
Genetic Analyses of Rabbit Survival and Individual Birth Weight
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals, Housing and Feeding
2.3. Genetic Analysis
3. Results
3.1. Descriptive Statistics
3.2. Heritabilities
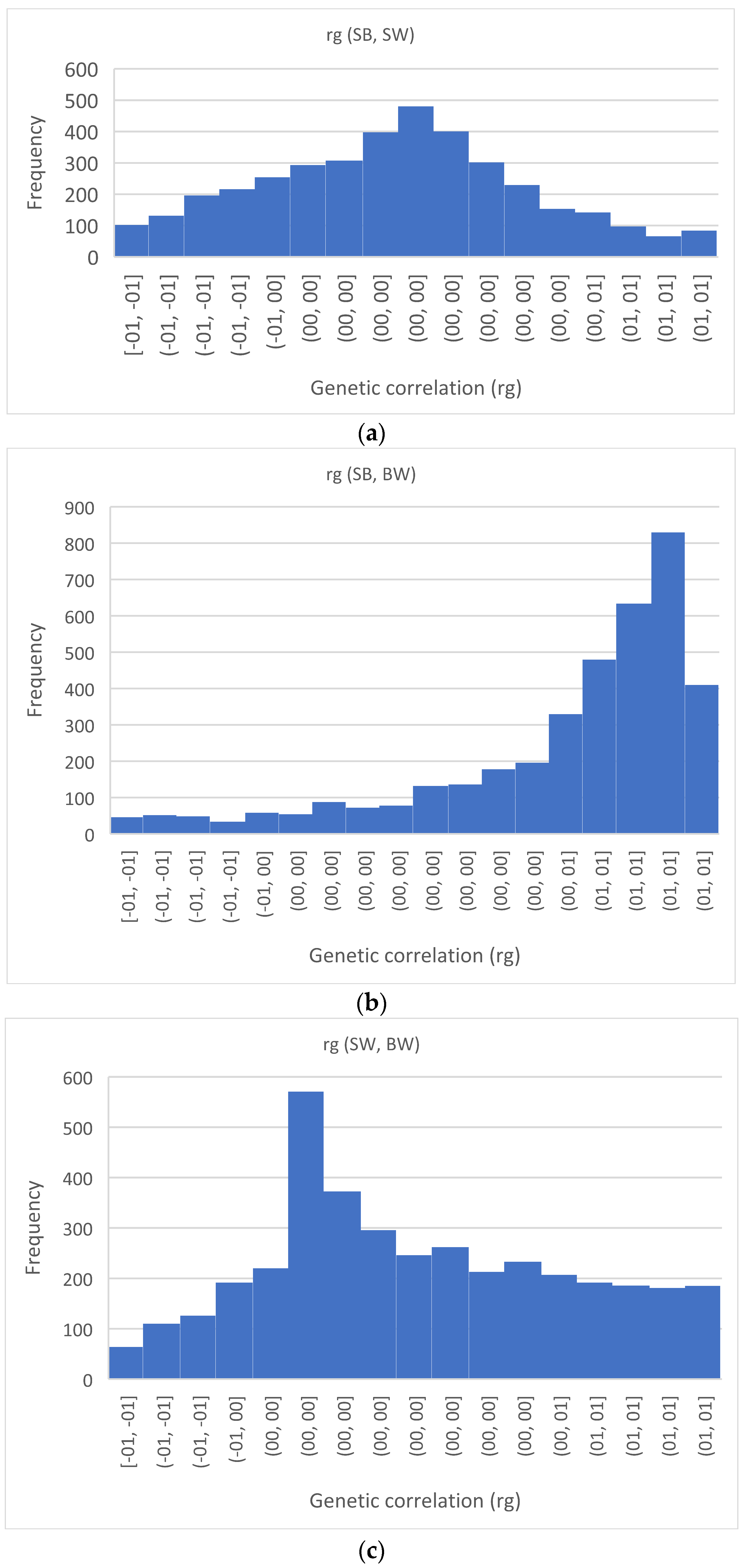
3.3. Genetic Correlations
3.4. Maternal Permanent Environmental and Common Litter Effects
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cartuche, L.; Pascual, M.; Gómez, E.A.; Blasco, A. Economic weights in rabbit meat production. World Rabbit Sci. 2014, 22, 165–177. [Google Scholar] [CrossRef]
- García, M.L.; Argente, M.J. The genetic improvement in meat rabbits. In Lagomorpha Charact; IntechOpen: London, UK, 2020; Volume 5, pp. 1–18. [Google Scholar]
- Szendrö, Z.; Cullere, M.; Atkári, T.; DalleZote, A. The birth weight of rabbits: Influencing factors and effect on behavioural, productive and reproductive traits: A review. Livestock Sci. 2019, 230, 1–10. [Google Scholar] [CrossRef]
- Argente, M.J.; Santacreu, M.A.; Climent, A.; Blasco, A. Relationships between uterine and fetal traits in rabbits selected on uterine capacity. J. Anim. Sci. 2003, 81, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Argente, M.J.; Santacreu, M.A.; Climent, A.; Blasco, A. Effects of intrauterine crowding on available uterine space per fetus in rabbits. Livest. Sci. 2008, 114, 211–219. [Google Scholar] [CrossRef]
- Roehe, R.; Shrestha, N.P.; Mekkawy, W.; Baxter, E.M.; Knap, P.W.; Smurthwaite, K.M.; Jarvis, S.; Lawrence, A.B.; Ed-wards, S.A. Genetic analyses of piglet survival and individual birth weight on first generation data of a selection experiment for piglet survival under outdoor conditions. Livest. Sci. 2009, 121, 173–181. [Google Scholar] [CrossRef]
- Blasco, A.; Bidanel, J.P.; Bolet, G.; Haley, C.S.; Santacreu, M.A. The genetics of prenatal survival of pigs and rabbits: A review. Livest. Prod. Sci. 1993, 37, 1–21. [Google Scholar] [CrossRef]
- Rothschild, M.F.; Bidanel, J.P. Biology and genetics of reproduction. In The Genetics of the Pig; Rothschild, M.F., Ruvinsky, A., Eds.; CAB Int.: Oxfordshire, UK, 1998; pp. 313–344. [Google Scholar]
- García, M.L.; Baselga, M. Estimation of genetic response to selection in litter size of rabbits using a cryopreserved control population. Livest. Prod. Sci. 2002, 74, 45–53. [Google Scholar] [CrossRef]
- Badawy, A.Y.; Peiró, R.; Blasco, A.; Santacreu, M.A. Correlated response on litter size traits and survival traits after two-stage selection for ovulation rate and litter size in rabbits. Animal 2019, 13, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, D.; Andersen, S.; Gianola, D.; Korsgaard, I. Bayesian inference in threshold models using Gibbs sampling. Genet. Sel. Evol. 1994, 27, 229–249. [Google Scholar] [CrossRef]
- Brun, J.M.; Baselga, M. Analysis of reproductive performances during the formation of a rabbit synthetic strain. In Proceedings of the World Rabbit Congress, Puebla, Mexico, 7–10 September 2004. [Google Scholar]
- Blumetto, O.; Olivas, I.; Torres, A.G.; Villagrá, A. Use of straw and wood shavings as nest material in primiparous does. World Rabbit Sci. 2010, 18, 237–242. [Google Scholar]
- Legarra, A.; Varona, L.; López de Maturana, E. Program TM Threshold Model. 2008. Available online: http://snp.toulouse.inra.fr/~alegarra/ (accessed on 17 August 2022).
- Sorensen, D.; Gianola, D. Likelihood, Bayesian, and MCMC Methods in Quantitative Genetics; Springer Science and Business Media: New York, NY, USA, 2002. [Google Scholar]
- Geyer, C.M. Practical Markov chain Monte Carlo. Stat. Sci. 1992, 7, 467–511. [Google Scholar] [CrossRef]
- Partridge, G.G.; Foley, S.; Corrigall, W. Reproductive performance in purebred and crossbred commercial rabbits. Anim. Prod. 1981, 32, 325–331. [Google Scholar] [CrossRef]
- Rashwan, A.A.; Marai, I.F.M. Mortalty in young rabbits: A review. World Rabbit Sci. 2000, 8, 111–124. [Google Scholar]
- Kadi, S.A.; Zerrouki, N.; Louchami, Y.; Dahmani, F. Pre-weaning mortality in rabbits of local Algerian population. In Proceedings of the 4th International Conferences on Rabbit Production in Hot Climate, Sharm El-Sheikh, Egypt, 24–25 February 2005. [Google Scholar]
- Van Arendonk, J.A.M.; van Rosmeulen, C.; Janss, L.L.G.; Knol, E.F. Estimation of direct and maternal genetic (co) variances for survival within litters of piglets. Livest. Prod. Sci. 1996, 46, 163–171. [Google Scholar] [CrossRef]
- Lund, S.M.; Puonti, M.; Rydhmer, L.; Jensen, J. Relationship between litter size and perinatal and preweaning survival in pigs. Anim. Sci. 2002, 74, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Knol, E.F.; Ducro, B.J.; Van Arendonk, J.A.M.; Van der Lende, T. Direct, maternal and nurse sow genetic effects on farrowing, pre-weaning and total piglet survival. Livest. Prod. Sci. 2002, 73, 153–164. [Google Scholar] [CrossRef]
- Su, G.; Sorensen, D.; Lund, M.S. Analysis of variance components for piglet survival using threshold model. In Proceedings of the 8th World Congress on Genetics Applied to Livestock Production, Belo Horizonte, Brazil, 13–18 August 2006. [Google Scholar]
- Kapell, D.N.; Ashworth, C.J.; Knap, P.W.; Roehe, R. Genetic parameters for piglet survival, litter size and birth weight or its variation within litter in sire and dam lines using Bayesian analysis. Livest. Sci. 2011, 135, 215–224. [Google Scholar] [CrossRef]
- Nguyen, T.Q.; Knap, P.W.; Simm, G.; Edwards, S.A.; Roehe, R. Evaluation of direct and maternal responses in reproduction traits based on different selection strategies for postnatal piglet survival in a selection experiment. Genet. Sel. Evol. 2021, 53, 28. [Google Scholar] [CrossRef]
- Knol, E.F.; Van der Lende, T.; Zak, L.J. Genetic aspects of piglet survival and related traits: A review. J. Anim. Sci. 2022, 100, 1–9. [Google Scholar] [CrossRef]
- Argente, M.J.; Santacreu, M.A.; Climent, A.; Blasco, A. Phenotypic and genetic parameters of birth weight and weaning weight of rabbits born from unilaterally ovariectomized and intact does. Livest. Prod. Sci. 1999, 57, 159–167. [Google Scholar] [CrossRef]
- Canario, L.; Cantoni, E.; Le Bihan, E.; Caritez, J.C.; Billon, Y.; Bidanel, J.P.; Foulley, J.L. Between-breed variability of stillbirth and its relationship with sow and piglet characteristics. J. Anim. Sci. 2006, 84, 3185–3196. [Google Scholar] [CrossRef] [PubMed]
- Roehe, R.; Kalm, E. Estimation of genetic and environmental risk factors associated with pre-weaning mortality in piglets using generalized linear mixed models. Anim. Sci. 2000, 70, 227–240. [Google Scholar] [CrossRef]


{kind=link}
| Temperature Outside (°C) | Temperature Inside (°C) | Relative Humidity Inside (%) | |||||
|---|---|---|---|---|---|---|---|
| Minimun | Maximum | Minimun | Maximum | Minimun | Maximum | ||
| Summer | June | 25 | 30 | 22 | 28 | 21 | 82 |
| July | 27 | 39 | 26 | 33 | 21 | 80 | |
| August | 30 | 35 | 28 | 36 | 23 | 80 | |
| Average | 27.3 | 34.7 | 25.3 | 32.3 | 21.7 | 80.7 | |
| Fall | September | 24 | 30 | 22 | 27 | 19 | 83 |
| October | 21 | 27 | 20 | 23 | 21 | 79 | |
| November | 16 | 21 | 19 | 23 | 20 | 68 | |
| Average | 20.3 | 26.0 | 20.3 | 24.3 | 20.0 | 76.7 | |
| Traits | N | Mean | SD |
|---|---|---|---|
| Individual birth weight (g) (with dead kits) | 1696 | 51.36 | 15.19 |
| Individual birth weight (g) (without dead kits) | 1536 | 52.79 | 14.14 |
| Individual birth weight of dead kits (g) | 160 | 37.67 | 17.93 |
| Number of kits born per litter | 208 | 8.15 | 3.16 |
| Number of kits born alive per litter | 208 | 7.37 | 2.96 |
| Number of kits weaned per litter | 208 | 6.59 | 2.41 |
| Survival at birth (%) a | 1536 | 90.56 | 23.03 |
| Survival at weaning (%) b | 1373 | 89.41 | 24.47 |
| Factors | Birth | Weaning | ||
|---|---|---|---|---|
| N | Weight (g) | N | Weight (g) | |
| Parity | ||||
| First | ||||
| Live | 617 | 46.84 | 467 | 48.1 |
| Dead | 61 | 35.33 | 150 | 42.9 |
| Second | ||||
| Live | 461 | 54.34 | 397 | 56.48 |
| Dead | 47 | 39.94 | 64 | 41.1 |
| Third | ||||
| Live | 458 | 59.25 | 395 | 61.37 |
| Dead | 52 | 37.21 | 63 | 45.97 |
| Lactation status | ||||
| Lactation | ||||
| Live | 881 | 49.8 | 690 | 51.51 |
| Dead | 80 | 35.8 | 191 | 43.58 |
| Non-lactation | ||||
| Live | 655 | 56.81 | 569 | 59.1 |
| Dead | 80 | 39.1 | 86 | 42.28 |
| Season of farrowing | ||||
| Summer | ||||
| Live | 945 | 49.22 | 753 | 58.81 |
| Dead | 97 | 37.45 | 192 | 42.97 |
| Autumn | ||||
| Live | 591 | 58.49 | 506 | 60.99 |
| Dead | 63 | 37.04 | 85 | 43.65 |
| Nest quality | ||||
| Bad | ||||
| Live | 20 | 52.5 | 17 | 51 |
| Dead | 3 | 51.33 | 3 | 61 |
| Intermediate | ||||
| Live | 79 | 52.45 | 62 | 54.43 |
| Dead | 10 | 47.9 | 17 | 45.23 |
| Excellent | ||||
| Live | 1437 | 52.84 | 1180 | 54.99 |
| Dead | 147 | 36.28 | 257 | 40.45 |
| Cannibalism | ||||
| Non | ||||
| Live | 1492 | 52.94 | 1222 | 55.1 |
| Dead | 145 | 37.99 | 270 | 43.2 |
| Yes | ||||
| Live | 44 | 47.52 | 37 | 48.54 |
| Dead | 15 | 30.53 | 7 | 42.14 |
| Place of birth in the cage | ||||
| Inside nest | ||||
| Live | 1467 | 52.49 | 1201 | 54.63 |
| Dead | 138 | 35.21 | 266 | 42.79 |
| Outside nest | ||||
| Live | 69 | 59.24 | 58 | 60.53 |
| Dead | 22 | 50.36 | 11 | 52.45 |
| Sex | ||||
| Male | ||||
| Live | 777 | 53.19 | 620 | 55.34 |
| Dead | 67 | 40 | 157 | 44.73 |
| Female | ||||
| Live | 759 | 52.38 | 639 | 54.48 |
| Dead | 93 | 35.34 | 120 | 41.15 |
| SB | SW | BW | |
|---|---|---|---|
| SB | 0.018 [0.001, 0.052] | −0.111 [−0.763, 0.613] P (rg < 0) = 0.67 | +0.134 [−0.953, 0.996] P (rg > 0) = 0.63 |
| SW | −0.008 [−0.099, 0.089] P (rp < 0) = 0.57 | 0.023 [0.002, 0.063] | +0.041 [−0.703, 0.935] P (rg > 0) = 0.47 |
| BW | +0.341 [0.260, 0.409] P (rp > 0) = 1.00 | 0.248 [0.162, 0.328] P (rp > 0) = 1.00 | 0.088 [0.006, 0.234] |
| SB | SW | BW | |
|---|---|---|---|
| SB | 0.021 [0.001, 0.057] | −0.072 [−0.744, 0.654] P (rg < 0) = 0.56 | +0.535 [−0.490, 0.944] P (rg > 0) = 0.87 |
| SW | −0.009 [−0.099, 0.096] P (rp < 0) = 0.58 | 0.027 [0.003, 0.071] | +0.083 [−0.628, 0.892] P (rg > 0) = 0.52 |
| BW | +0.355 [0.290, 0.419] P (rp > 0) = 1.00 | +0.228 [0.155, 0.305] P (rp > 0) = 1.00 | 0.146 [0.039, 0.292] |
| SB | SW | BW | |
|---|---|---|---|
| SB | 0.043 [0.003, 0.106] | −0.003 [−0.791, 0.727] P (rm < 0) = 0.50 | −0.437 [−0.958, 0.335] P(rm < 0) = 0.85 |
| SW | 0.024 [0.001, 0.071] | +0.098 [−0.923, 0.972] P(rm > 0) = 0.57 | |
| BW | 0.070 [0.007, 0,149] |
| SB | SW | BW | |
|---|---|---|---|
| SB | 0.050 [0.005, 0.114] | +0.064 [−0.632, 0.862] P (rm > 0) = 0.54 | −0.702 [−0.996, −0.001] P (rm < 0) = 0.95 |
| SW | 0.024 [0.001, 0.067] | −0.273 [−0.984, 0.657] P (rm < 0) = 0.68 | |
| BW | 0.037 [0.003, 0.093] |
| SB | SW | BW | |
|---|---|---|---|
| SB | 0.199 [0.127, 0.270] | −0.029 [−0.435, 0.390] P (rc < 0) = 0.55 | +0.632 [0.384, 0.834] P (rc > 0) = 0.99 |
| SW | 0.234 [0.161, 0.308] | +0.362 [0.111, 0.608] P (rc > 0) = 0.99 | |
| BW | 0.435 [0.351, 0.512] |
| SB | SW | BW | |
|---|---|---|---|
| SB | 0.195 [0.125, 0.268] | −0.063 [−0.434, 0.454] P (rc < 0) = 0.59 | +0.768 [0.541, 0.962] P (rc > 0) = 1.00 |
| SW | 0.225 [0.158, 0.298] | +0.312 [0.041, 0.596] P (rc > 0) = 0.97 | |
| BW | 0.251 [0.192, 0.312] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belabbas, R.; Ezzeroug, R.; Berbar, A.; de la Luz Garcia, M.; Zitouni, G.; Taalaziza, D.; Boudjella, Z.; Boudahdir, N.; Diss, S.; Argente, M.-J. Genetic Analyses of Rabbit Survival and Individual Birth Weight. Animals 2022, 12, 2695. https://doi.org/10.3390/ani12192695
Belabbas R, Ezzeroug R, Berbar A, de la Luz Garcia M, Zitouni G, Taalaziza D, Boudjella Z, Boudahdir N, Diss S, Argente M-J. Genetic Analyses of Rabbit Survival and Individual Birth Weight. Animals. 2022; 12(19):2695. https://doi.org/10.3390/ani12192695
Chicago/Turabian StyleBelabbas, Rafik, Rym Ezzeroug, Ali Berbar, María de la Luz Garcia, Ghania Zitouni, Djamel Taalaziza, Zoulikha Boudjella, Nassima Boudahdir, Samir Diss, and María-José Argente. 2022. "Genetic Analyses of Rabbit Survival and Individual Birth Weight" Animals 12, no. 19: 2695. https://doi.org/10.3390/ani12192695