, 4, 37. [Google Scholar] [CrossRef] [Green Version]
- Erdbrugger, U.; Le, T. Extracellular Vesicles as a Novel Diagnostic and Research Tool for Patients with HTN and Kidney Disease. Am. J. Ren. Physiol. 2019, 317, F641–F647. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.T.; Blostein, R.; Johnstone, R.M. Loss of the Transferrin Receptor during the Maturation of Sheep Reticulocytes in Vitro. An Immunological Approach. Biochem. J. 1983, 210, 37–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rout, E.D.; Webb, T.L.; Laurence, H.M.; Long, L.; Olver, C.S. Transferrin Receptor Expression in Serum Exosomes as a Marker of Regenerative Anaemia in the Horse. Equine Vet. J. 2015, 47, 101–106. [Google Scholar] [CrossRef]
- Martinez, C.R.; Santangelo, K.S.; Olver, C.S. Variability in the Cleavage of Exosome-Associated Transferrin Receptor Questions the Utility of Clinically Useful Soluble Transferrin Receptor Assays for Dogs, Cats, and Horses. Exp. Hematol. 2020, 86, 43.e1–52.e1. [Google Scholar] [CrossRef] [PubMed]
- van Weeren, P.R.; Back, W. Musculoskeletal Disease in Aged Horses and Its Management. Vet. Clin. North Am. Equine Pract. 2016, 32, 229–247. [Google Scholar] [CrossRef]
- Malda, J.; McIlwraith, C.W. Current Trends in Cartilage Science: An Impression from the ICRS World Conference 2012. Cartilage 2013, 4, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Boere, J.; Malda, J.; Van De Lest, C.H.A.; Van Weeren, P.R.; Wauben, M.H.M. Extracellular Vesicles in Joint Disease and Therapy. Front. Immunol. 2018, 9, 2575. [Google Scholar] [CrossRef] [Green Version]
- McIlwraith, C.W.; Frisbie, D.D.; Kawcak, C.E. The Horse as a Model of Naturally Occurring Osteoarthritis. Bone Joint Res. 2012, 1, 297–309. [Google Scholar] [CrossRef]
- Esa, A.; Connolly, K.D.; Williams, R.; Archer, C.W. Extracellular Vesicles in the Synovial Joint: Is There a Role in the Pathophysiology of Osteoarthritis? Malaysian Orthop. J. 2019, 13, 1–7. [Google Scholar] [CrossRef]
- Boere, J.; van de Lest, C.H.A.; Libregts, S.F.W.M.; Arkesteijn, G.J.A.; Geerts, W.J.C.; Nolte-’t Hoen, E.N.M.; Malda, J.; van Weeren, P.R.; Wauben, M.H.M. Synovial Fluid Pretreatment with Hyaluronidase Facilitates Isolation of CD44+ Extracellular Vesicles. J. Extracell. Vesicles 2016, 5, 31751. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.M.; An, J.H.; Yang, S.J.; Park, S.M.; Lee, J.H.; Chae, H.K.; Song, W.J.; Youn, H.Y. Influence of Canine Macrophage-Derived Extracellular Vesicles on Apoptosis in Canine Melanoma and Osteosarcoma Cell Lines. Anticancer Res. 2021, 41, 719–730. [Google Scholar] [CrossRef]
- Bacakova, L.; Zarubova, J.; Travnickova, M.; Musilkova, J.; Pajorova, J.; Slepicka, P.; Kasalkova, N.S.; Svorcik, V.; Kolska, Z.; Motarjemi, H.; et al. Stem Cells: Their Source, Potency and Use in Regenerative Therapies with Focus on Adipose-Derived Stem Cells—A Review. Biotechnol. Adv. 2018, 36, 1111–1126. [Google Scholar] [CrossRef] [PubMed]
- Keshtkar, S.; Azarpira, N.; Ghahremani, M.H. Mesenchymal Stem Cell-Derived Extracellular Vesicles: Novel Frontiers in Regenerative Medicine. Stem Cell Res. Ther. 2018, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Kornicka-Garbowska, K.; Pȩdziwiatr, R.; Woźniak, P.; Kucharczyk, K.; Marycz, K. Microvesicles Isolated from 5-Azacytidine-and-Resveratrol-Treated Mesenchymal Stem Cells for the Treatment of Suspensory Ligament Injury in Horse—A Case Report. Stem Cell Res. Ther. 2019, 10, 394. [Google Scholar] [CrossRef] [PubMed]
- Perrini, C.; Strillacci, M.G.; Bagnato, A.; Esposti, P.; Marini, M.G.; Corradetti, B.; Bizzaro, D.; Idda, A.; Ledda, S.; Capra, E.; et al. Microvesicles Secreted from Equine Amniotic-Derived Cells and Their Potential Role in Reducing Inflammation in Endometrial Cells in an in-Vitro Model. Stem Cell Res. Ther. 2016, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kuwahara, Y.; Yoshizaki, K.; Nishida, H.; Kamishina, H.; Maeda, S.; Takano, K.; Fujita, N.; Nishimura, R.; Jo, J.; Tabata, Y.; et al. Extracellular Vesicles Derived From Canine Mesenchymal Stromal Cells in Serum Free Culture Medium Have Anti-Inflammatory Effect on Microglial Cells. Front. Vet. Sci. 2021, 8, 633426. [Google Scholar] [CrossRef]
- An, J.H.; Li, Q.; Ryu, M.O.; Nam, A.R.; Bhang, D.H.; Jung, Y.C.; Song, W.J.; Youn, H.Y. TSG-6 in Extracellular Vesicles from Canine Mesenchymal Stem/Stromal Is a Major Factor in Relieving DSS-Induced Colitis. PLoS ONE 2020, 15, e0220756. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Liu, Q.; Reisdorf, R.L.; Boroumand, S.; Behfar, A.; Moran, S.L.; Amadio, P.C.; Gingery, A.; Zhao, C. Characterization of a Purified Exosome Product and Its Effects on Canine Flexor Tenocyte Biology. J. Orthop. Res. 2020, 38, 1845–1855. [Google Scholar] [CrossRef]
- McFarland, R.; Verthelyi, D.; Casey, W.; Arcinega, J.; Isbrucker, R.; Schmitt, M.; Finn, T.; Descamp, J.; Horiuchi, Y.; Sesardic, D.; et al. Non-Animal Replacement Methods for Human Vaccine Potency Testing: State of the Science and Future Directions. Procedia Vaccinol. 2011, 5, 16–32. [Google Scholar] [CrossRef] [Green Version]
- Morin, M.; Robinson, Y. Causes of Mystery Swine Disease. Can. Vet. J. 1992, 33, 6. [Google Scholar]
- Sarli, G.; D’annunzio, G.; Gobbo, F.; Benazzi, C.; Ostanello, F. The Role of Pathology in the Diagnosis of Swine Respiratory Disease. Vet. Sci. 2021, 8, 256. [Google Scholar] [CrossRef] [PubMed]
- Risser, J.; Ackerman, M.; Evelsizer, R.; Wu, S.; Kwon, B.; Hammer, J.M. Porcine Reproductive and Respiratory Syndrome Virus Genetic Variability a Management and Diagnostic Dilemma. Virol. J. 2021, 18, 206. [Google Scholar] [CrossRef] [PubMed]
- Montaner-Tarbes, S.; Borrás, F.E.; Montoya, M.; Fraile, L.; Del Portillo, H.A. Serum-Derived Exosomes from Non-Viremic Animals Previously Exposed to the Porcine Respiratory and Reproductive Virus Contain Antigenic Viral Proteins. Vet. Res. 2016, 47, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shivaramaiah, S.; Barta, J.; Hernandez-Velasco, X.; Tellez, G.; Hargis, B. Coccidiosis: Recent Advancements in the Immunobiology of Eimeria Species, Preventive Measures, and the Importance of Vaccination as a Control Tool against These Apicomplexan Parasites. Vet. Med. Res. Reports 2014, 5, 23–34. [Google Scholar] [CrossRef] [Green Version]
- del Cacho, E.; Gallego, M.; Lee, S.H.; Lillehoj, H.S.; Quilez, J.; Lillehoj, E.P.; Sánchez-Acedo, C. Induction of Protective Immunity against Eimeria Tenella, Eimeria Maxima, and Eimeria Acervulina Infections Using Dendritic Cell-Derived Exosomes. Infect. Immun. 2012, 80, 1909–1916. [Google Scholar] [CrossRef]
- Del Cacho, E.; Gallego, M.; Lillehoj, H.S.; Quilez, J.; Lillehoj, E.P.; Sánchez-Acedo, C. Induction of Protective Immunity against Experimental Eimeria Tenella Infection Using Serum Exosomes. Vet. Parasitol. 2016, 224, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Mehanny, M.; Lehr, C.M.; Fuhrmann, G. Extracellular Vesicles as Antigen Carriers for Novel Vaccination Avenues. Adv. Drug Deliv. Rev. 2021, 173, 164–180. [Google Scholar] [CrossRef]
- Nath Neerukonda, S.; Egan, N.A.; Patria, J.; Assakhi, I.; Tavlarides-Hontz, P.; Modla, S.; Muñoz, E.R.; Hudson, M.B.; Parcells, M.S. Comparison of Exosomes Purified via Ultracentrifugation (UC) and Total Exosome Isolation (TEI) Reagent from the Serum of Marek’s Disease Virus (MDV)-Vaccinated and Tumor-Bearing Chickens. J. Virol. Methods 2019, 263, 1–9. [Google Scholar] [CrossRef]
- Neerukonda, S.N.; Tavlarides-Hontz, P.; McCarthy, F.; Pendarvis, K.; Parcells, M.S. Comparison of the Transcriptomes and Proteomes of Serum Exosomes from Marek’s Disease Virus-Vaccinated and Protected and Lymphoma-Bearing Chickens. Genes 2019, 10, 116. [Google Scholar] [CrossRef] [Green Version]
- Admyre, C.; Johansson, S.M.; Qazi, K.R.; Filén, J.-J.; Lahesmaa, R.; Norman, M.; Neve, E.P.A.; Scheynius, A.; Gabrielsson, S. Exosomes with Immune Modulatory Features Are Present in Human Breast Milk. J. Immunol. 2007, 179, 1969–1978. [Google Scholar] [CrossRef] [Green Version]
- Kandimalla, R.; Aqil, F.; Tyagi, N.; Gupta, R. Milk Exosomes: A Biogenic Nanocarrier for Small Molecules and Macromolecules to Combat Cancer. Am. J. Reprod. Immunol. 2021, 85, e13349. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Inoshima, Y.; Matsuda, T.; Ishiguro, N. Comparison of Methods for Isolating Exosomes from Bovine Milk. J. Vet. Med. Sci. 2012, 74, 1523–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benmoussa, A.; Michel, S.; Gilbert, C.; Provost, P. Isolating Multiple Extracellular Vesicles Subsets, Including Exosomes and Membrane Vesicles, from Bovine Milk Using Sodium Citrate and Differential Ultracentrifugation. Bio-Protocol 2020, 10, e3636. [Google Scholar] [CrossRef] [PubMed]
- Blans, K.; Hansen, M.S.; Sørensen, L.V.; Hvam, M.L.; Howard, K.A.; Möller, A.; Wiking, L.; Larsen, L.B.; Rasmussen, J.T. Pellet-Free Isolation of Human and Bovine Milk Extracellular Vesicles by Size-Exclusion Chromatography. J. Extracell. Vesicles 2017, 6, 1294340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaswani, K.; Koh, Y.Q.; Almughlliq, F.B.; Peiris, H.N.; Mitchell, M.D. A Method for the Isolation and Enrichment of Purified Bovine Milk Exosomes. Reprod. Biol. 2017, 17, 341–348. [Google Scholar] [CrossRef]
- Vaswani, K.; Mitchell, M.D.; Holland, O.J.; Qin Koh, Y.; Hill, R.J.; Harb, T.; Davies, P.S.W.; Peiris, H. A Method for the Isolation of Exosomes from Human and Bovine Milk. J. Nutr. Metab. 2019, 2019, 5764740. [Google Scholar] [CrossRef]
- Rahman, M.M.; Shimizu, K.; Yamauchi, M.; Takase, H.; Ugawa, S.; Okada, A.; Inoshima, Y. Acidification Effects on Isolation of Extracellular Vesicles from Bovine Milk. PLoS ONE 2019, 14, e0222613. [Google Scholar] [CrossRef] [Green Version]
- Somiya, M.; Yoshioka, Y.; Ochiya, T. Biocompatibility of Highly Purified Bovine Milk-Derived Extracellular Vesicles. J. Extracell. Vesicles 2018, 7, 1440132. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, M.; Shimizu, K.; Rahman, M.; Ishikawa, H.; Takase, H.; Ugawa, S.; Okada, A.; Inoshima, Y. Efficient Method for Isolation of Exosomes from Raw Bovine Milk. Drug Dev. Ind. Pharm. 2019, 45, 359–364. [Google Scholar] [CrossRef]
- Hata, T.; Murakami, K.; Nakatani, H.; Yamamoto, Y.; Matsuda, T.; Aoki, N. Isolation of Bovine Milk-Derived Microvesicles Carrying MRNAs and MicroRNAs. Biochem. Biophys. Res. Commun. 2010, 396, 528–533. [Google Scholar] [CrossRef]
- Benmoussa, A.; Lee, C.H.C.; Laffont, B.; Savard, P.; Laugier, J.; Boilard, E.; Gilbert, C.; Fliss, I.; Provost, P. Commercial Dairy Cow Milk Micrornas Resist Digestion under Simulated Gastrointestinal Tract Conditions. J. Nutr. 2016, 146, 2206–2215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zempleni, J.; Aguilar-lozano, A.; Sadri, M.; Sukreet, S.; Manca, S.; Wu, D.; Zhou, F.; Mutai, E. Biological Activities of Extracellular Vesicles and Their Cargos from Bovine and Human Milk in Humans and Implications for Infants. J. Nutr. 2017, 147, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, B.; Kim, Y.; Park, D.J.; Oh, S. Comparative Analysis of Dietary Exosome-Derived MicroRNAs from Human, Bovine and Caprine Colostrum and Mature Milk. J. Anim. Sci. Technol. 2021, 63, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Zhou, B.; Ross, S.A.; Zempleni, J. Nutrition, MicroRNAs, and Human Health. Adv. Nutr. 2017, 8, 105–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Shi, Z.; Wang, X.; Mu, S.; Xu, X.; Shen, L.; Ping, L. Protective Effects of Bovine Milk Exosomes against Oxidative Stress in IEC-6 Cells. Eur. J. Nutr. 2021, 60, 317–327. [Google Scholar] [CrossRef]
- Tong, L.; Hao, H.; Zhang, X.; Zhang, Z.; Lv, Y.; Zhang, L.; Yi, H. Oral Administration of Bovine Milk-Derived Extracellular Vesicles Alters the Gut Microbiota and Enhances Intestinal Immunity in Mice. Mol. Nutr. Food Res. 2020, 64, e1901251. [Google Scholar] [CrossRef]
- Li, B.; Hock, A.; Wu, R.Y.; Minich, A.; Botts, S.R.; Lee, C.; Antounians, L.; Miyake, H.; Koike, Y.; Chen, Y.; et al. Bovine Milk-Derived Exosomes Enhance Goblet Cell Activity and Prevent the Development of Experimental Necrotizing Enterocolitis. PLoS ONE 2019, 14, e0211431. [Google Scholar] [CrossRef]
- Maghraby, M.K.; Li, B.; Chi, L.; Ling, C.; Benmoussa, A.; Provost, P.; Postmus, A.C.; Abdi, A.; Pierro, A.; Bourdon, C.; et al. Extracellular Vesicles Isolated from Milk Can Improve Gut Barrier Dysfunction Induced by Malnutrition. Sci. Rep. 2021, 11, 7635. [Google Scholar] [CrossRef]
- Tong, L.; Hao, H.; Zhang, Z.; Lv, Y.; Liang, X.; Liu, Q.; Liu, T.; Gong, P.; Zhang, L.; Cao, F.; et al. Milk-Derived Extracellular Vesicles Alleviate Ulcerative Colitis by Regulating the Gut Immunity and Reshaping the Gut Microbiota. Theranostics 2021, 11, 8570–8586. [Google Scholar] [CrossRef]
- Stremmel, W.; Weiskirchen, R.; Melnik, B.C. Milk Exosomes Prevent Intestinal Inflammation in a Genetic Mouse Model of Ulcerative Colitis: A Pilot Experiment. Inflamm. Intest. Dis. 2020, 5, 117–123. [Google Scholar] [CrossRef]
- Reif, S.; Elbaum-Shiff, Y.; Koroukhov, N.; Shilo, I.; Musseri, M.; Golan-Gerstl, R. Cow and Human Milk-Derived Exosomes Ameliorate Colitis in Dss Murine Model. Nutrients 2020, 12, 2589. [Google Scholar] [CrossRef] [PubMed]
- Thorning, T.K.; Raben, A.; Tholstrup, T.; Soedamah-Muthu, S.S.; Givens, I.; Astrup, A. Milk and Dairy Products: Good or Bad for Human Health? An Assessment of the Totality of Scientific Evidence. Food Nutr. 2016, 22, 32527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Go, G.; Jeon, J.; Lee, G.; Lee, J.H.; Lee, S.H. Bovine Milk Extracellular Vesicles Induce the Proliferation and Differentiation of Osteoblasts and Promote Osteogenesis in Rats. J. Food Biochem. 2021, 45, e13705. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.C.; Pieters, B.C.H.; Guimarães, P.B.; Duffles, L.F.; Heredia, J.E.; Silveira, A.L.M.; Oliveira, A.C.C.; Teixeira, M.M.; Ferreira, A.V.M.; Silva, T.A.; et al. Bovine Milk Extracellular Vesicles Are Osteoprotective by Increasing Osteocyte Numbers and Targeting RANKL/OPG System in Experimental Models of Bone Loss. Front. Bioeng. Biotechnol. 2020, 8, 891. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.; Maburutse, B.E.; Kang, M.; Park, M.R.; Park, D.J.; Kim, Y.; Oh, S. Short Communication: Dietary Bovine Milk–Derived Exosomes Improve Bone Health in an Osteoporosis-Induced Mouse Model. J. Dairy Sci. 2020, 103, 7752–7760. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C. Milk Exosomal MiRNAs: Potential Drivers of AMPK-To-MTORC1 Switching in β-Cell de-Differentiation of Type 2 Diabetes Mellitus. Nutr. Metab. 2019, 16, 85. [Google Scholar] [CrossRef] [PubMed]
- Fonseka, P.; Kang, T.; Chee, S.; Chitti, S.V.; Sanwlani, R.; Ang, C.S.; Mathivanan, S. Temporal Quantitative Proteomics Analysis of Neuroblastoma Cells Treated with Bovine Milk-Derived Extracellular Vesicles Highlights the Anti-Proliferative Properties of Milk-Derived Extracellular Vesicles. Cells 2021, 10, 750. [Google Scholar] [CrossRef]
- Mungala, R.; Aqil, F.; Jeyabalan, J.; Gupta, R.C. Bovine Milk-Derived Exosomes for Drug Delivery. Cancer Lett. 2016, 371, 48–61. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Yao, S.; Zhou, Z.; Shi, J.; Huang, Z.; Wu, Z. Hyaluronan Decoration of Milk Exosomes Directs Tumor-Specific Delivery of Doxorubicin. Carbohydr. Res. 2020, 493, 108032. [Google Scholar] [CrossRef]
- Warren, M.R.; Zhang, C.; Vedadghavami, A.; Bokvist, K.; Dhal, P.K.; Bajpayee, A.G. Milk Exosomes with Enhanced Mucus Penetrability for Oral Delivery SiRNA. Biomater Sci. 2021, 9, 4260–4277. [Google Scholar] [CrossRef]
- Cai, M.; He, H.; Jia, X.; Chen, S.; Wang, J.; Shi, Y.; Liu, B.; Xiao, W.; Lai, S. Genome-Wide MicroRNA Profiling of Bovine Milk-Derived Exosomes Infected with Staphylococcus aureus. Cell Stress Chaperones 2018, 23, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Mecocci, S.; Gevi, F.; Pietrucci, D.; Cavinato, L.; Luly, F.R.; Pascucci, L.; Petrini, S.; Ascenzioni, F.; Zolla, L.; Chillemi, G.; et al. Anti-Inflammatory Potential of Cow, Donkey and Goat Milk Extracellular Vesicles as Revealed by Metabolomic Profile. Nutrients 2020, 12, 2908. [Google Scholar] [CrossRef] [PubMed]
- Mecocci, S.; Pietrucci, D.; Milanesi, M.; Pascucci, L.; Filippi, S.; Rosato, V.; Chillemi, G.; Capomaccio, S.; Cappelli, K. Transcriptomic Characterization of Cow, Donkey and Goat Milk Extracellular Vesicles Reveals Their Anti-Inflammatory and Immunomodulatory Potential. Int. J. Mol. Sci. 2021, 22, 12759. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.N.; Hu, H.; Wen, P.C.; Lian, S.; Xie, X.L.; Song, H.L.; Yang, Z.N.; Ren, F.Z. Yak Milk–Derived Exosomes Alleviate Lipopolysaccharide-Induced Intestinal Inflammation by Inhibiting PI3K/AKT/C3 Pathway Activation. J. Dairy Sci. 2021, 104, 8411–8424. [Google Scholar] [CrossRef]
- Gao, H.N.; Ren, F.Z.; Wen, P.C.; Xie, L.X.; Wang, R.; Yang, Z.N.; Li, Y.X. Yak Milk–Derived Exosomal MicroRNAs Regulate Intestinal Epithelial Cells on Proliferation in Hypoxic Environment. J. Dairy Sci. 2021, 104, 1291–1303. [Google Scholar] [CrossRef]
- Xie, M.Y.; Chen, T.; Xi, Q.Y.; Hou, L.J.; Luo, J.Y.; Zeng, B.; Li, M.; Sun, J.J.; Zhang, Y.L. Porcine Milk Exosome MiRNAs Protect Intestinal Epithelial Cells against Deoxynivalenol-Induced Damage. Biochem. Pharmacol. 2020, 175, 113898. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moccia, V.; Sammarco, A.; Cavicchioli, L.; Castagnaro, M.; Bongiovanni, L.; Zappulli, V. Extracellular Vesicles in Veterinary Medicine. Animals 2022, 12, 2716. https://doi.org/10.3390/ani12192716
Moccia V, Sammarco A, Cavicchioli L, Castagnaro M, Bongiovanni L, Zappulli V. Extracellular Vesicles in Veterinary Medicine. Animals. 2022; 12(19):2716. https://doi.org/10.3390/ani12192716
Chicago/Turabian StyleMoccia, Valentina, Alessandro Sammarco, Laura Cavicchioli, Massimo Castagnaro, Laura Bongiovanni, and Valentina Zappulli. 2022. "Extracellular Vesicles in Veterinary Medicine" Animals 12, no. 19: 2716. https://doi.org/10.3390/ani12192716