
Effect of Acute Cold Stress on Neuroethology in Mice and Establishment of Its Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Treatment
2.1.1. Animal Management
2.1.2. Cold Exposure
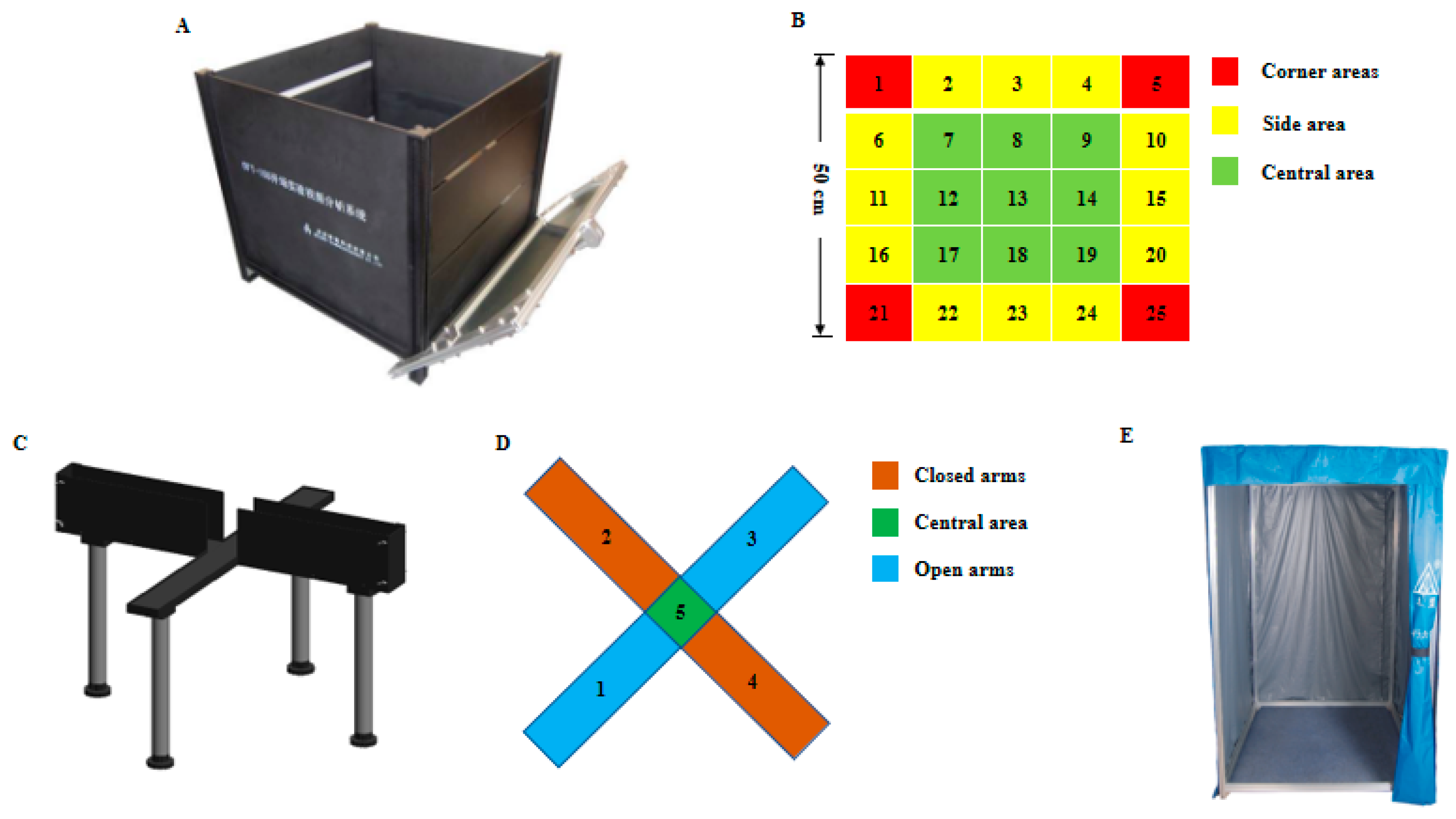
2.2. Behavioral Measurement and Operating Procedures
2.3. Blood Sample Collection and Determination of Serum Related Hormones
2.4. Statistical Analysis
3. Results
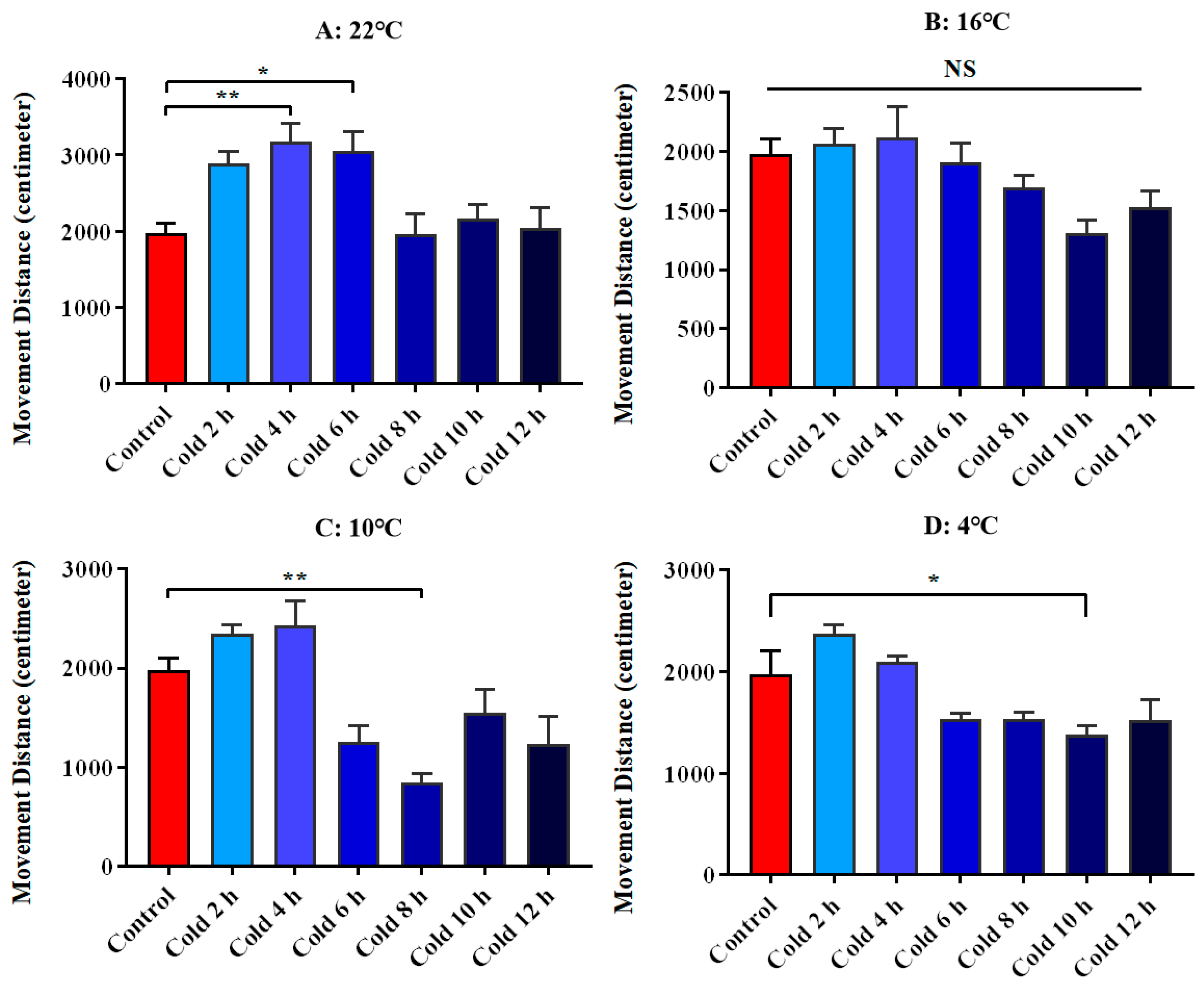
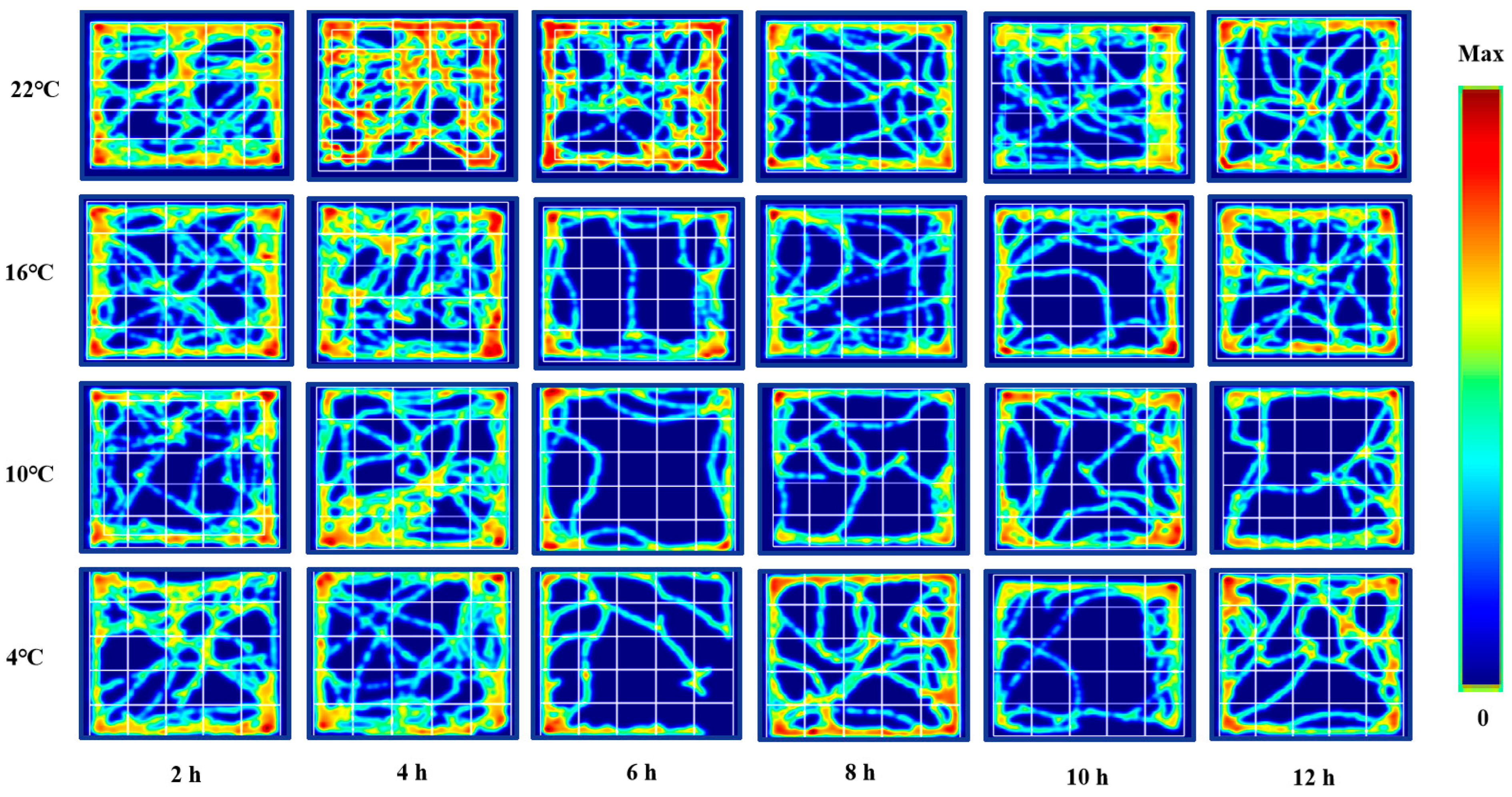
3.1. Changes in Spontaneous Movement of ICR Mice in Open Field Test under Different Intensity of Cold Exposure
3.2. Changes in Exploratory Behavior of ICR Mice in Open Field Test under Different Intensity of Cold Exposure
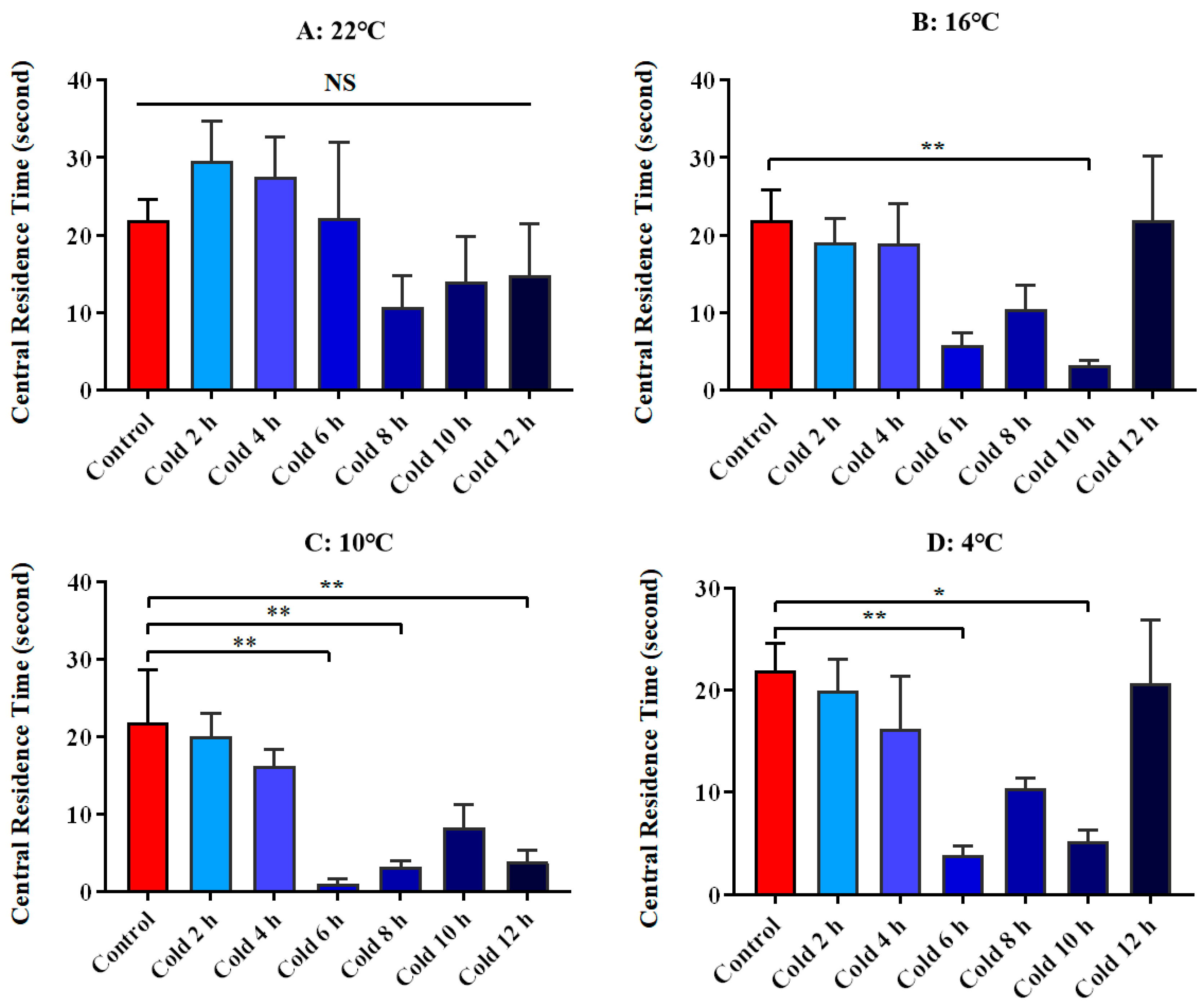
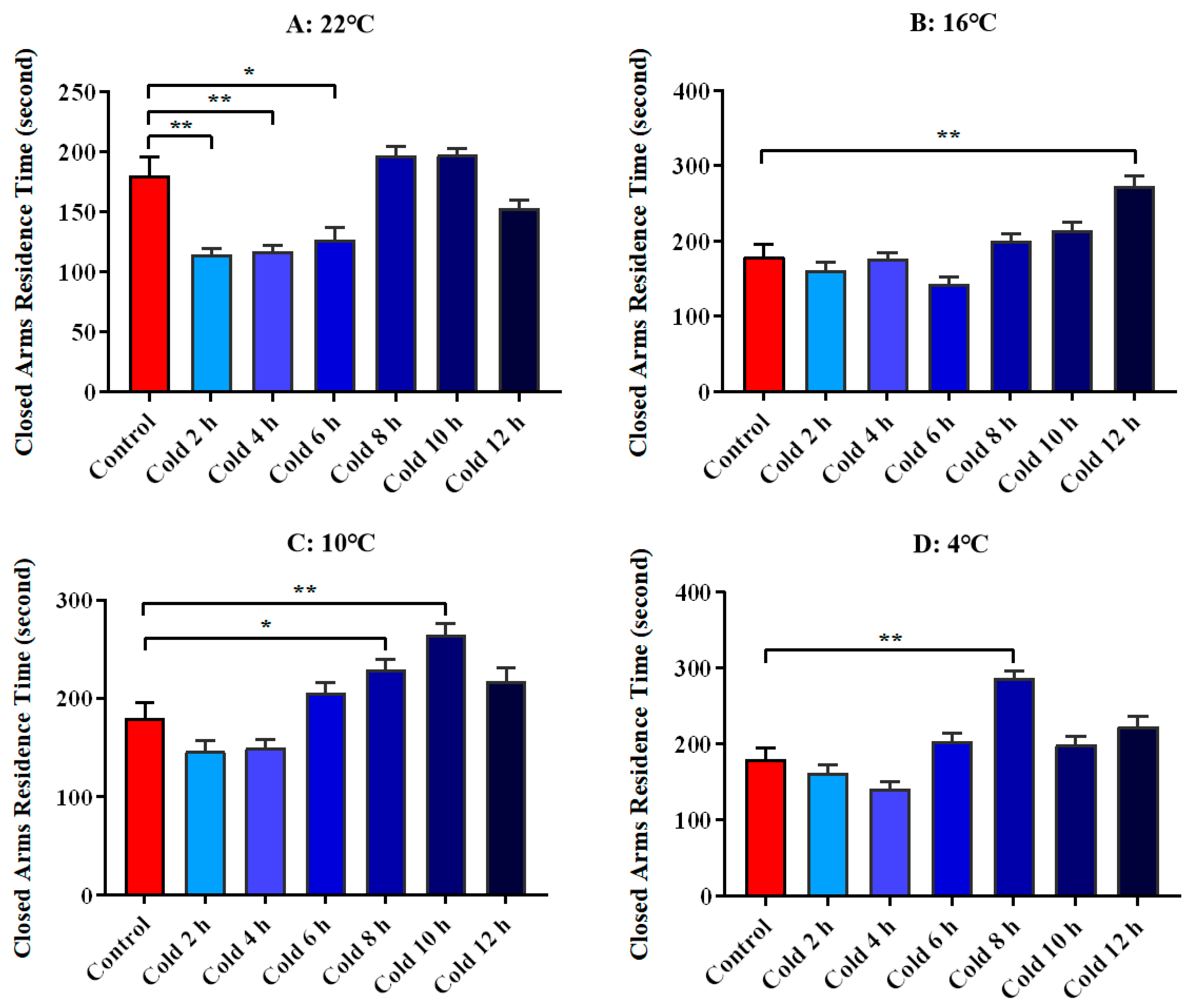
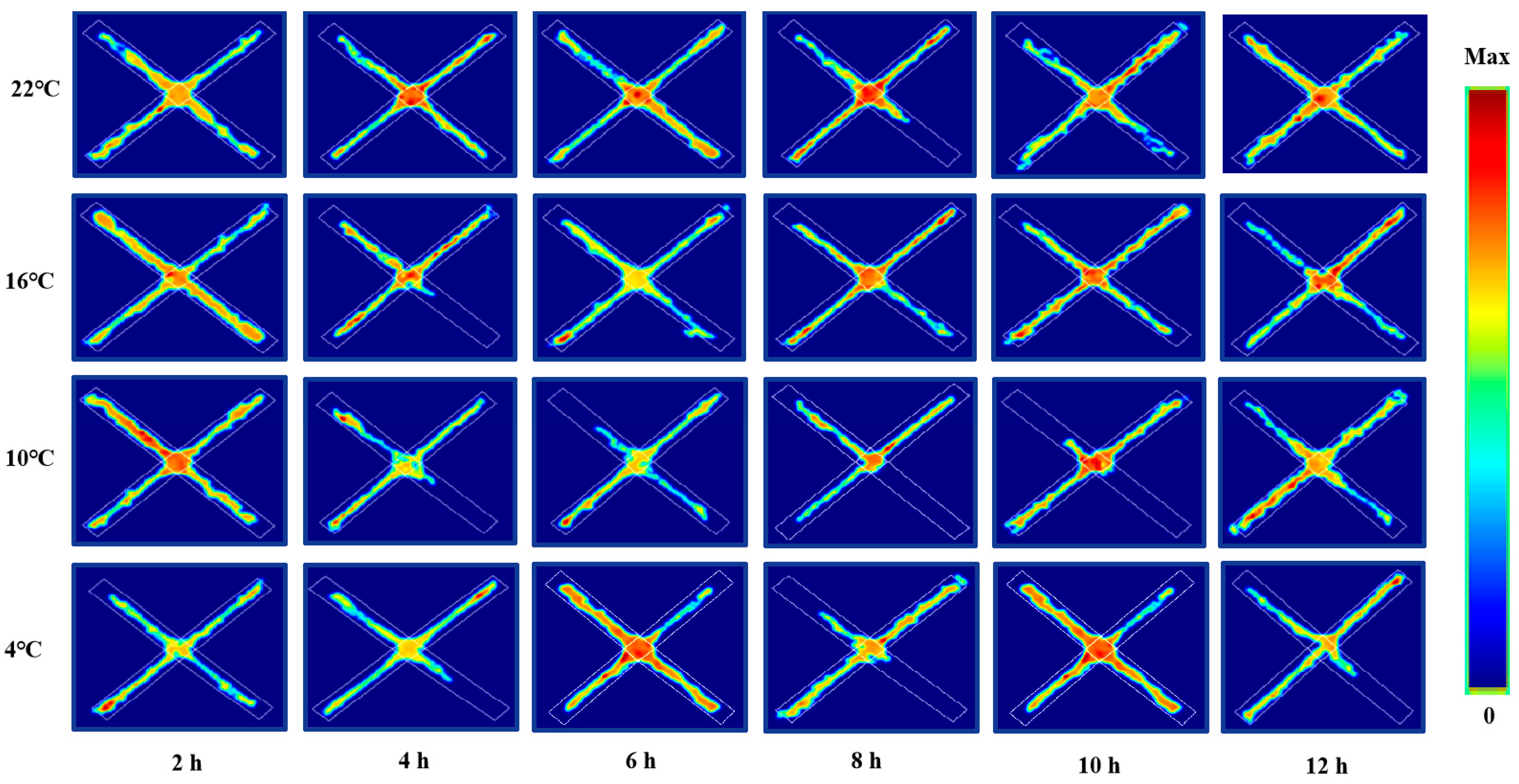
3.3. Changes in Anxious Mood of ICR Mice in Elevated Plus Maze Test under Different Intensity of Cold Exposure
3.4. Changes in Related Hormones of ICR Mice in Elevated Plus Maze Test under Different Intensity of Cold Exposure
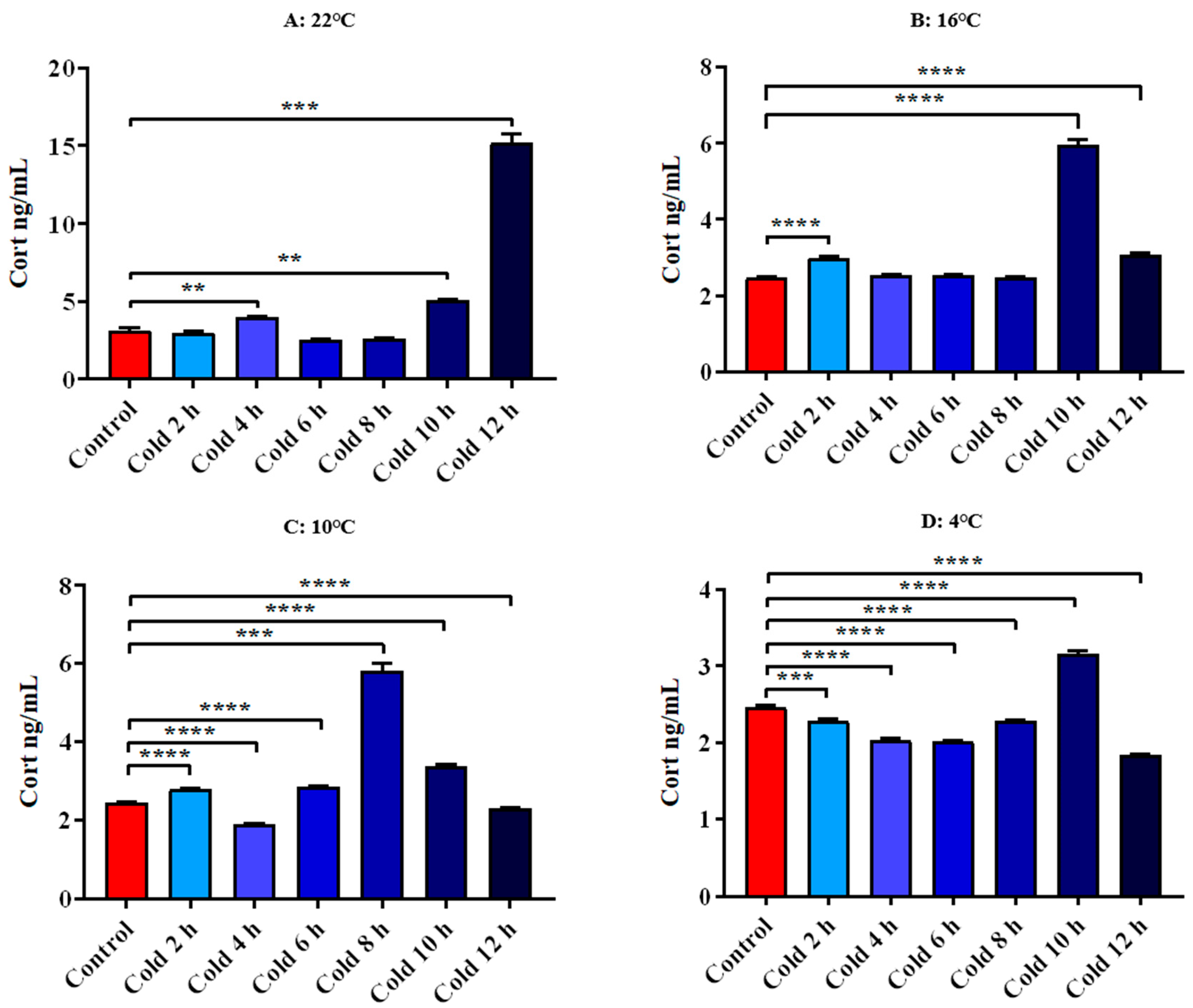
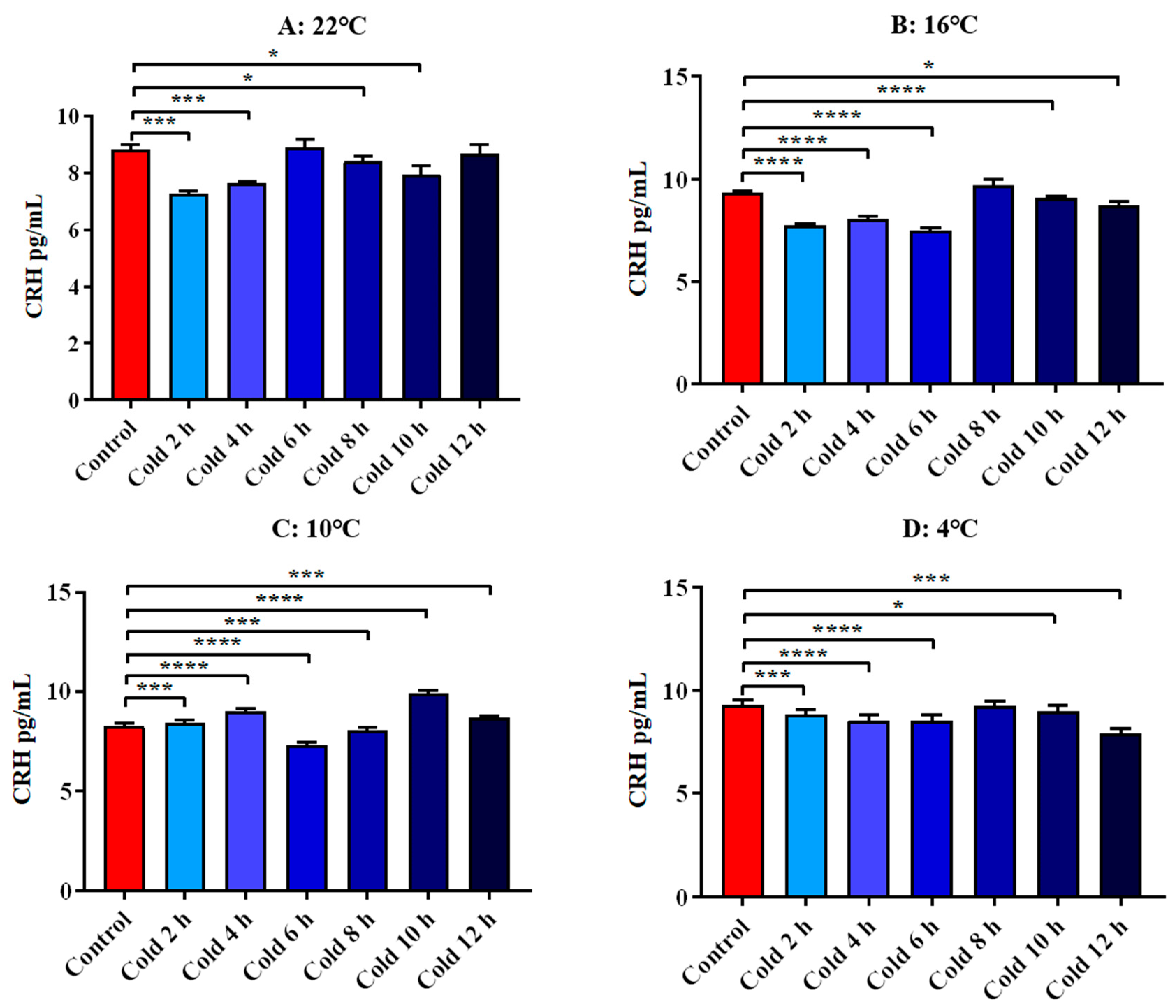
3.4.1. Changes in Serum Cort and CRH of ICR Mice in Elevated Plus Maze Test under Different Intensity of Cold Exposure
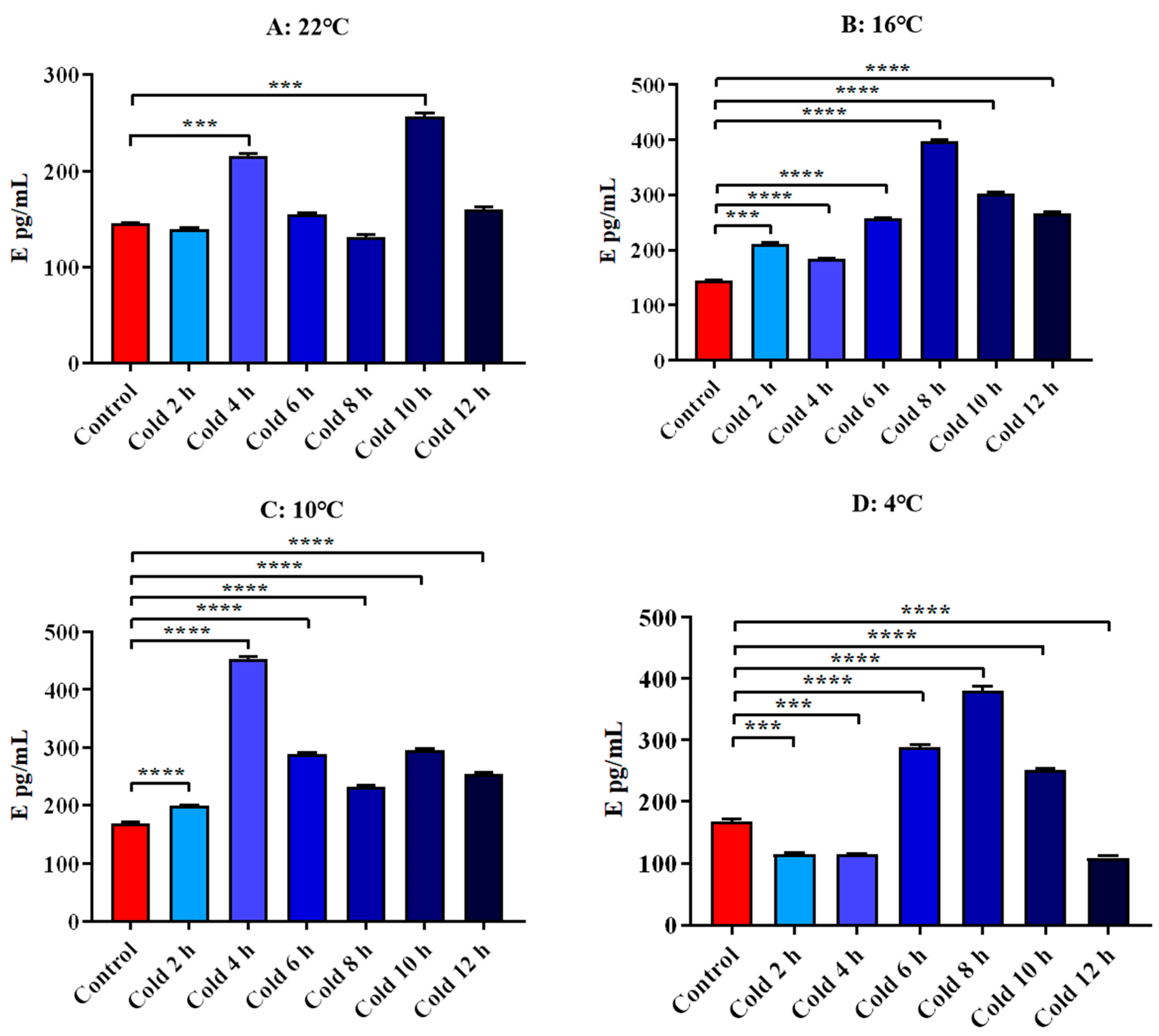
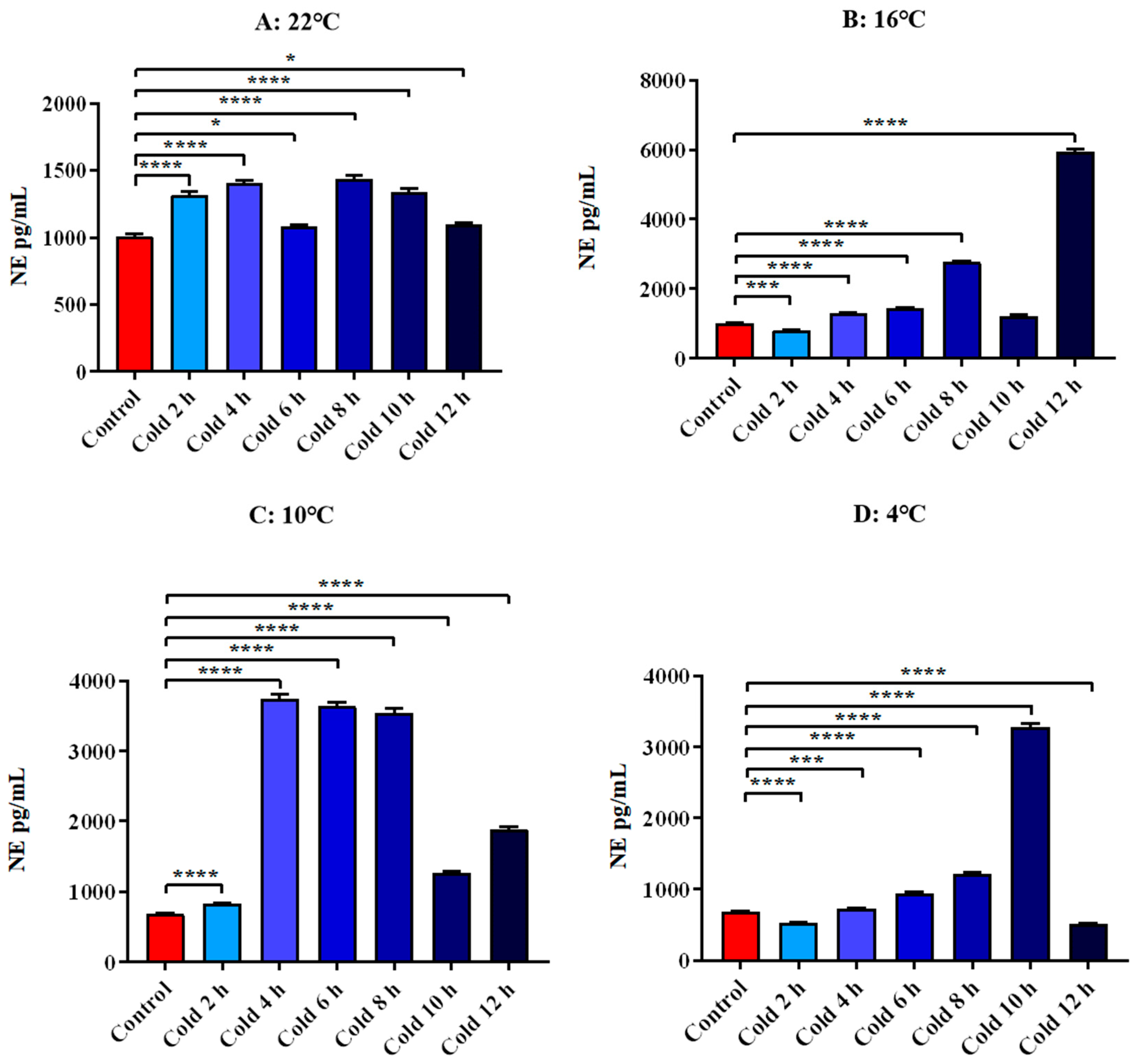
3.4.2. Changes in Serum E and NE of ICR Mice in Elevated Plus Maze Test under Different Intensity of Cold Exposure
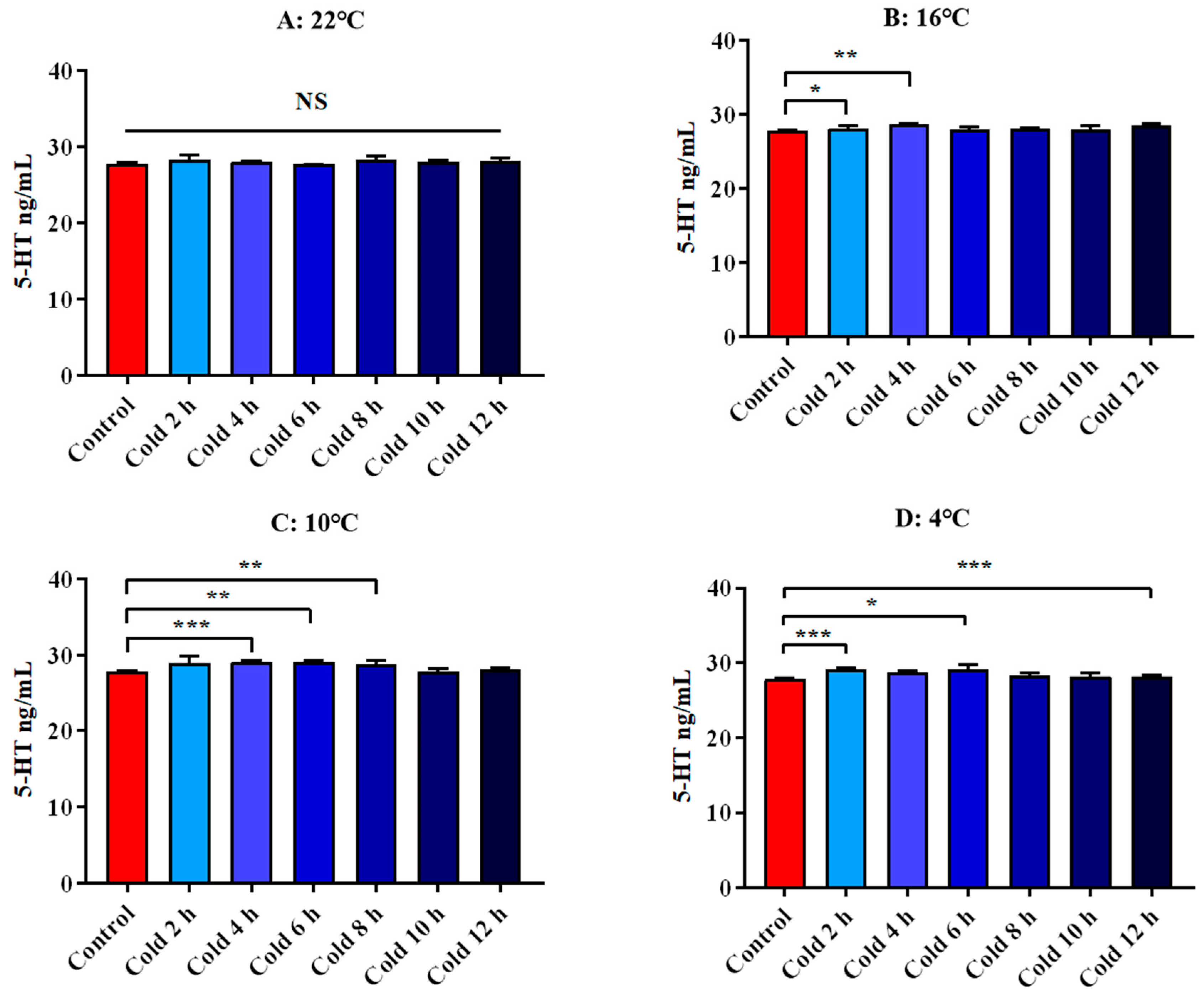
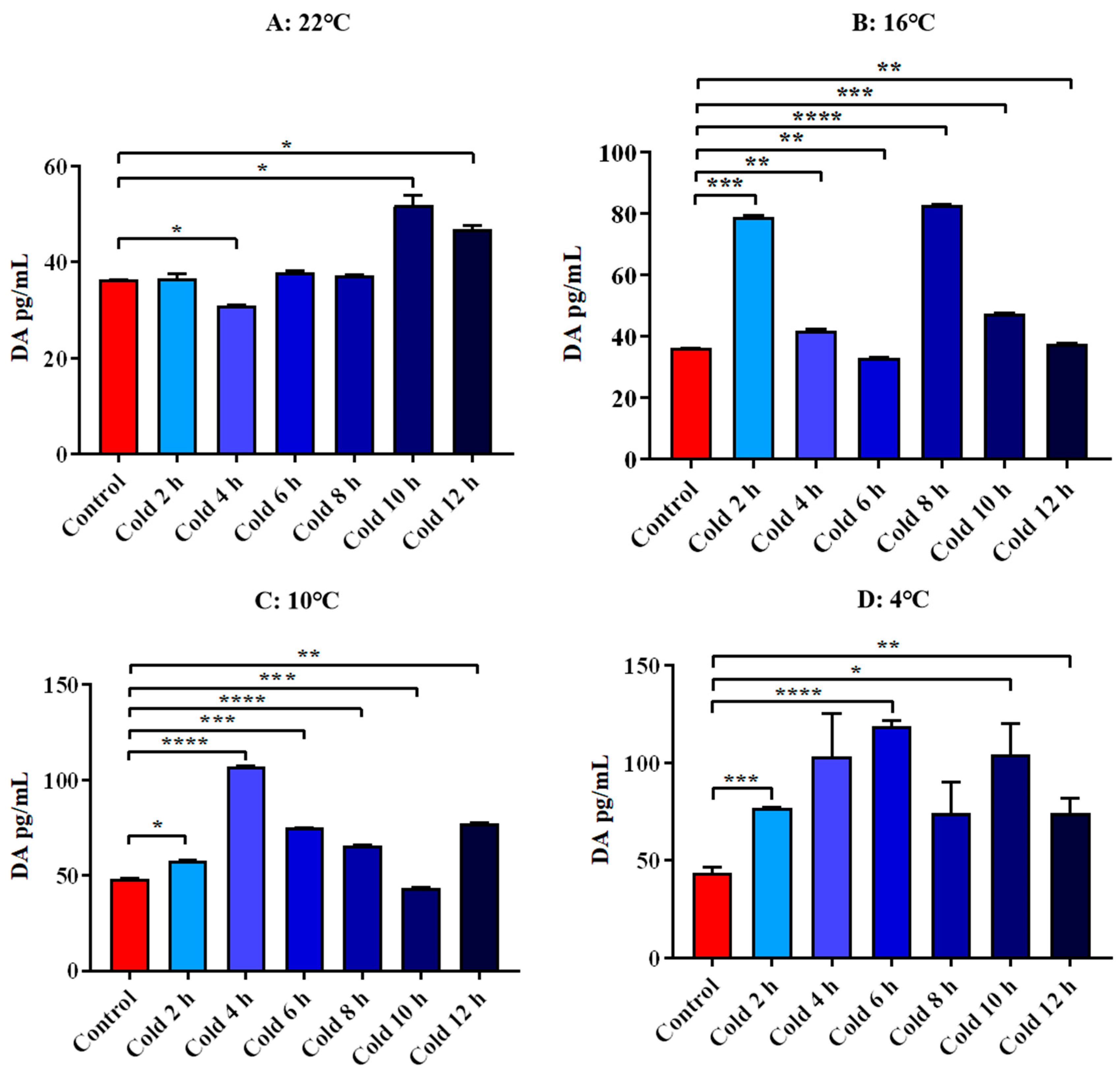
3.4.3. Changes in Serum 5-HT and DA of ICR Mice in Elevated Plus Maze Test under Different Intensity of Cold Exposure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yao, R.; Yang, Y.; Lian, S.; Shi, H.; Liu, P.; Liu, Y.; Yang, H.; Li, S. Effects of Acute Cold Stress on Liver O-GlcNAcylation and Glycometabolism in Mice. Int. J. Mol. Sci. 2018, 19, 2815. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Yao, R.; Lian, S.; Liu, P.; Liu, Y.; Yang, Y.Y.; Yang, H.; Li, S. Regulating glycolysis, the TLR4 signal pathway and expression of RBM3 in mouse liver in response to acute cold exposure. Stress 2019, 22, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Yao, R.; Shi, H.; Liu, Y.; Lian, S.; Yang, Y.; Yang, H.; Li, S. Effects of Cold-inducible RNA-binding Protein (CIRP) on Liver Glycolysis during Acute Cold Exposure in C57BL/6 Mice. Int. J. Mol. Sci. 2019, 20, 1470. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Lang, L.-M.; Li, S.-Z.; Guo, J.-R.; Wang, J.-F.; Wang, D.; Zhang, L.-P.; Yang, H.-M.; Lian, S. Cortisol Excess-Mediated Mitochondrial Damage Induced Hippocampal Neuronal Apoptosis in Mice Following Cold Exposure. Cells 2019, 8, 612. [Google Scholar] [CrossRef] [Green Version]
- Lian, S.; Guo, J.; Wang, L.; Li, W.; Wang, J.; Ji, H.; Kong, F.; Xu, B.; Li, S.; Yang, H. Impact of prenatal cold stress on placental physiology, inflammatory response, and apoptosis in rats. Oncotarget 2017, 8, 115304–115314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Niu, C.; Jia, H.; Chen, X. Effects of acute cold exposure on oxidative balance and total antioxidant capacity in juvenile Chinese soft-shelled turtle, Pelodiscus sinensis. Integr. Zool. 2017, 12, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Sturman, O.; Germain, P.-L.; Bohacek, J. Exploratory rearing: A context- and stress-sensitive behavior recorded in the open-field test. Stress 2018, 21, 443–452. [Google Scholar] [CrossRef]
- Willner, P. Chronic mild stress (CMS) revisited: Consistency and behavioural-neurobiological concordance in the effects of CMS. Neuropsychobiology 2005, 52, 90–110. [Google Scholar] [CrossRef] [PubMed]
- Omata, K. Behavior and body weight changes under intermittent exposure to cold in C57BL/6J mice. Physiol. Behav. 1990, 47, 743–748. [Google Scholar] [CrossRef]
- Castellani, J.W.; Young, A.J. Human physiological responses to cold exposure: Acute responses and acclimatization to prolonged exposure. Auton. Neurosci. 2016, 196, 63–74. [Google Scholar] [CrossRef]
- Xu, X.; Tikuisis, P. Thermoregulatory Modeling for Cold Stress. Compr. Physiol. 2014, 4, 1057–1081. [Google Scholar] [PubMed]
- Glaser, R.; Kiecolt-Glaser, J.K. Stress-induced immune dysfunction: Implications for health. Nat. Rev. Immunol. 2005, 5, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Chrousos, G.P. Stress and disorders of the stress system. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef]
- Eshkevari, L.; Permaul, E.; Mulroney, S.E. Acupuncture blocks cold stress-induced increases in the hypothalamus-pituitary-adrenal axis in the rat. J. Endocrinol. 2013, 217, 95–104. [Google Scholar] [CrossRef]
- Jedema, H.P.; Finlay, J.M.; Sved, A.F.; Grace, A.A. Chronic cold exposure potentiates CRH-evoked increases in electrophysiologic activity of locus coeruleus neurons. Biol. Psychiatry 2001, 49, 351–359. [Google Scholar] [CrossRef]
- Krizanova, O.; Babula, P.; Pacak, K. Stress, catecholaminergic system and cancer. Stress 2016, 19, 419–428. [Google Scholar] [CrossRef]
- Eimonte, M.; Eimantas, N.; Daniuseviciute, L.; Paulauskas, H.; Vitkauskiene, A.; Dauksaite, G.; Brazaitis, M. Recovering body temperature from acute cold stress is associated with delayed proinflammatory cytokine production in vivo. Cytokine 2021, 143, 155510. [Google Scholar] [CrossRef]
- Raimondi, L.A.; Alfarano, C.; Pacini, A.; Livi, S.; Ghelardini, C.; DeSiena, G.; Pirisino, R. Methylamine-dependent release of nitric oxide and dopamine in the CNS modulates food intake in fasting rats. Br. J. Pharmacol. 2007, 150, 1003–1010. [Google Scholar] [CrossRef] [Green Version]
- Prasad, G.L.; Naik, B.R.; Ko, J.E.; Nagaraju, G.P. Effects of naloxone, serotonin, and dopamine on reproduction of the freshwater crab Barytelphusa guerini. J. Exp. Zool. A Ecol. Genet Physiol. 2014, 321, 173–182. [Google Scholar] [CrossRef]
- Henrikson, Z.; Vermette, C.; Schwean-Lardner, K.; Crowe, T. Effects of cold exposure on physiology, meat quality, and behavior of turkey hens and toms crated at transport density. Poult. Sci. 2018, 97, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Abdulhay, A.; Benton, N.A.; Klingerman, C.M.; Krishnamoorthy, K.; Brozek, J.M.; Schneider, J.E. Estrous cycle fluctuations in sex and ingestive behavior are accentuated by exercise or cold ambient temperatures. Horm. Behav. 2014, 66, 135–147. [Google Scholar] [CrossRef] [Green Version]
- Henderson, D.; Fort, M.M.; Rashotte, M.E.; Henderson, R.P. Ingestive behavior and body temperature of pigeons during long-term cold exposure. Physiol. Behav. 1992, 52, 455–469. [Google Scholar] [CrossRef]
- Li, W.J.; Yang, H.M.; Lian, S.; Xu, B.; Wang, L.P.; Zang, S.C.; Bin Yuan, J.; Ding, M.Y. Effects of prenatal maternal cold stress on spontaneous, exploratory and anxious behaviors in offspring. Zhongguo Ying Yong Sheng Li Xue Za Zhi 2019, 35, 135–139. [Google Scholar] [PubMed]
- Lian, S.; Wang, D.; Xu, B.; Guo, W.; Wang, L.; Li, W.; Ji, H.; Wang, J.; Kong, F.; Zhen, L.; et al. Prenatal cold stress: Effect on maternal hippocampus and offspring behavior in rats. Behav. Brain Res. 2018, 346, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lian, S.; Xu, B.; Wang, D.; Wang, L.; Li, W.; Yao, R.; Ji, H.; Wang, J.; Guo, J.; Li, S.; et al. Possible mechanisms of prenatal cold stress induced-anxiety-like behavior depression in offspring rats. Behav. Brain Res. 2019, 359, 304–311. [Google Scholar] [CrossRef]
- Garcia, M.J.; Teets, N.M. Cold stress results in sustained locomotor and behavioral deficits in Drosophila melanogaster. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2019, 331, 192–200. [Google Scholar] [CrossRef]
- Castellani, J.W.; Tipton, M.J. Cold Stress Effects on Exposure Tolerance and Exercise Performance. Compr. Physiol. 2015, 6, 443–469. [Google Scholar] [PubMed]
- Kuniishi, H.; Ichisaka, S.; Yamamoto, M.; Ikubo, N.; Matsuda, S.; Futora, E.; Harada, R.; Ishihara, K.; Hata, Y. Early deprivation increases high-leaning behavior, a novel anxiety-like behavior, in the open field test in rats. Neurosci. Res. 2017, 123, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Choleris, E.; Thomas, A.W.; Kavaliers, M.; Prato, F.S. A detailed ethological analysis of the mouse open field test: Effects of diazepam, chlordiazepoxide and an extremely low frequency pulsed magnetic field. Neurosci. Biobehav. Rev. 2001, 25, 235–260. [Google Scholar] [CrossRef]
- Kraeuter, A.-K.; Guest, P.C.; Sarnyai, Z. The Open Field Test for Measuring Locomotor Activity and Anxiety-Like Behavior. Methods Mol. Biol. 2019, 1916, 99–103. [Google Scholar]
- DeVallance, E.; Riggs, D.; Jackson, B.; Parkulo, T.; Zaslau, S.; Chantler, P.D.; Olfert, I.M.; Bryner, R.W. Effect of chronic stress on running wheel activity in mice. PLoS ONE 2017, 12, e0184829. [Google Scholar] [CrossRef] [PubMed]
- Von Ziegler, L.M.; Floriou-Servou, A.; Waag, R.; Das Gupta, R.R.; Sturman, O.; Gapp, K.; Maat, C.A.; Kockmann, T.; Lin, H.Y.; Duss, S.N.; et al. Multiomic profiling of the acute stress response in the mouse hippocampus. Nat. Commun. 2022, 13, 1824. [Google Scholar] [CrossRef] [PubMed]
- Oksa, J.; Rintamäki, H.; Mäkinen, T. The effect of training of military skills on performance in cold environment. Mil. Med. 2006, 171, 757–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparling, J.E.; Barbeau, K.; Boileau, K.; Konkle, A.T. Environmental enrichment and its influence on rodent offspring and maternal behaviours, a scoping style review of indices of depression and anxiety. Pharmacol. Biochem. Behav. 2020, 197, 172997. [Google Scholar] [CrossRef] [PubMed]
- Badache, S.; Bouslama, S.; Brahmia, O.; Baïri, A.M.; Tahraoui, A.K.; Ladjama, A. Prenatal noise and restraint stress interact to alter exploratory behavior and balance in juvenile rats, and mixed stress reverses these effects. Stress 2017, 20, 320–328. [Google Scholar] [CrossRef]
- Ostheim, J. Coping with food-limited conditions: Feeding behavior, temperature preference, and nocturnal hypothermia in pigeons. Physiol. Behav. 1992, 51, 353–361. [Google Scholar] [CrossRef]
- Donald, R.D.; Healy, S.D.; Lawrence, A.B.; Rutherford, K.M. Emotionality in growing pigs: Is the open field a valid test? Physiol. Behav. 2011, 104, 906–913. [Google Scholar] [CrossRef]
- Knight, P.; Chellian, R.; Wilson, R.; Behnood-Rod, A.; Panunzio, S.; Bruijnzeel, A.W. Sex differences in the elevated plus-maze test and large open field test in adult Wistar rats. Pharmacol. Biochem. Behav. 2021, 204, 173168. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Elevated Plus Maze Test for Measuring Anxiety-Like Behavior in Rodents. Methods Mol. Biol. 2019, 1916, 69–74. [Google Scholar]
- Costa, A.A.; Morato, S.; Roque, A.C.; Tinós, R. A computational model for exploratory activity of rats with different anxiety levels in elevated plus-maze. J. Neurosci. Methods 2014, 236, 44–50. [Google Scholar] [CrossRef]
- DeGroot, S.R.; Zhao-Shea, R.; Chung, L.; Klenowski, P.M.; Sun, F.; Molas, S.; Gardner, P.D.; Li, Y.; Tapper, A.R. Midbrain Dopamine Controls Anxiety-like Behavior by Engaging Unique Interpeduncular Nucleus Microcircuitry. Biol. Psychiatry 2020, 88, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.C.; Fogaca, M.V.; Aguiar, D.C.; Guimaraes, F.S. Animal models of anxiety disorders and stress. Braz. J. Psychiatry 2013, 35 (Suppl. 2), S101–S111. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Matsunaga, D.; Ishiwata, T. Effect of heat acclimation on anxiety-like behavior of rats in an open field. J. Therm. Biol. 2020, 87, 102458. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.-M.; Yuan, S.-N.; Guan, X.-T.; Xie, X.; Shao, F.; Wang, W.-W. Juvenile stress affects anxiety-like behavior and limbic monoamines in adult rats. Physiol. Behav. 2014, 135, 7–16. [Google Scholar] [CrossRef]
- Vallee, M.; Mayo, W.; Dellu, F.; Le Moal, M.; Simon, H.; Maccari, S. Prenatal stress induces high anxiety and postnatal handling induces low anxiety in adult offspring: Correlation with stress-induced corticosterone secretion. J. Neurosci. 1997, 17, 2626–2636. [Google Scholar] [CrossRef] [Green Version]
- Freels, T.G.; Lester, D.B.; Cook, M.N. Arachidonoyl serotonin (AA-5-HT) modulates general fear-like behavior and inhibits mesolimbic dopamine release. Behav. Brain Res. 2019, 362, 140–151. [Google Scholar] [CrossRef]
- Tseng, H.-H.; Watts, J.; Kiang, M.; Suridjan, I.; Wilson, A.A.; Houle, S.; Rusjan, P.; Mizrahi, R. Nigral Stress-Induced Dopamine Release in Clinical High Risk and Antipsychotic-Naïve Schizophrenia. Schizophr. Bull 2018, 44, 542–551. [Google Scholar] [CrossRef] [Green Version]
- Pijlman, F.T.; van Ree, J.M. Physical but not emotional stress induces a delay in behavioural coping responses in rats. Behav. Brain Res. 2002, 136, 365–373. [Google Scholar] [CrossRef]
- Pijlman, F.T.; Herremans, A.H.; van de Kieft, J.; Kruse, C.G.; van Ree, J.M. Behavioural changes after different stress paradigms: Prepulse inhibition increased after physical, but not emotional stress. Eur. Neuropsychopharmacol. 2003, 13, 369–380. [Google Scholar] [CrossRef]
- Bacqué-Cazenave, J.; Bharatiya, R.; Barrière, G.; Delbecque, J.-P.; Bouguiyoud, N.; Di Giovanni, G.; Cattaert, D.; De Deurwaerdère, P. Serotonin in Animal Cognition and Behavior. Int. J. Mol. Sci. 2020, 21, 1649. [Google Scholar] [CrossRef] [Green Version]
- Popova, N.K.; Naumenko, V.S. 5-HT1A receptor as a key player in the brain 5-HT system. Rev. Neurosci. 2013, 24, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, H.S. Studies on the neuroendocrine role of serotonin. Dan. Med. Bull. 2007, 54, 266–288. [Google Scholar] [PubMed]
- Gutknecht, L.; Popp, S.; Waider, J.; Sommerlandt, F.M.J.; Göppner, C.; Post, A.; Reif, A.; Van Den Hove, D.; Strekalova, T.; Schmitt, A.; et al. Interaction of brain 5-HT synthesis deficiency, chronic stress and sex differentially impact emotional behavior in Tph2 knockout mice. Psychopharmacology 2015, 232, 2429–2441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinstock, M. Prenatal stressors in rodents: Effects on behavior. Neurobiol. Stress 2016, 6, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinlein, S.A.; Phillips, D.J.; Keller, C.R.; Karatsoreos, I.N. Role of corticosterone in altered neurobehavioral responses to acute stress in a model of compromised hypothalamic-pituitary-adrenal axis function. Psychoneuroendocrinology 2019, 102, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.; Wu, H.; Wang, Q.; Cai, M.; Shi, W.; Shang, J. Chronic stress suppresses the expression of cutaneous hypothalamic–pituitary–Adrenocortical axis elements and melanogenesis. PLoS ONE 2014, 9, e98283. [Google Scholar] [CrossRef] [PubMed]
- Spencer, R.L.; Deak, T. A users guide to HPA axis research. Physiol. Behav. 2016, 178, 43–65. [Google Scholar] [CrossRef]
- Peng, B.; Xu, Q.; Liu, J.; Guo, S.; Borgland, S.L.; Liu, S. Corticosterone Attenuates Reward-Seeking Behavior and Increases Anxiety via D2 Receptor Signaling in Ventral Tegmental Area Dopamine Neurons. J. Neurosci. 2021, 41, 1566–1581. [Google Scholar] [CrossRef]
- Lebedeva, K.A.; Caruncho, H.J.; Kalynchuk, L.E. Cyclical corticosterone administration sensitizes depression-like behavior in rats. Neurosci. Lett. 2017, 650, 45–51. [Google Scholar] [CrossRef]
- Cui, X.-Y.; Yang, G.; Cui, S.-Y.; Cao, Q.; Huang, Y.-L.; Ding, H.; Ye, H.; Zhang, X.-Q.; Wang, Z.-J.; Zhang, Y.-H. Sleep patterns deteriorate over time in chronic corticosterone-treated rats. Neurosci. Lett. 2018, 682, 74–78. [Google Scholar] [CrossRef]
- Dieterich, A.; Stech, K.; Srivastava, P.; Lee, J.; Sharif, A.; Samuels, B.A. Chronic corticosterone shifts effort-related choice behavior in male mice. Psychopharmacology 2020, 237, 2103–2110. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.E.; Rincón-Cortés, M.; Braren, S.H.; Brandes-Aitken, A.N.; Opendak, M.; Pollonini, G.; Chopra, D.; Raver, C.C.; Alberini, C.M.; Blair, C.; et al. Corticosterone administration targeting a hypo-reactive HPA axis rescues a socially-avoidant phenotype in scarcity-adversity reared rats. Dev. Cogn. Neurosci. 2019, 40, 100716. [Google Scholar] [CrossRef] [PubMed]
- Singleton, J.M.; Garland, T., Jr. Influence of corticosterone on growth, home-cage activity, wheel running, and aerobic capacity in house mice selectively bred for high voluntary wheel-running behavior. Physiol. Behav. 2019, 198, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Ketchesin, K.D.; Stinnett, G.S.; Seasholtz, A.F. Corticotropin-releasing hormone-binding protein and stress: From invertebrates to humans. Stress 2017, 20, 449–464. [Google Scholar] [CrossRef] [PubMed]
- Jaferi, A.; Bhatnagar, S. Corticotropin-releasing hormone receptors in the medial prefrontal cortex regulate hypothalamic-pituitary-adrenal activity and anxiety-related behavior regardless of prior stress experience. Brain Res. 2007, 1186, 212–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holsboer, F. Corticotropin-releasing hormone modulators and depression. Curr. Opin. Investig. Drugs 2003, 4, 46–50. [Google Scholar] [PubMed]
- Carrasco, G.A.; van de Kar, L.D. Neuroendocrine pharmacology of stress. Eur. J. Pharmacol. 2003, 463, 235–272. [Google Scholar] [CrossRef]
- Prokopová, I. Noradrenaline and behavior. Cesk. Fysiol. 2010, 59, 51–58. [Google Scholar]
- Rougemont-Buecking, A.; Rougemont, E.; Toth, R.; Simon, O.; Besson, J. “Epinephrine-addiction” and risk-seeking behaviour in athletes: What realities? Rev. Med. Suisse 2007, 3, 1536–1540. [Google Scholar]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Liu, Y.; Li, S. Effect of Acute Cold Stress on Neuroethology in Mice and Establishment of Its Model. Animals 2022, 12, 2671. https://doi.org/10.3390/ani12192671
Hu Y, Liu Y, Li S. Effect of Acute Cold Stress on Neuroethology in Mice and Establishment of Its Model. Animals. 2022; 12(19):2671. https://doi.org/10.3390/ani12192671
Chicago/Turabian StyleHu, Yajie, Yang Liu, and Shize Li. 2022. "Effect of Acute Cold Stress on Neuroethology in Mice and Establishment of Its Model" Animals 12, no. 19: 2671. https://doi.org/10.3390/ani12192671